Germination under Moderate Salinity Increases Phenolic Content and Antioxidant Activity in Rapeseed (Brassica napus var oleifera Del.) Sprouts


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Germination and Seedling Growth
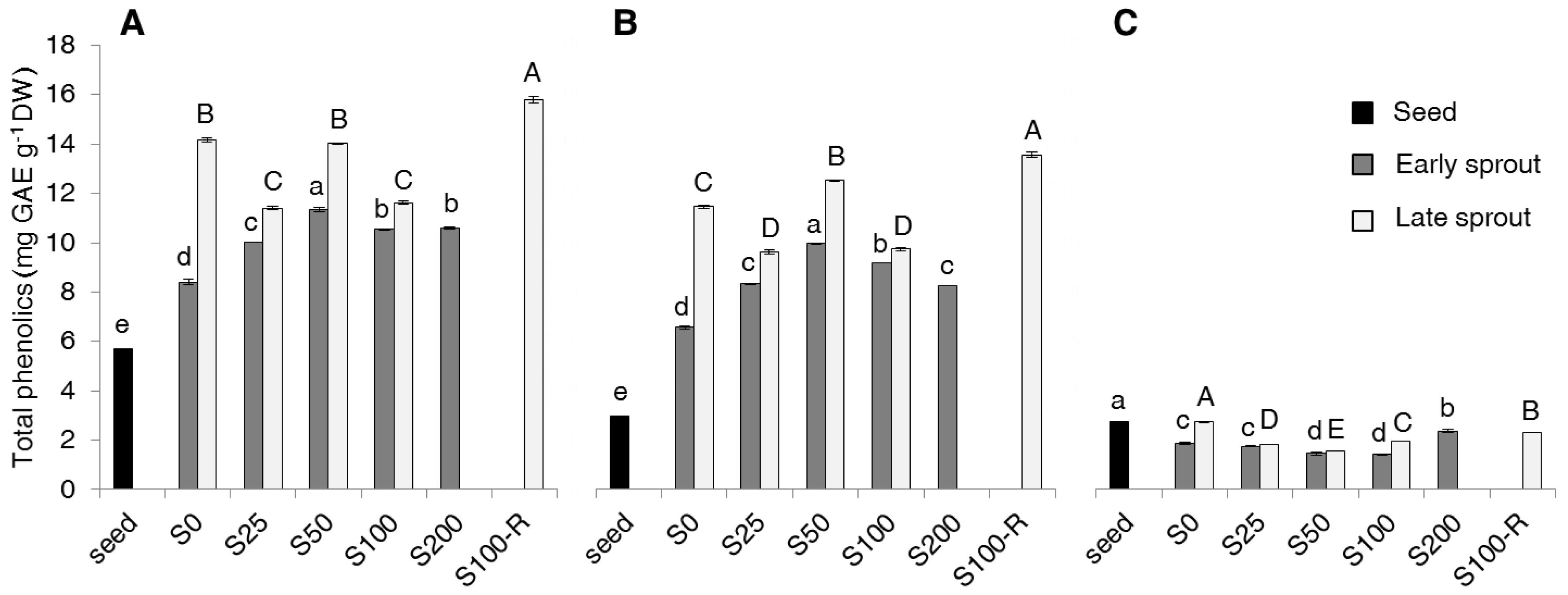
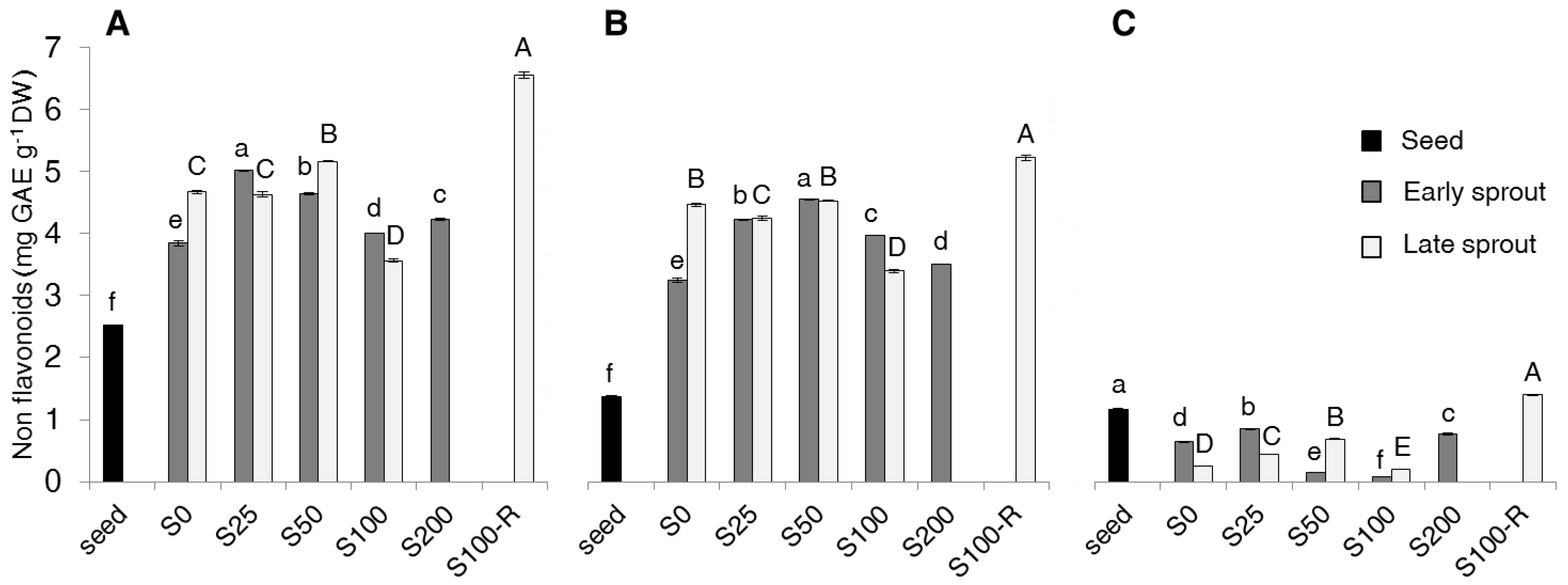
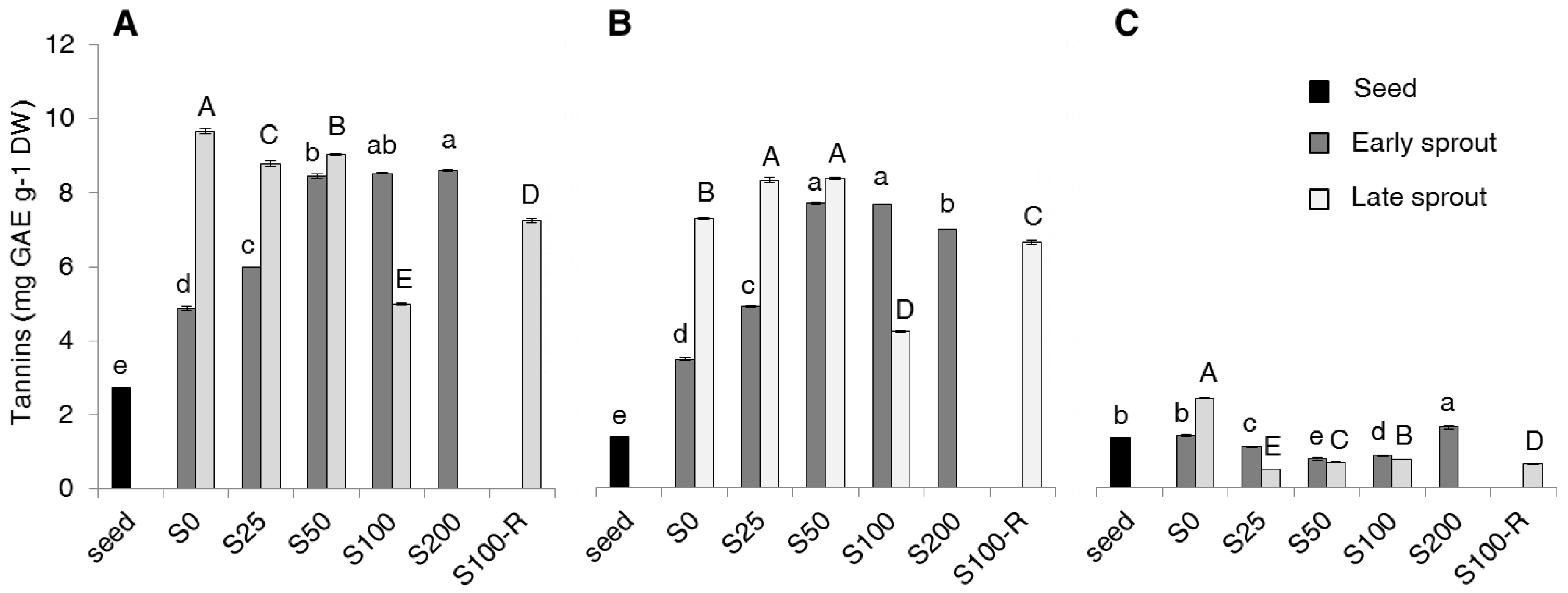
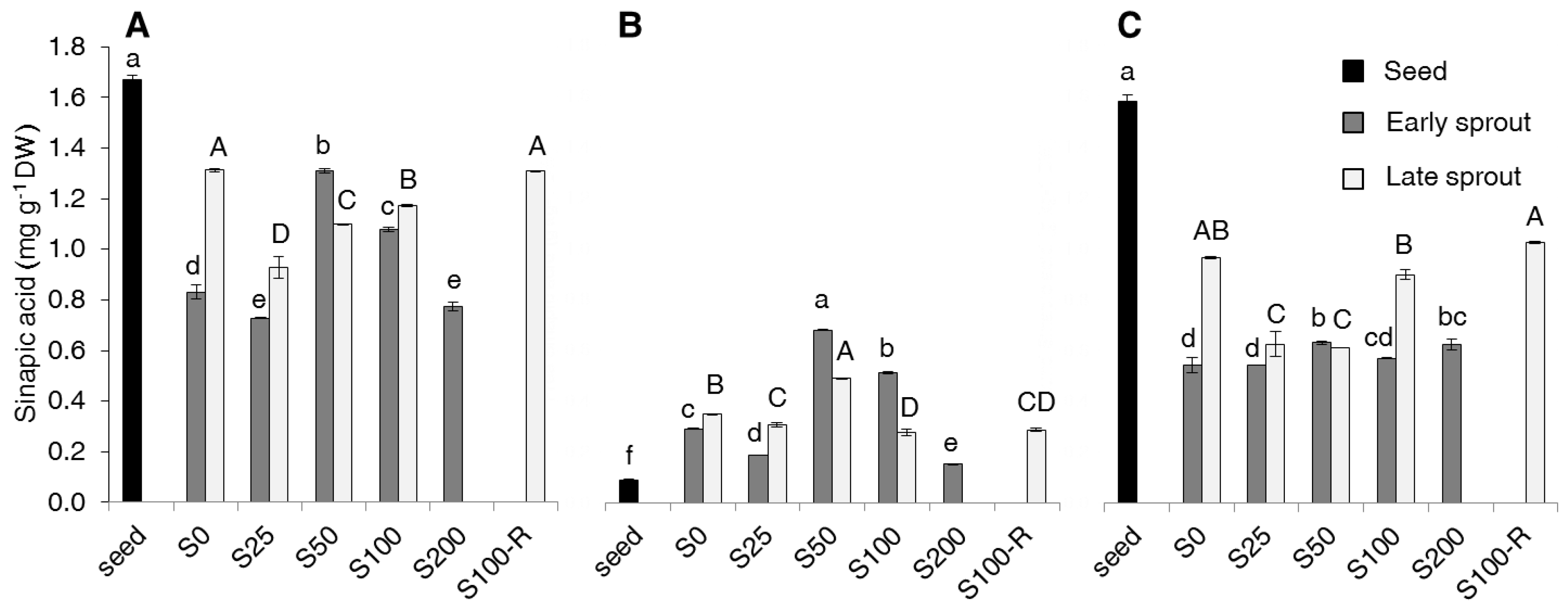
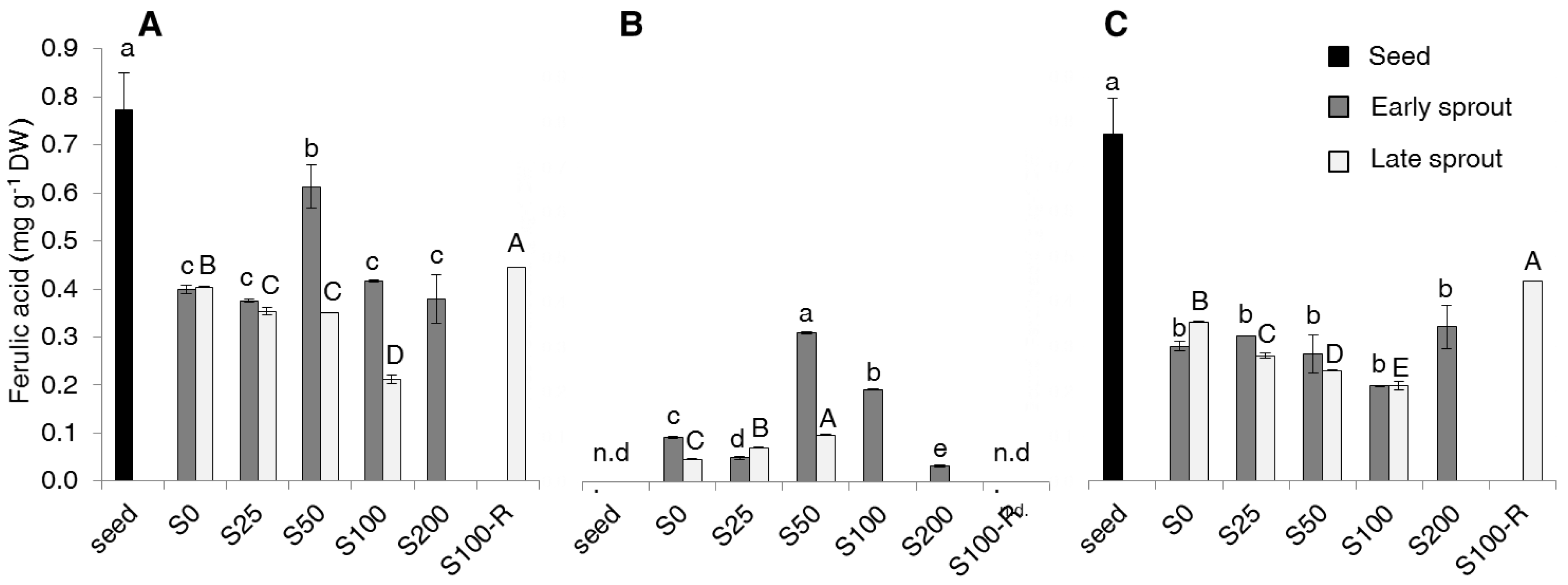
2.2. Contents of Total Phenolics, Non-Flavonoids, Tannins and PAs
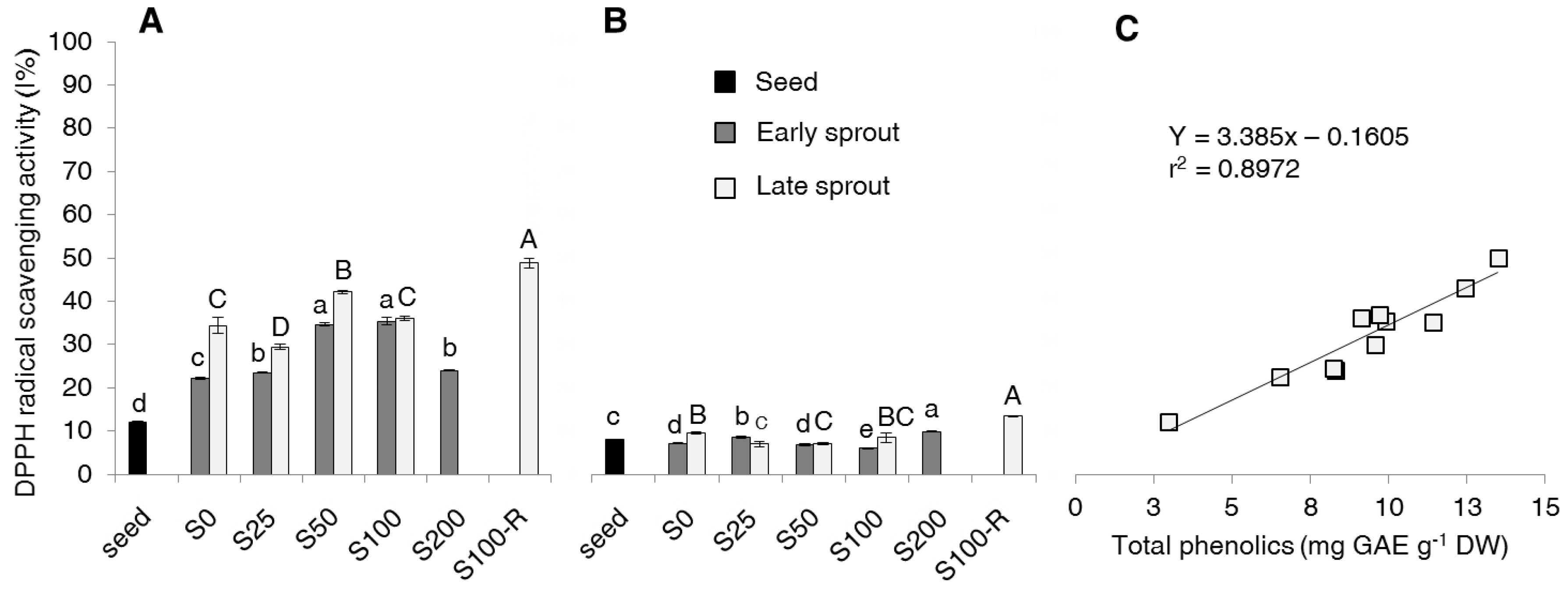
2.3. DPPH Scavenging Activity of Free and Bound Phenolic Fraction
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Chemicals and Apparatus
4.3. Extraction of Free and Bound Phenolic Fractions
4.4. Total Phenolics
4.5. Total Non-Flavonoid and Tannic Content
4.6. Analysis of Phenolic Acids Contents
4.7. DPPH Assay
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Màrton, M.; Màndoki, Z.; Csapó-Kiss, Z.; Csapó, J. The role of sprouts in human nutrition. A review. Acta Universitatis Sapientiae Alimentaria 2010, 3, 81–117. [Google Scholar]
- Baenas, N.; Moreno, D.A.; García-Viguera, C. Selecting sprouts of Brassicaceae for optimum phytochemical composition. J. Agric. Food Chem. 2012, 60, 11409–11420. [Google Scholar] [CrossRef] [PubMed]
- Moreno, D.A.; Carvajal, M.; López-Berenguer, C.; García-Viguera, C. Chemical and biological characterisation of nutraceutical compounds of broccoli. J. Pharm. Biomed. Anal. 2006, 41, 1508–1522. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica food: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Sauer, F.D.; Kramer, J.K.G. The problems associated with the feeding of high erucic acid rapeseed oils and some fish oils to experimental animals. In High and Low Erucic Acid Rapeseed Oils Production, Usage, Chemistry, and Toxicological Evaluation; Kramer, J.K.G., Sauer, F.D., Pgdeon, W.J., Eds.; Academic Press: San Diego, ON, USA, 1983; pp. 254–288. ISBN 0-12-425080-7. [Google Scholar]
- Felker, P.; Bunch, R.; Leung, A.M. Concentrations of thiocyanate and goitrin in human plasma, their precursor concentrations in Brassica vegetables, and associated potential risk for hypothyroidism. Nutr. Rev. 2016, 74, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Del Gatto, A.; Melilli, M.G.; Raccuia, S.A.; Pieri, S.; Mangoni, L.; Pacifico, L.; Signor, M.; Duca, D.; Foppa Pedretti, E.; Mengarelli, C. A comparative study of oilseed crops (Brassica napus L. subsp. oleifera and Brassica carinata A. Braun) in the biodiesel production chain and their adaptability to different Italian areas. Ind. Crops Prod. 2015, 75, 98–107. [Google Scholar] [CrossRef]
- Khanbabaee, K.; Van Ree, T. Tannins: Classification and definition. Nat. Prod. Rep. 2001, 18, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.A.; Beta, T.; Arntfield, S.D. A comparative study on the phenolic acids identified and quantified in dry beans using HPLC by different extraction and hydrolysis methods. Food Chem. 2009, 113, 336–344. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in food, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Pająk, P.; Socha, R.; Galkowska, D.; Rożnowski, J.; Fortuna, T. Phenolic profile and antioxidant activity in selected seeds and sprouts. Food Chem. 2014, 143, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Impact of germination on phenolic content and antioxidant activity of 13 edible seed species. Food Chem. 2010, 119, 1485–1490. [Google Scholar] [CrossRef]
- Zieliński, H.; Piskuła, M.K.; Kozłowska, H. Biologically active compounds in Cruciferae sprouts and their changes after thermal treatments. Pol. J. Food Nutr. Sci. 2005, 14, 375–380. [Google Scholar]
- Zieliński, H.; Piskuła, M.K.; Michalska, A.; Kozłowska, H. Antioxidant capacity and its components of cruciferous sprouts. Pol. J. Food Nutr. Sci. 2007, 57, 315–322. [Google Scholar]
- Pace, R.; Benincasa, P.; Ghanem, M.E.; Quinet, M.; Lutts, S. Germination of untreated and primed seeds in rapeseed (Brassica napus var. oleifera Del.) under salinity and low matric potential. Exp. Agric. 2012, 48, 238–251. [Google Scholar] [CrossRef]
- Benincasa, P.; Pace, R.; Quinet, M.; Lutts, S. Effect of salinity and priming on seedling growth in rapeseed (Brassica napus var oleifera Del.). Acta Sci. Agron. 2013, 35, 479–486. [Google Scholar] [CrossRef]
- Benincasa, P.; Galieni, A.; Manetta, A.C.; Pace, R.; Guiducci, M.; Pisante, M.; Stagnari, F. Phenolic compounds in grains, sprouts and wheatgrass of hulled and non-hulled wheat species. J. Sci. Food Agric. 2015, 95, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Turkan, I.; Demiral, T. Review: Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Gao, S.; Uuyang, C.; Wang, S.; Xu, Y.; Tang, L.; Chen, F. Effects of salt stress on growth, antioxidant enzyme and phenylalanine ammonia-lyase activities in Jatropha curcas L. seedlings. Plant Soil Environ. 2008, 58, 374–381. [Google Scholar]
- Lim, J.H.; Park, K.J.; Kim, B.K.; Jeong, J.W.; Kim, H.J. Effect of salinity stress on phenolic compounds and carotenoids in buckwheat (Fagopyrum esculentum M.) sprout. Food Chem. 2012, 135, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, A.; Muzolf-Panek, M.; Goliński, P. Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 283–314. [Google Scholar]
- Mert-Türk, F. Phytoalexins: Defence or just a response to stress? J. Cell. Biol. Mol. Sci. 2002, 1, 1–6. [Google Scholar]
- Yuan, G.; Wang, X.; Guo, R.; Wang, Q. Effect of salt stress on phenolic compounds, glucosinolates, myrosinase and antioxidant activity in radish sprouts. Food Chem. 2010, 121, 1014–1019. [Google Scholar] [CrossRef]
- Guo, L.; Yang, R.; Wang, Z.; Guo, Q.; Gu, Z. Effect of NaCl stress on health-promoting compounds and antioxidant activity in the sprouts of three broccoli cultivars. Int. J. Food Sci. Nutr. 2014, 65, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Khan, M.H. Growth, oxidative damage and antioxidant responses in greengram (Vigna radiata L.) under short-term salinity stress and its recovery. J. Agron. Crop Sci. 2009, 195, 442–454. [Google Scholar] [CrossRef]
- Ti, H.; Zhang, R.; Zhang, M.; Li, Q.; Wei, Z.; Zhang, Y.; Tang, X.; Deng, Y.; Liu, L.; Ma, Y. Dynamic changes in the free and bound phenolic compounds and antioxidant activity of brown rice at different germination stages. Food Chem. 2014, 161, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, S.; Udipi, S.A.; Ghugre, P. Polyphenols and tannins in Indian pulses: Effect of soaking, germination and pressure cooking. Food Res. Int. 2010, 43, 526–530. [Google Scholar] [CrossRef]
- Ghavidel, R.A.; Prakash, J. The impact of germination and dehulling on nutrients, antinutrients, in vitro iron and calcium bioavailability and in vitro starch and protein digestibility of some legume seeds. LWT Food Sci. Technol. 2007, 40, 1292–1299. [Google Scholar] [CrossRef]
- Ramakrishna, V.; Rani, P.J.; Rao, P.R. Anti-Nutritional factors during germination in Indian bean (Dolichos lablab L.) seeds. World J. Dairy Food Sci. 2006, 1, 6–11. [Google Scholar]
- Sangronis, E.; Machado, C.J. Influence of germination on the nutritional quality of Phaseolus vulgaris and Cajanus cajan. LWT Food Sci. Technol. 2007, 40, 116–120. [Google Scholar] [CrossRef]
- Rozema, J.; Van De Staaij, J.; Björn, L.O.; Caldwell, M. UV-B as an environmental factor in plant life: Stress and regulation. Trends Ecol. Evol. 1997, 12, 22–28. [Google Scholar] [CrossRef]
- Reinoso, H.; Sosa, L.; Ramírez, L.; Luna, V. Salt-induced changes in the vegetative anatomy of Prosopis strombulifera (Leguminosae). Can. J. Bot. 2004, 82, 618–628. [Google Scholar] [CrossRef]
- Odjegba, V.J.; Alokolaro, A.A. Simulated Drought and salinity modulates the production of phytochemicals in Acalypha wilkesiana. J. Plant Stud. 2013, 2, 105–112. [Google Scholar] [CrossRef]
- Szydlowska-Czerniak, A.; Trokowski, K.; Karlovits, G.; Szlyk, E. Determination of antioxidant capacity, phenolic acids, fatty acid composition of rapeseed varieties. J. Agric. Food Chem. 2010, 58, 7502–7509. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists (AOAC): Arlington, VA, USA, 1995. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with posphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Montedoro, G.; Fantozzi, P. Dosage des tanins dans les mouts et les vins a l’aide de la methylcellulose et evaluation d’autres fractions phenoliques. LWT Food Sci. Technol. 1974, 7, 155–171. (in French). [Google Scholar]
- Kramling, T.E.; Singleton, V.L. An estimate of the nonflavonoid phenols in wines. Am. J. Enol. Vitic. 1969, 20, 86–92. [Google Scholar]
- Floridi, S.; Montanari, L.; Marconi, O.; Fantozzi, P. Determination of free phenolic acids in wort and beer by coulometric array detection. J. Agric. Food Chem. 2003, 51, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Nencini, C.; Cavallo, F.; Capasso, A.; Franchi, G.G.; Giorgi, G.; Micheli, L. Evaluation of antioxidative properties of Allium species growing wild in Italy. Phytother. Res. 2007, 21, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org/ (accessed on 23 May 2017).
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | G (%) | T50 (d) | Early Sprouts | Late Sprouts | ||||
|---|---|---|---|---|---|---|---|---|
| DAS | FW (mg) | DW (mg) | DAS | FW (mg) | DW (mg) | |||
| S0 | 99 | 2.5 | 4 | 28.7(3.74) | 3.51(0.04) | 7 | 37.3(1.04) | 2.96(0.019) |
| S25 | 97 | 2.5 | 4 | 22.8(3.42) | 3.60(0.16) | 8 | 36.0(5.29) | 3.62(0.091) |
| S50 | 99 | 2.5 | 5 | 12.3(0.67) | 3.42(0.01) | 9 | 20.4(1.84) | 3.56(0.169) |
| S100 | 97 | 2.5 | 6 | 15.3(0.61) | 3.54(0.29) | 11 | 20.7(2.67) | 3.83(0.379) |
| S200 | 96 | 2.5 | 8 | 13.6(0.92) | 3.29(0.34) | - | - | - |
| S100-R | 98 | 2.5 | 6 | - | - | 9 | 23.2(2.39) | 3.95(0.152) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falcinelli, B.; Sileoni, V.; Marconi, O.; Perretti, G.; Quinet, M.; Lutts, S.; Benincasa, P. Germination under Moderate Salinity Increases Phenolic Content and Antioxidant Activity in Rapeseed (Brassica napus var oleifera Del.) Sprouts. Molecules 2017, 22, 1377. https://doi.org/10.3390/molecules22081377
Falcinelli B, Sileoni V, Marconi O, Perretti G, Quinet M, Lutts S, Benincasa P. Germination under Moderate Salinity Increases Phenolic Content and Antioxidant Activity in Rapeseed (Brassica napus var oleifera Del.) Sprouts. Molecules. 2017; 22(8):1377. https://doi.org/10.3390/molecules22081377
Chicago/Turabian StyleFalcinelli, Beatrice, Valeria Sileoni, Ombretta Marconi, Giuseppe Perretti, Muriel Quinet, Stanley Lutts, and Paolo Benincasa. 2017. "Germination under Moderate Salinity Increases Phenolic Content and Antioxidant Activity in Rapeseed (Brassica napus var oleifera Del.) Sprouts" Molecules 22, no. 8: 1377. https://doi.org/10.3390/molecules22081377