
Hydrogen Sulphide Production in Healthy and Ulcerated Gastric Mucosa of Rats

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
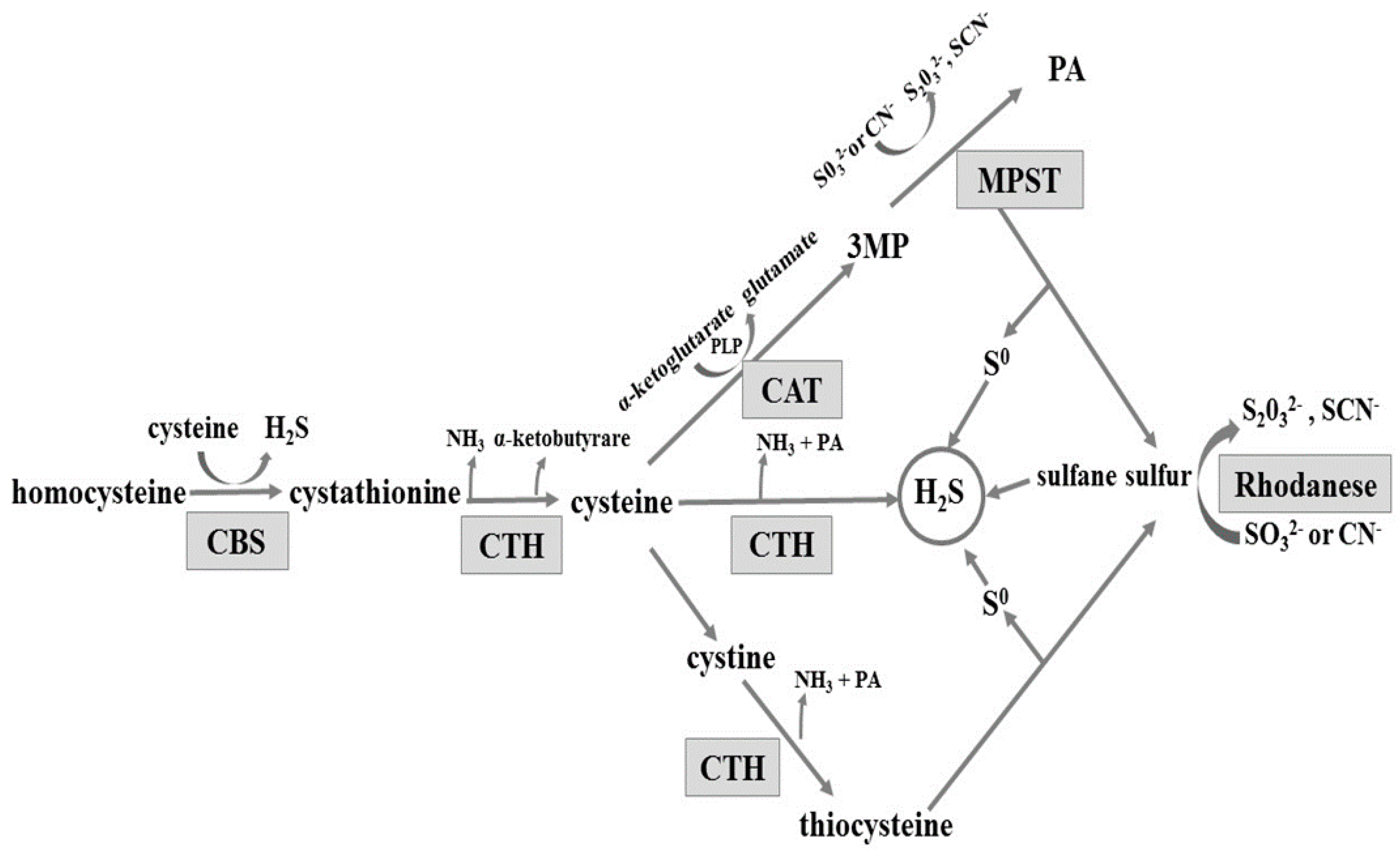
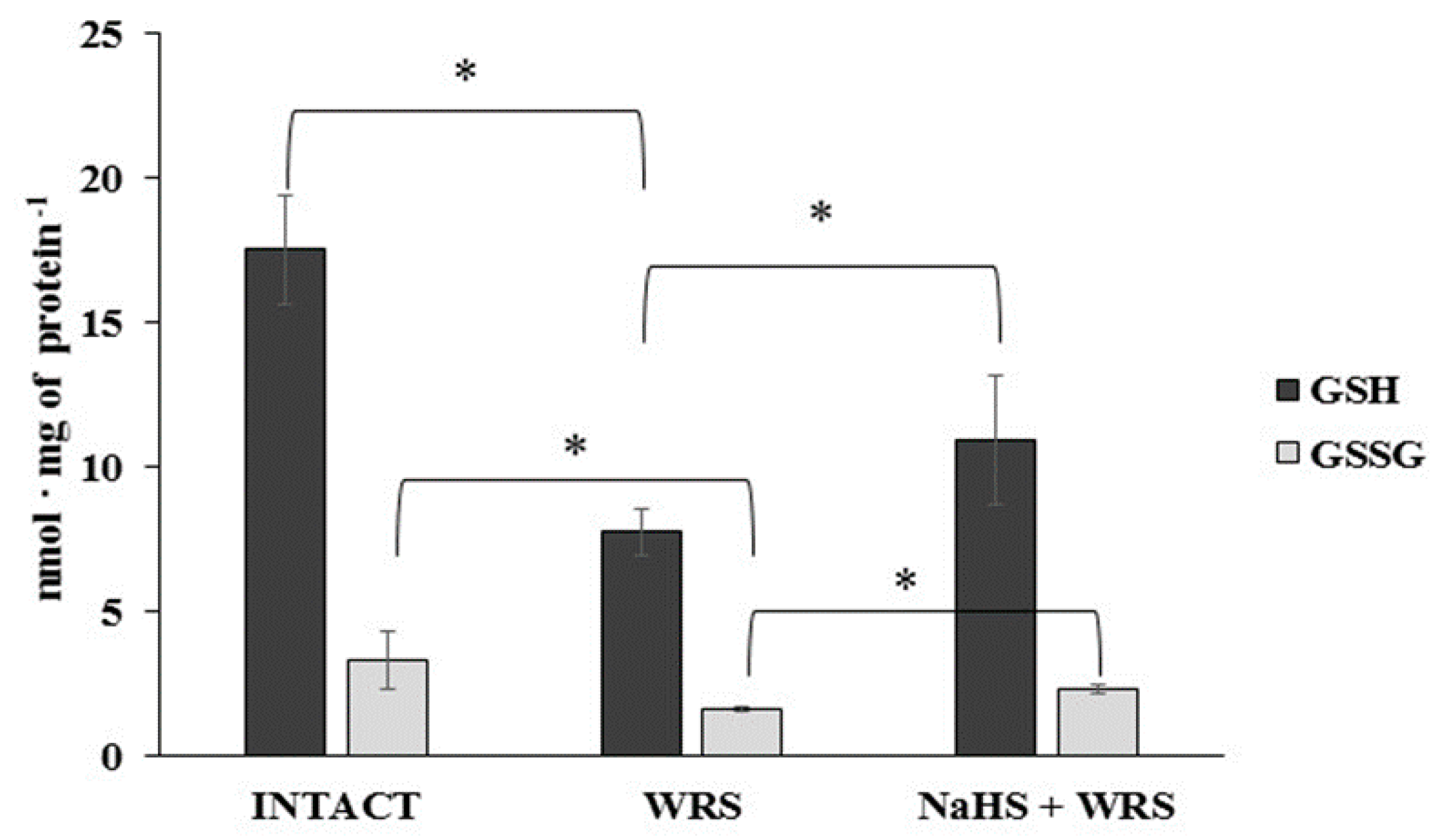
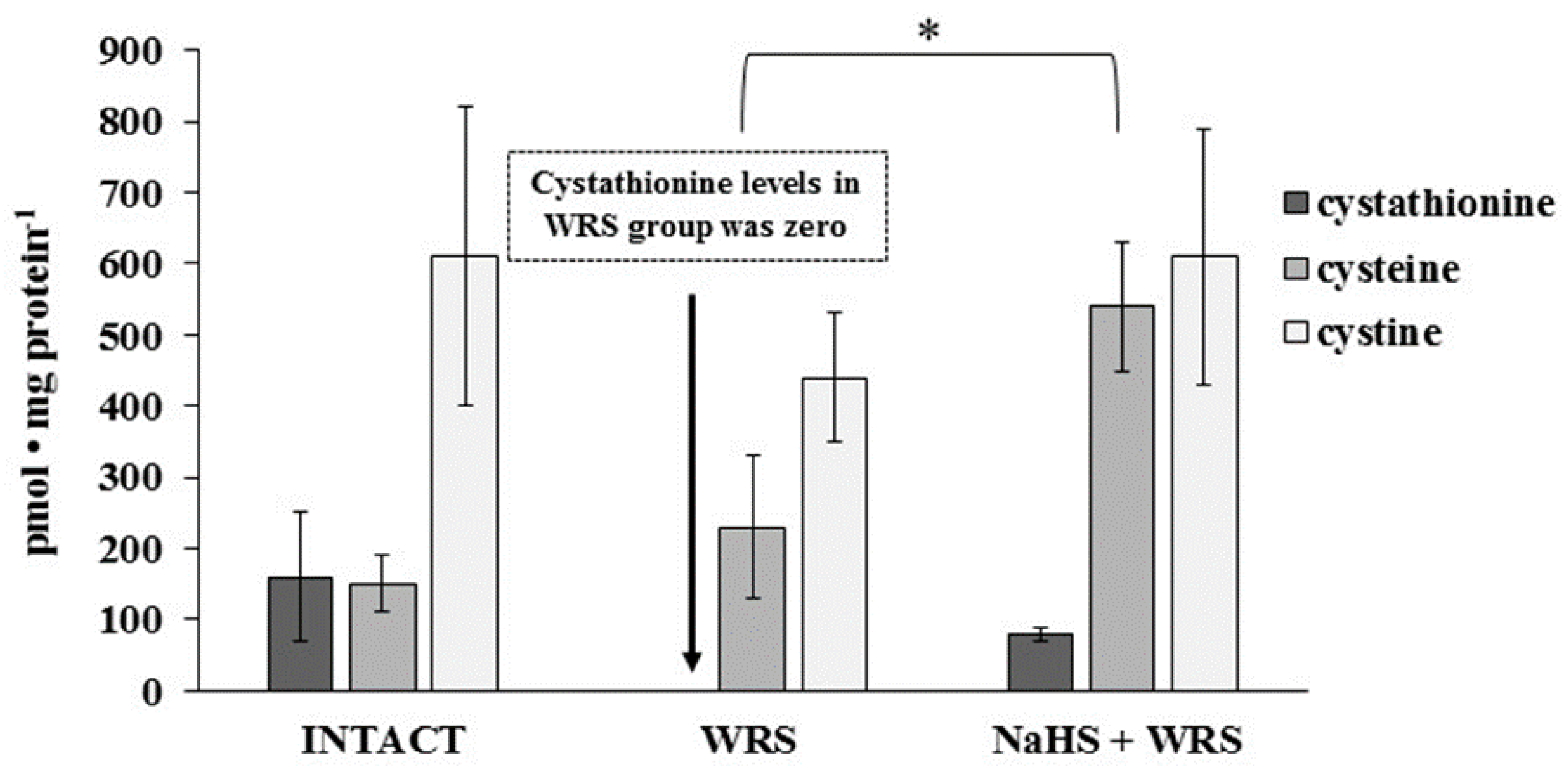
2.1. H2S Generation in Gastric Mucosa of Healthy Rats
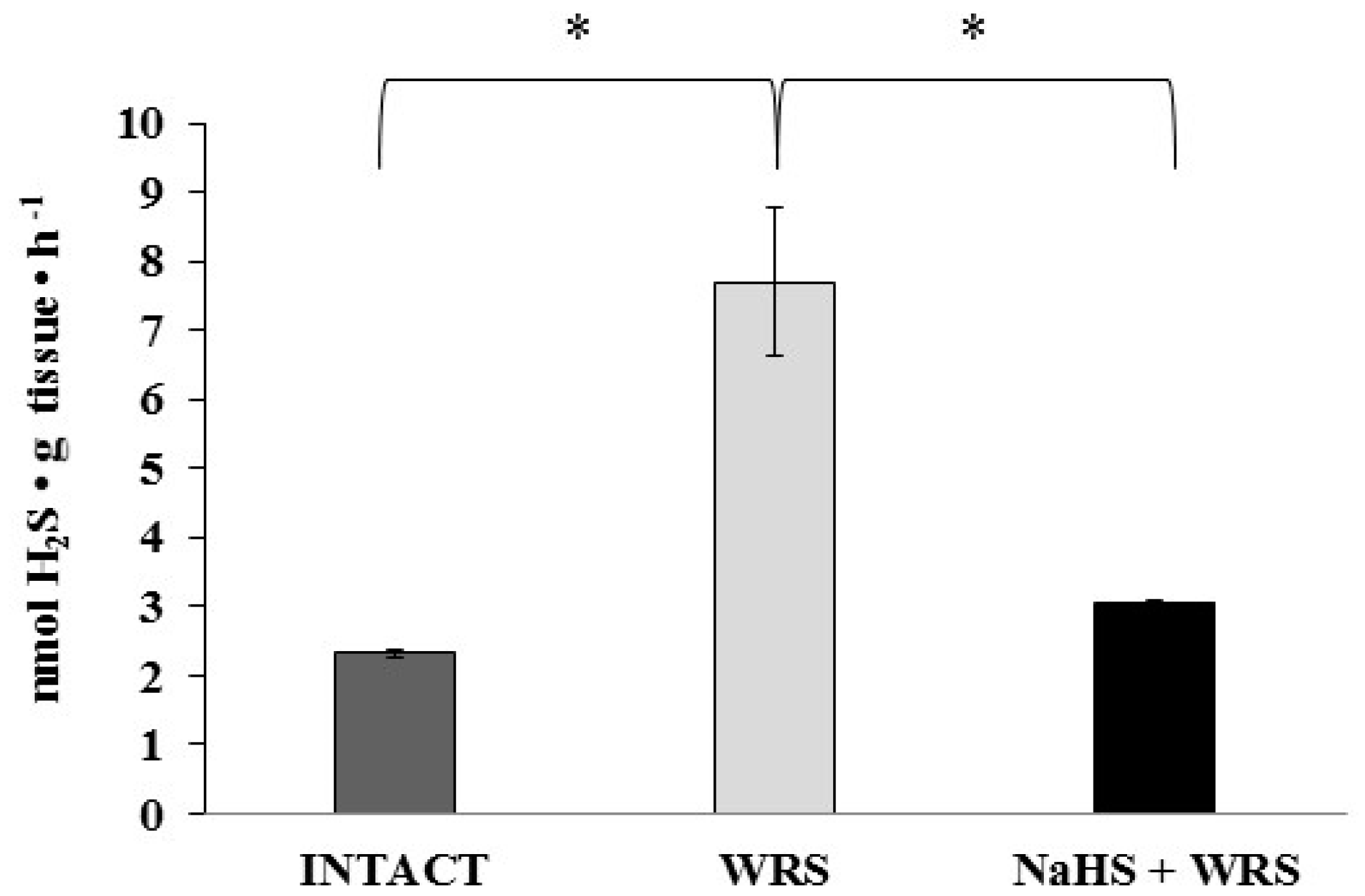
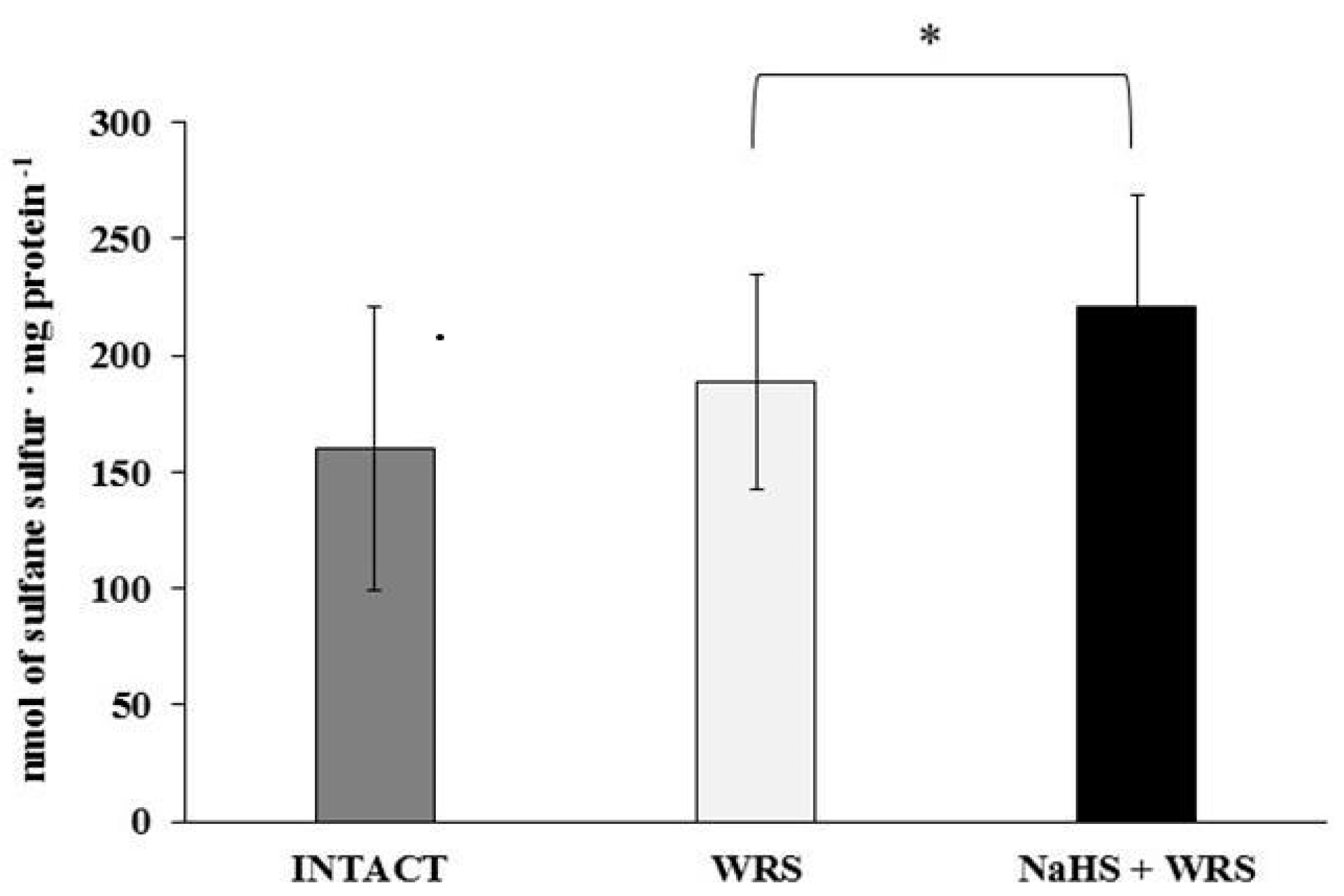
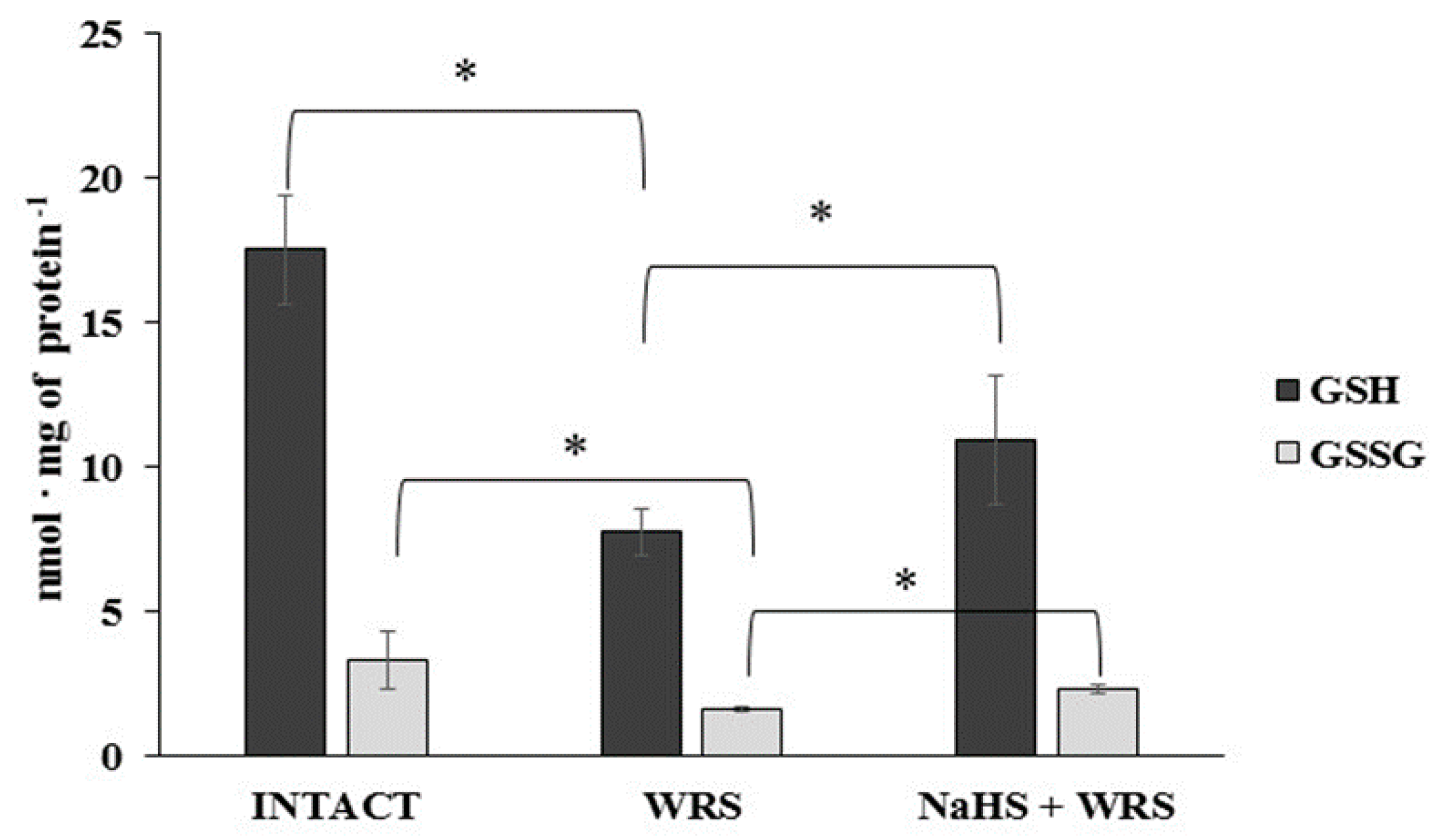
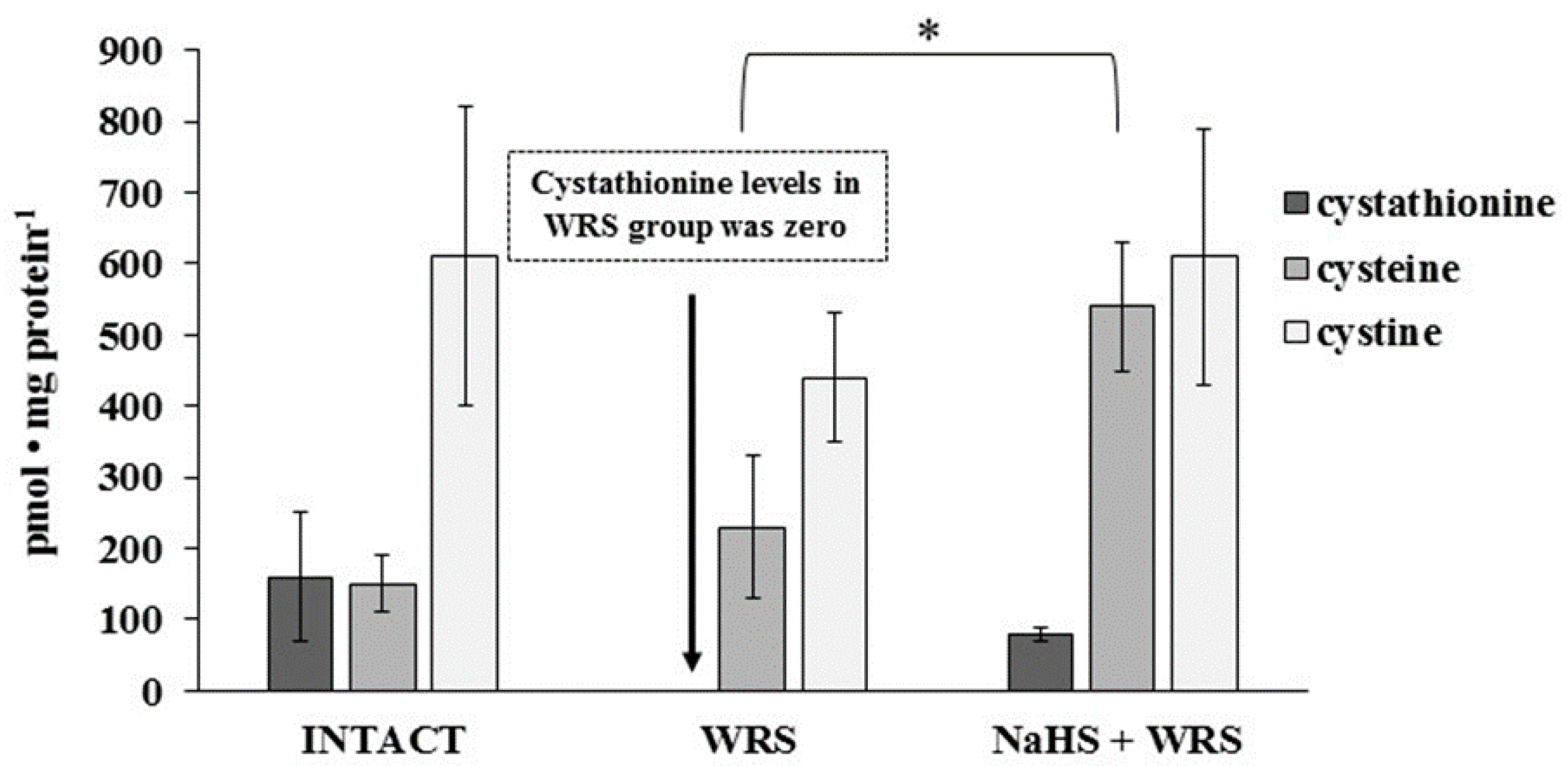
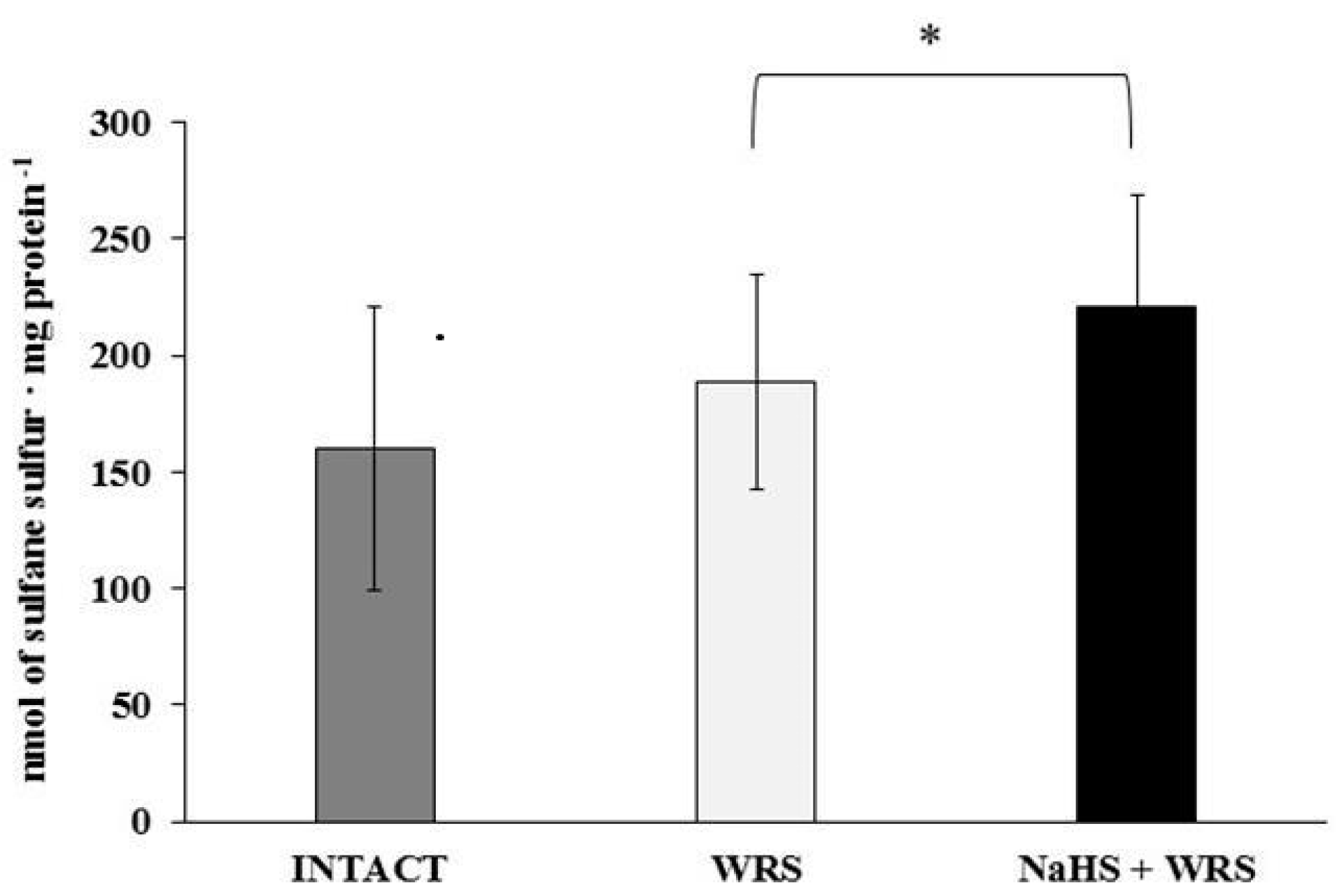
2.2. Gastric Mucosa from Rats with Ulcers Induced by Water Immersion and Restraint Stress
2.3. NaHS Pretreatment Affects the Formation of Hydrogen Sulphide in Gastric Mucosa of Rats with WRS Induced Ulcers
3. Material and Methods
3.1. Chemicals
3.2. Animals
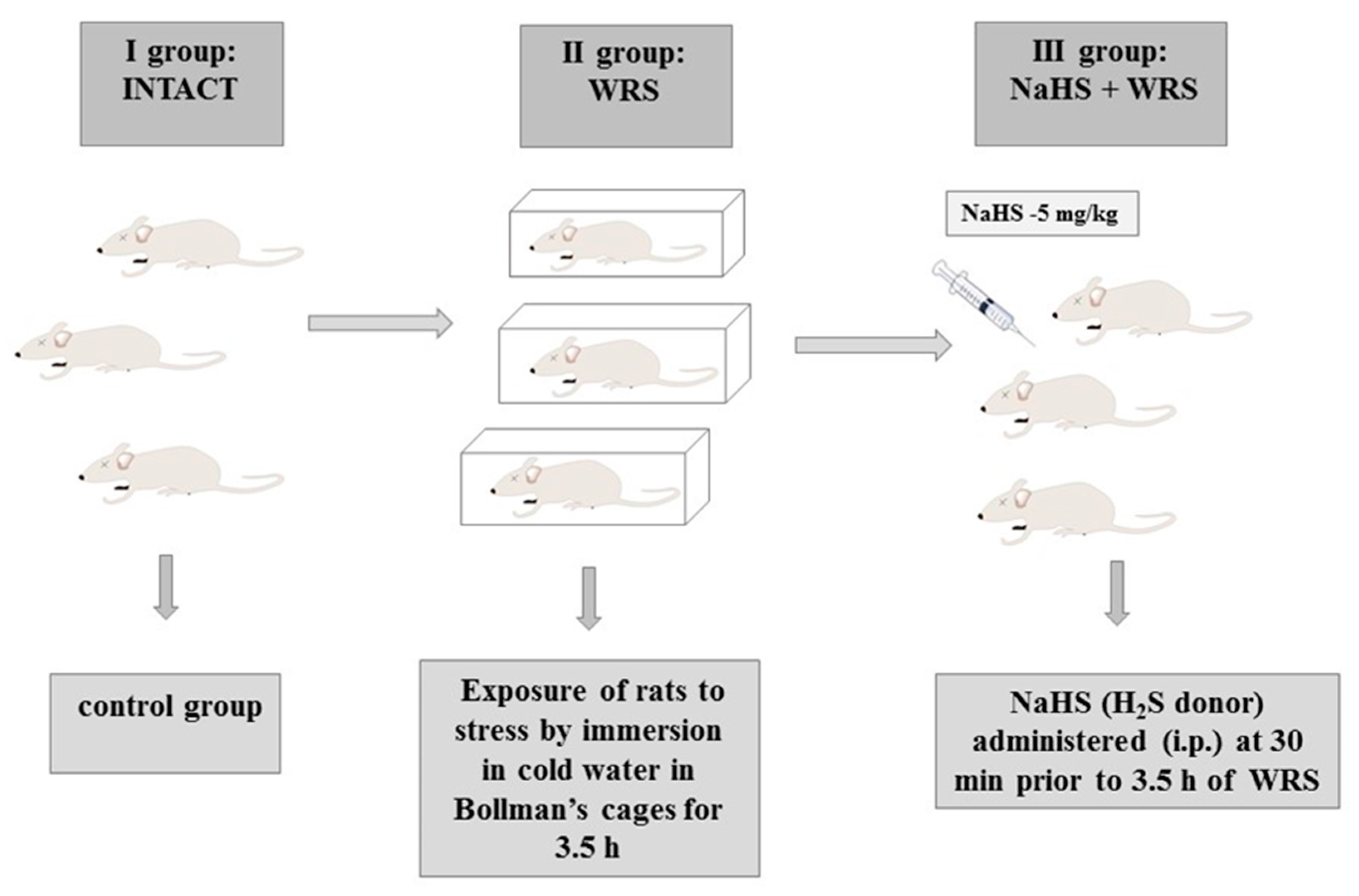
3.3. Experimental Group
3.4. Tissue Homogenates
3.5. RP-HPLC
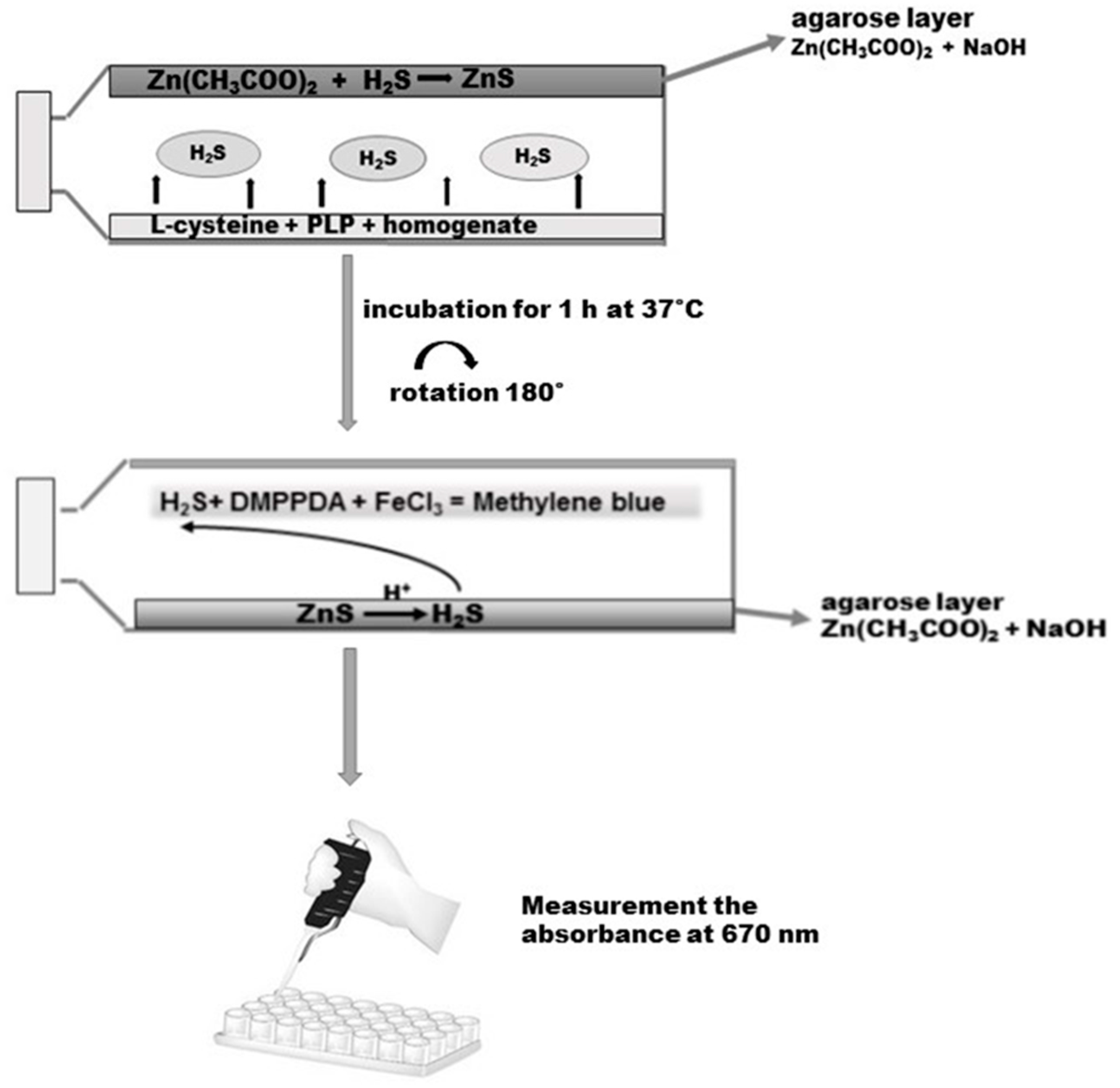
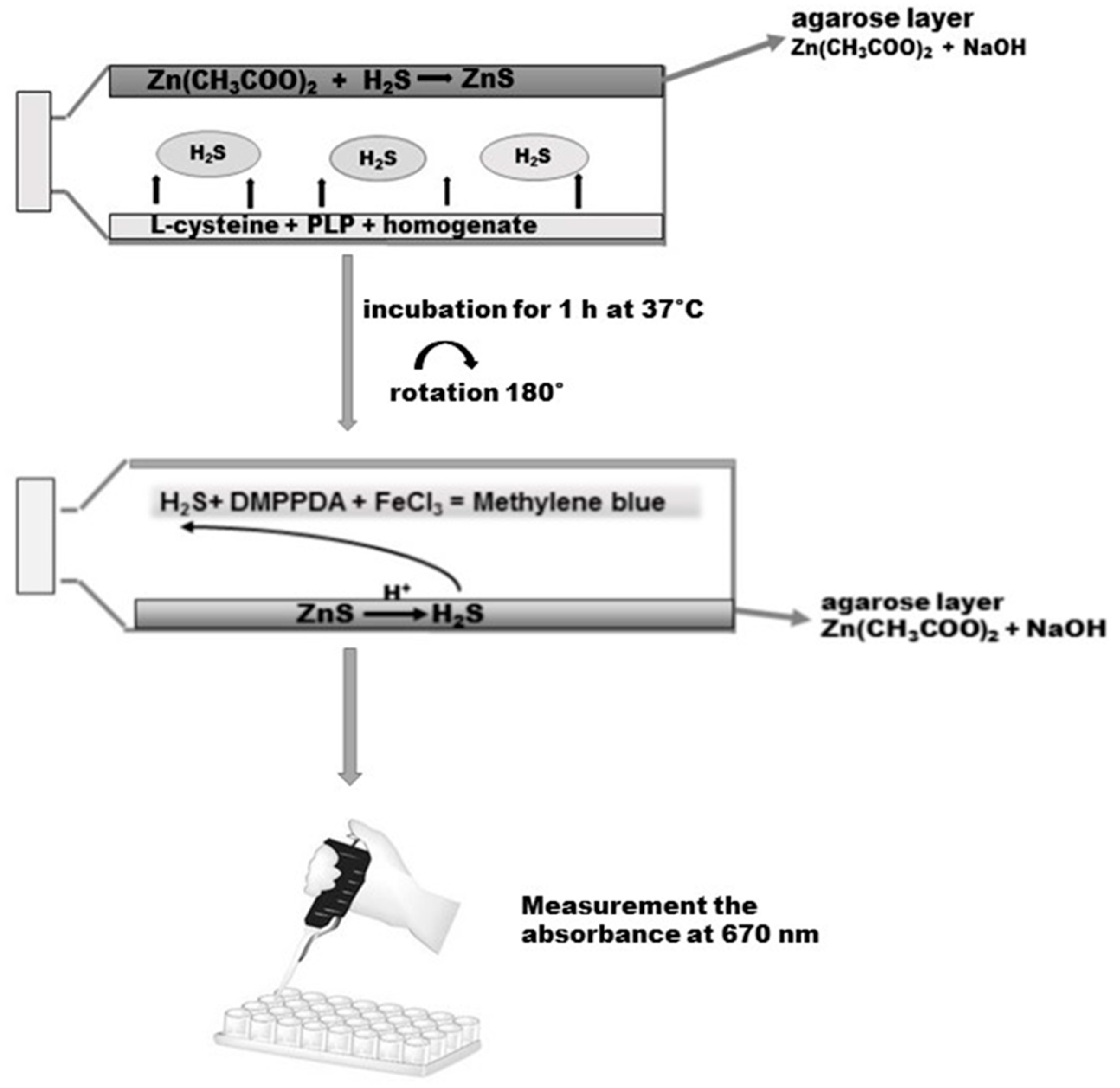
3.6. Determination of H2S in the Homogenate of the Gastric Mucosa
3.7. Enzymes Assay in the Gastric Mucosa Homogenates
3.8. Expression of MPST, CTH, and CBS in the Gastric Mucosa Homogenates
3.8.1. RNA Extraction
3.8.2. Reverse Transcription of RNA
3.8.3. cDNA Synthesis and RT-PCR Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stipanuk, M.H.; Ueki, I. Dealing with methionine/homocysteine sulfur: Cysteine metabolism to taurine and inorganic sulfur. J. Inherit. Metab. Dis. 2011, 34, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Vitvitsky, V.; Banerjee, R. Sulfur as a Signaling Nutrient through Hydrogen Sulfide. Annu. Rev. Nutr. 2014, 34, 171–205. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Bailey, S.M. Redox biology of hydrogen sulfide: Implications for physiology, pathophysiology, and pharmacology. Redox Biol. 2013, 1, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Mard, S.A.; Veisi, A.; Ahangarpour, A; Gharib-Naseri, M.K. Mucosal acidification increases hydrogen sulfide release through up-regulating gene and protein expressions of cystathionine gamma-lyase in the rat gastric mucosa. Iran. J. Basic Med. Sci. 2016, 19, 172–177. [Google Scholar] [PubMed]
- Tadeusiewicz, J.; Olas, B. Siarkowodór—Gaz nie tylko o właściwościach toksycznych. Kosmos 2014, 63, 125–135. [Google Scholar]
- Zaichko, N.V.; Melnik, A.V.; Yoltukhivsky, M.M.; Olhovskiy, A.S.; Palamarchuk, I.V. Hydrogen sulfide: Metabolism, biological and medical role. Ukr. Biochem. J. 2014, 86, 5–25. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Okabe, S. The effects of drugs on the production and recovery processes of the stress ulcer. Jpn. J. Pharmacol. 1968, 19, 9–19. [Google Scholar] [CrossRef]
- Lou, L.X.; Geng, B.; Du, J.B.; Tang, C.S. Hydrogen sulphide-induced hypothermia attenuates stress-related ulceration in rats. Clin. Exp. Pharmacol. Physiol. 2008, 35, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Dicay, M.; McKnight, W.; Martin, G.R. Hydrogen sulfide enhances ulcer healing in rats. FASEB J. 2007, 21, 4070–4076. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [PubMed]
- Toohey, J.I.; Cooper, A.J.L. Thiosulfoxide (Sulfane) Sulfur: New Chemistry and New Regulatory Roles in Biology. Molecules 2014, 19, 12789–12813. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.R.; McKnight, G.W.; Dicay, M.S.; Coffin, C.S.; Ferraz., J.G.; Wallace, J.L. Hydrogen sulphide synthesis in the rat and mouse gastrointestinal tract. Dig. Liver Dis. 2010, 42, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Magierowski, M.; Jasnos, K.; Kwiecień, S.; Brzozowski, T. Rola siarkowodoru w fizjologii przewodu pokarmowego i w mechanizmie gastroprotekcji. Postępy Hig. Med. Dośw. 2013, 67, 150–156. [Google Scholar] [CrossRef]
- Wallace, J.L. Physiological and Pathophysiological Roles of Hydrogen Sulfide in the Gastrointestinal Tract. Antioxid. Redox Signal. 2010, 9, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Toohey, J.L. Sulfur signaling: Is the agent sulfide or sulfane? Anal. Biochem. 2011, 413, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Czekaj, R.; Majka, J.; Ptak-Belowska, A.; Szlachcic, A.; Targosz, A.; Magierowska, K.; Strzalka, M.; Magierowski, M.; Brzozowski, T. Role of curcumin in protection of gastric mucosa against stress-induced gastric mucosal damage. Involvement of hypoacidity, vasoactive mediators and sensory neuropeptides. J. Physiol. Pharmacol. 2016, 67, 261–275. [Google Scholar] [PubMed]
- Magierowska, K.; Magierowski, M.; Surmiak, M.; Adamski, J.; Mazur-Biały, A.I.; Pajdo, R.; Sliwowski, Z.; Kwiecien, S.; Brzozowski, T. The protective role of carbon monoxide (CO) produced by heme oxygenases and derived from the CO-releasing molecule CORM-2 in the pathogenesis of stress-induced gastric lesions: Evidence for non-involvement of nitric oxide (NO). Int. J. Mol. Sci. 2016, 17, 442. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Meixner, H.; Ptak, A.; Hahn, E.G.; Konturek, S.J. Central and peripheral neural aspects of gastroprotective and ulcer healing effects of lipopolysaccharides. J. Physiol. Pharmacol. 2001, 52, 611–623. [Google Scholar] [PubMed]
- Dominik, P.K.; Cassidy, P.B.; Roberts, J.C. A new and versatile method for determination of thiolamines of biological importance. J. Chromatogr. B 2001, 761, 1–12. [Google Scholar] [CrossRef]
- Bronowicka-Adamska, P.; Wróbel, M.; Zagajewski, J. RP-HPLC method for quantitative determination of cystathionine, cysteine and glutathione: An application for the study of the metabolism of cysteine in human brain. J. Chromatogr. B 2011, 879, 2005–2009. [Google Scholar] [CrossRef] [PubMed]
- Bronowicka-Adamska, P.; Wróbel, M.; Zagajewski, J. An application of RP-HPLC for determination of the activity of cystathionine β-synthase and γ-cystathionase in tissue homogenates. Nitric Oxide 2015, 46, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Wróbel, M.; Lewandowska, I.; Bronowicka-Adamska, P.; Paszewski, A. The level of sulfane sulfur in the fungus Aspergillus nidulans wild type and mutant strains. Amino Acids. 2009, 37, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Kartha, R.V.; Zhou, J.; Hovde, L.B.; Belinda, W.Y.; Cheung, B.W.Y.; Schröder, H. Enhanced detection of hydrogen sulfide generated in cell culture using an agar trap method. Anal. Biochem. 2012, 423, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Valentine, W.N.; Frankelfeld, J.K. 3-Mercaptopyuruvate sulfurtransferase (EC 2.8.1.2): A simple assay adapted to human blood cells. Clin. Chim. Acta 1974, 14, 205–210. [Google Scholar] [CrossRef]
- Wróbel, M.; Jurkowska, H.; Sliwa, L.; Srebro, Z. Sulfurtransferases and cyanide detoxification in mouse liver, kidney, and brain. Toxicol. Mech. Methods 2004, 14, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Greenberg, D.M. A crystalline enzyme that cleaves homoserine and cystathionine. J. Biol. Chem. 1958, 230, 545–560. [Google Scholar] [PubMed]
- Czubak, J.; Wróbel, M.; Jurkowska, H. Cystathionine γ-lyase (EC 4.4.1.1): An enzymatic assay of α-ketobutyrate using lactate dehydrogenase. Acta Biol. Crac. Zool. 2002, 44, 113–117. [Google Scholar]
- Wood, L. Sulfane sulfur. Methods Enzymol. 1987, 43, 25–29. [Google Scholar]
- Sörbo, B. Rhodanese. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1955; Volume 2, pp. 334–337. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.I. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
Sample Availability: Not available. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bronowicka-Adamska, P.; Wróbel, M.; Magierowski, M.; Magierowska, K.; Kwiecień, S.; Brzozowski, T. Hydrogen Sulphide Production in Healthy and Ulcerated Gastric Mucosa of Rats. Molecules 2017, 22, 530. https://doi.org/10.3390/molecules22040530
Bronowicka-Adamska P, Wróbel M, Magierowski M, Magierowska K, Kwiecień S, Brzozowski T. Hydrogen Sulphide Production in Healthy and Ulcerated Gastric Mucosa of Rats. Molecules. 2017; 22(4):530. https://doi.org/10.3390/molecules22040530
Chicago/Turabian StyleBronowicka-Adamska, Patrycja, Maria Wróbel, Marcin Magierowski, Katarzyna Magierowska, Sławomir Kwiecień, and Tomasz Brzozowski. 2017. "Hydrogen Sulphide Production in Healthy and Ulcerated Gastric Mucosa of Rats" Molecules 22, no. 4: 530. https://doi.org/10.3390/molecules22040530