Effects of Refined Xiaoyaosan on Depressive-Like Behaviors in Rats with Chronic Unpredictable Mild Stress through Neurosteroids, Their Synthesis and Metabolic Enzymes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
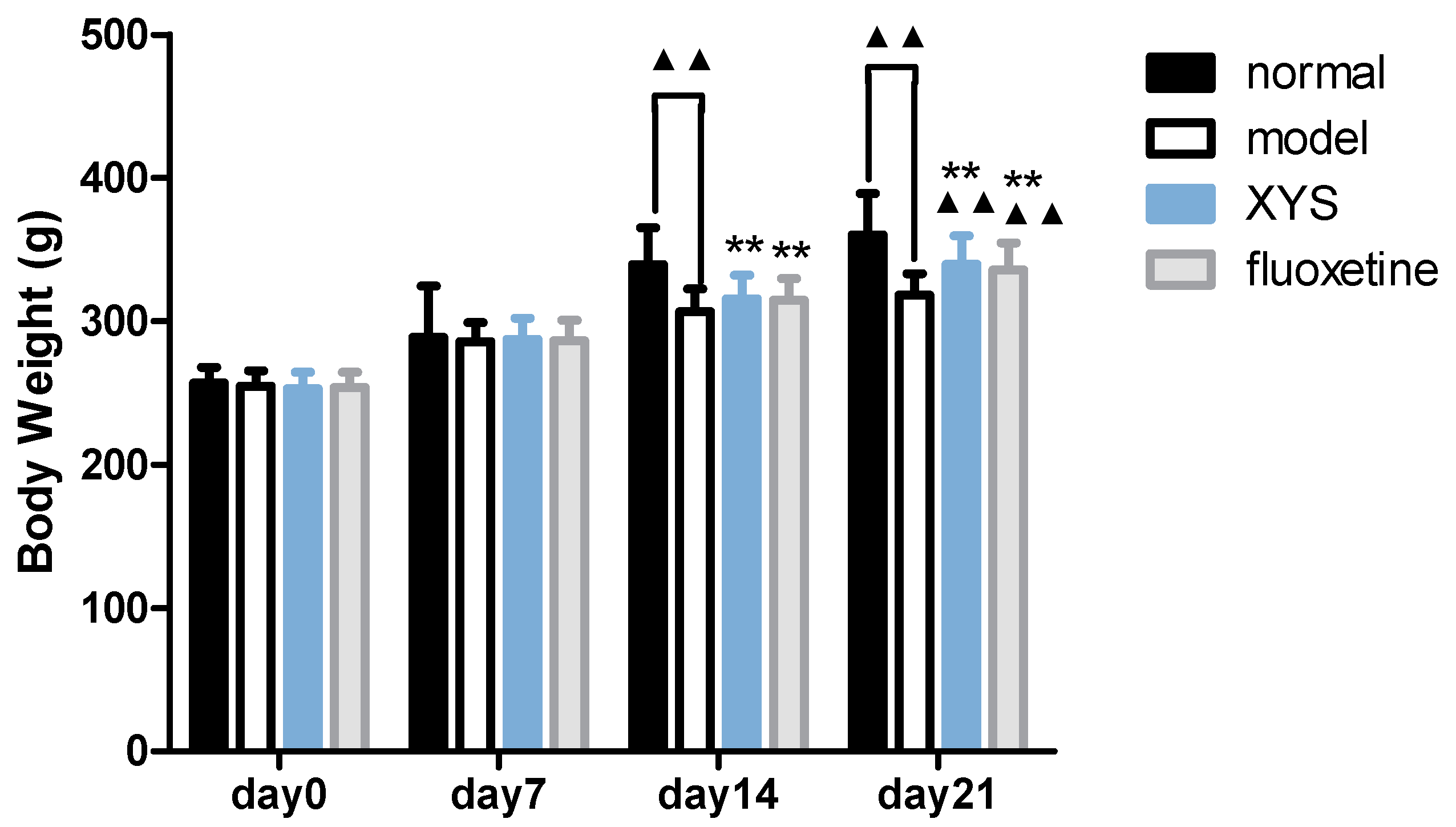
2.1. Effect of XYS on Body Weight of CUMS Rats
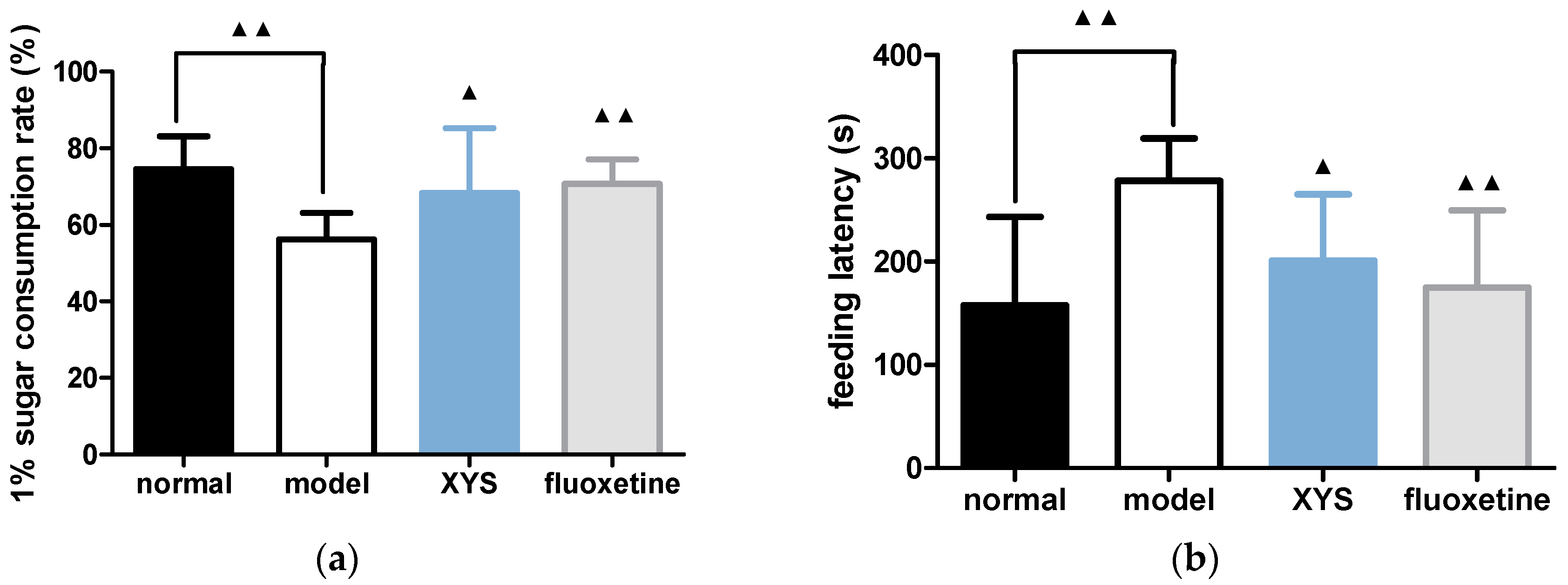
2.2. Effect of XYS on Sugar Consumption Rate and Novelty Suppressed Feeding Test of CUMS Rats
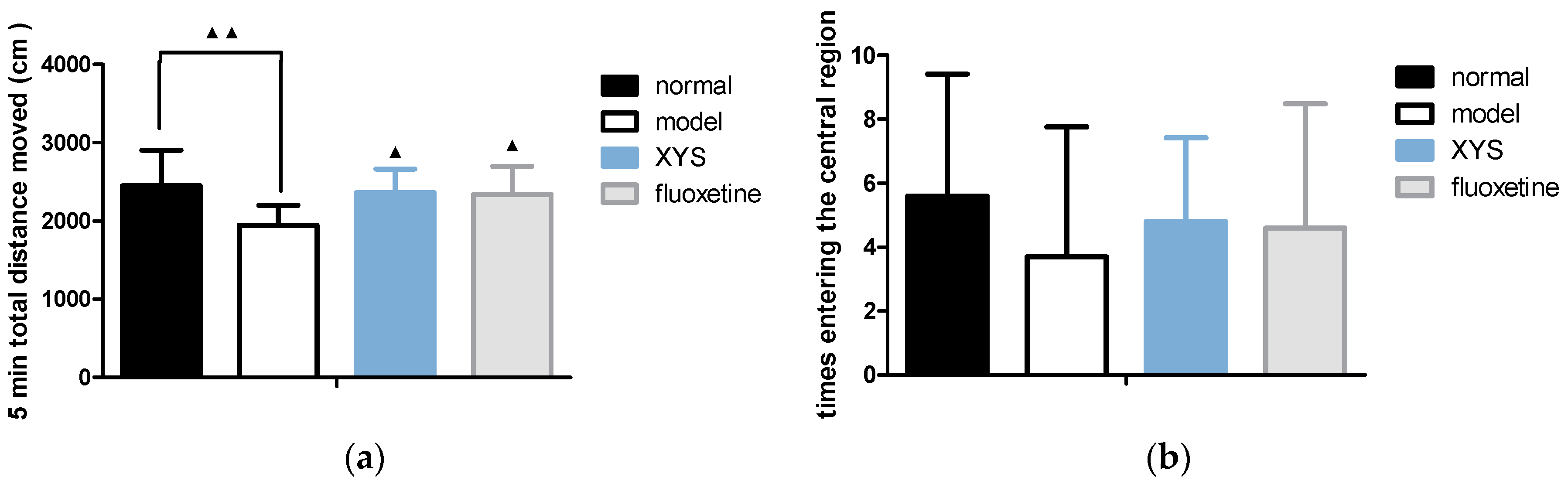
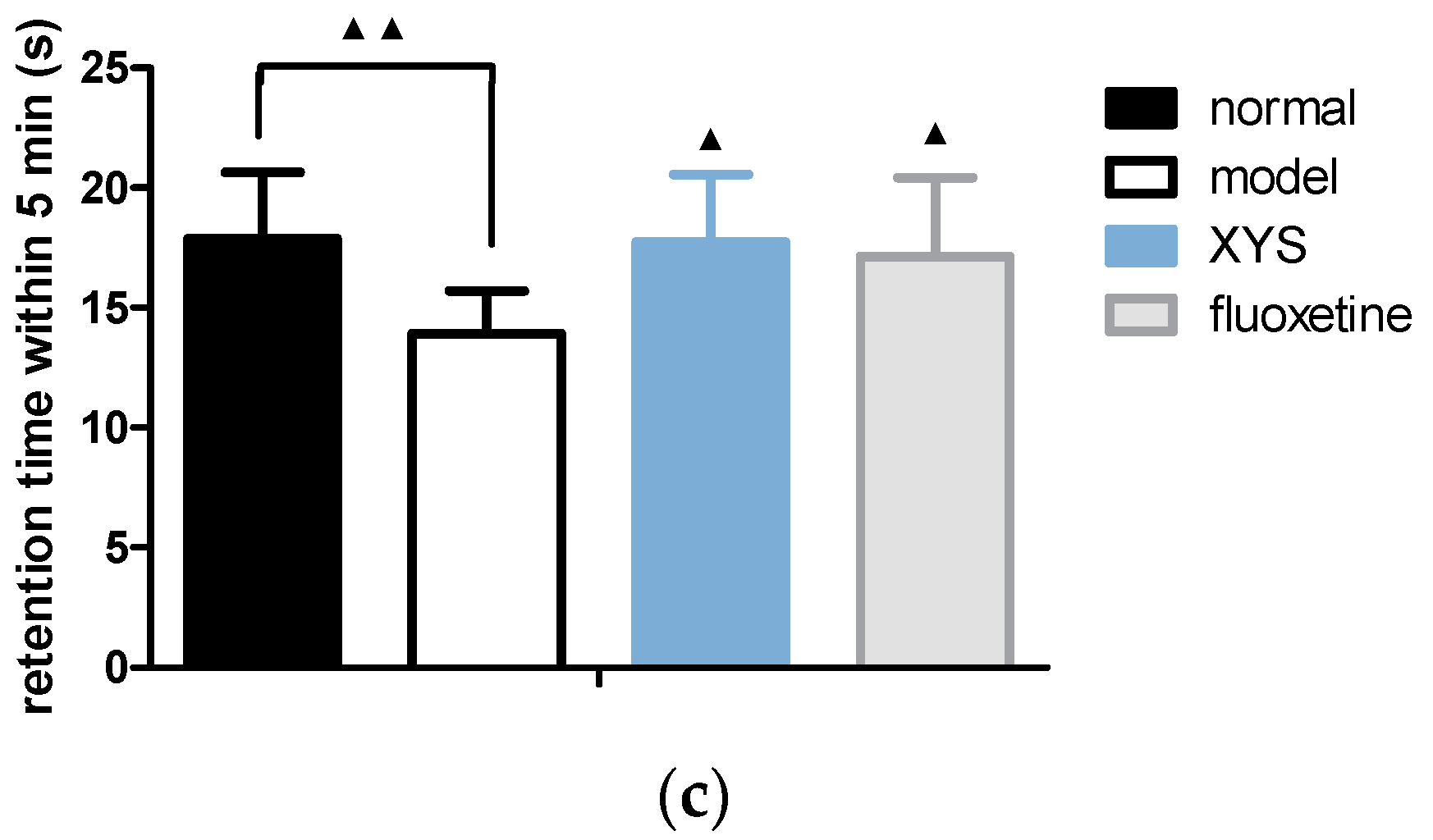
2.3. Effect of XYS on Open Field Test of CUMS Rats
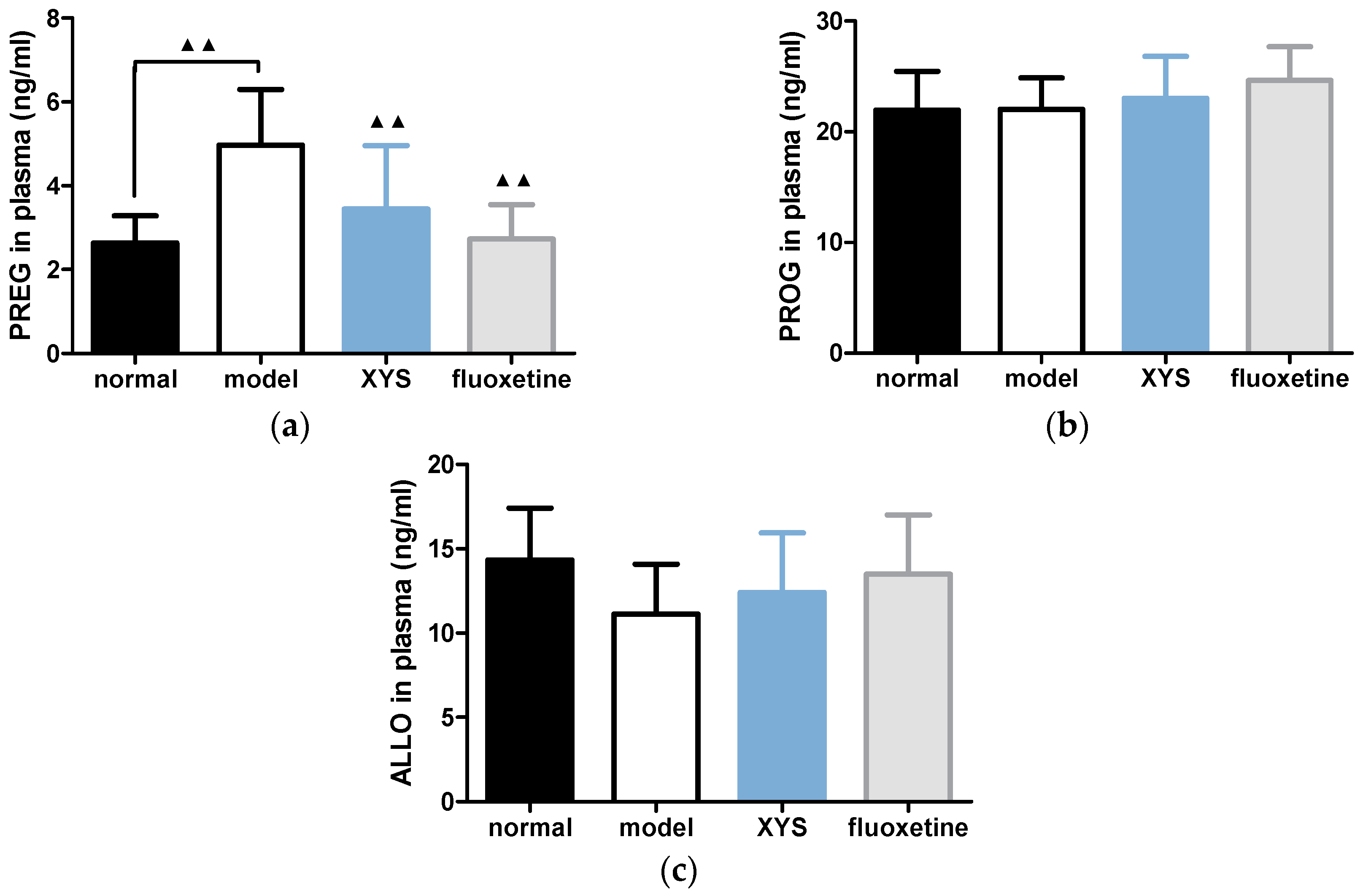
2.4. Effect of XYS on the Contents of PREG, PROG and ALLO in Plasma of CUMS Rats
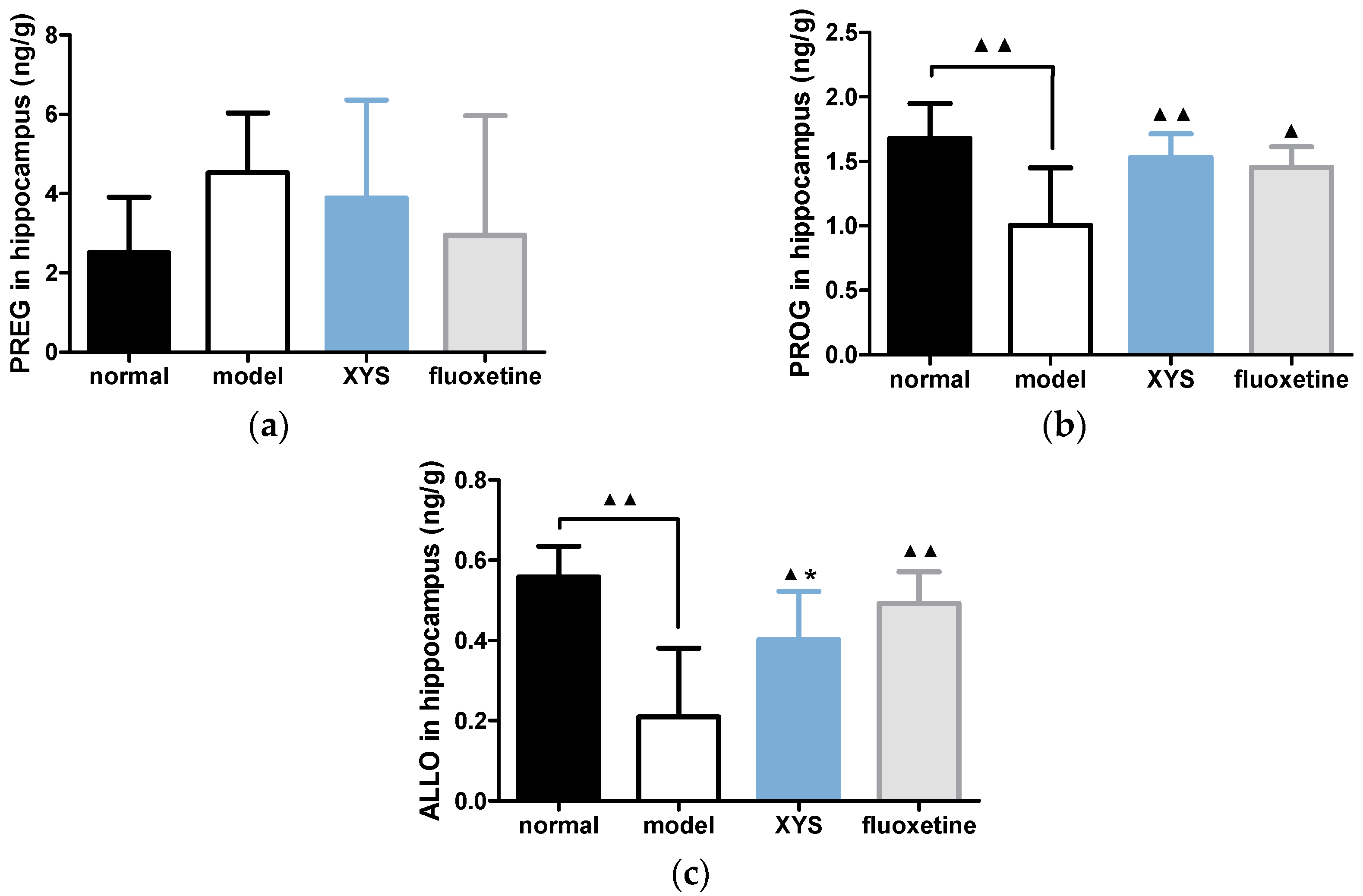
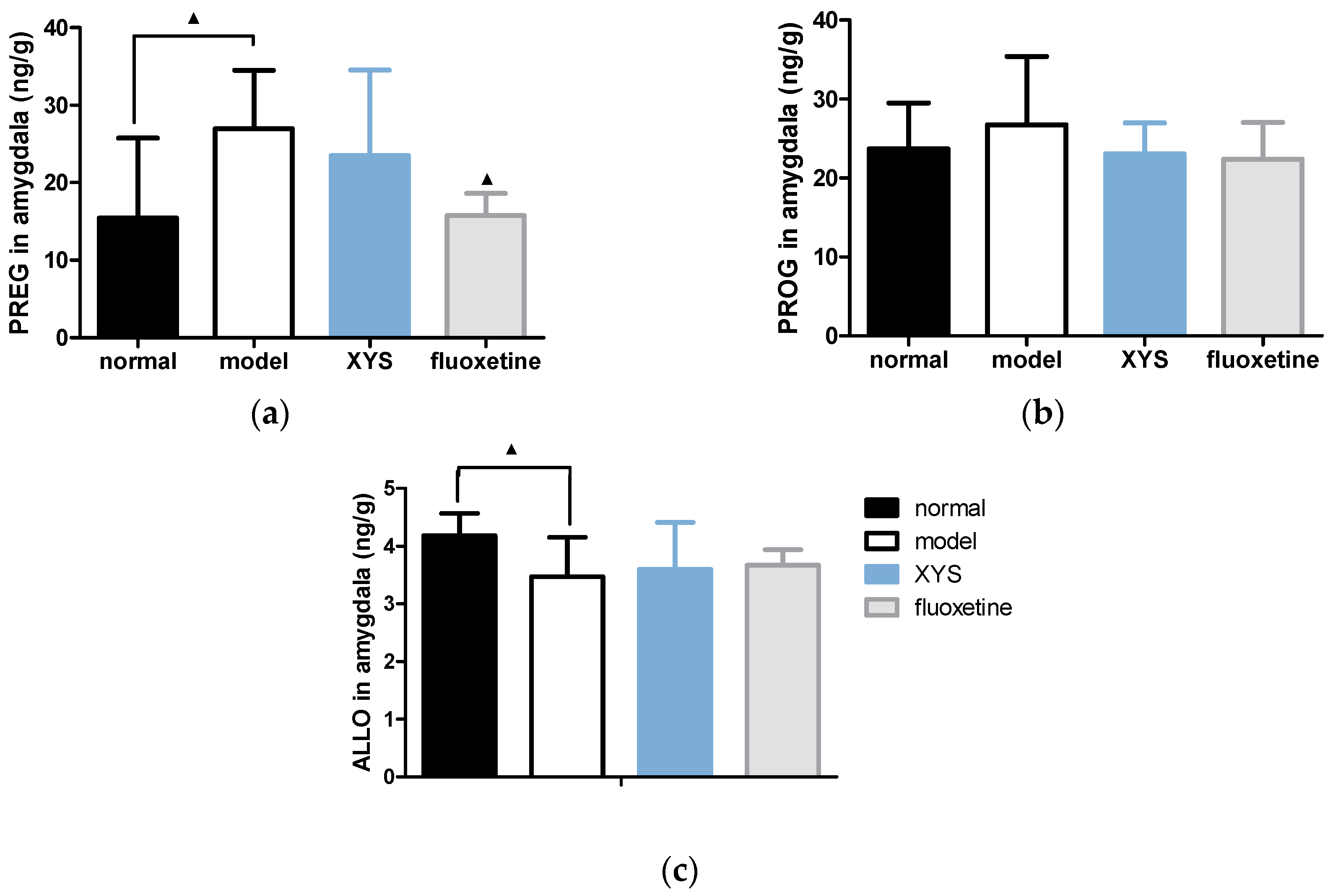
2.5. Effect of XYS on the Contents of PREG, PROG and ALLO in Brain of CUMS Rats
2.5.1. Effect of XYS on the Contents of PREG, PROG and ALLO in Hippocampus of CUMS Rats
2.5.2. Effect of XYS on the Contents of PREG, PROG and ALLO in Amygdala of CUMS Rats
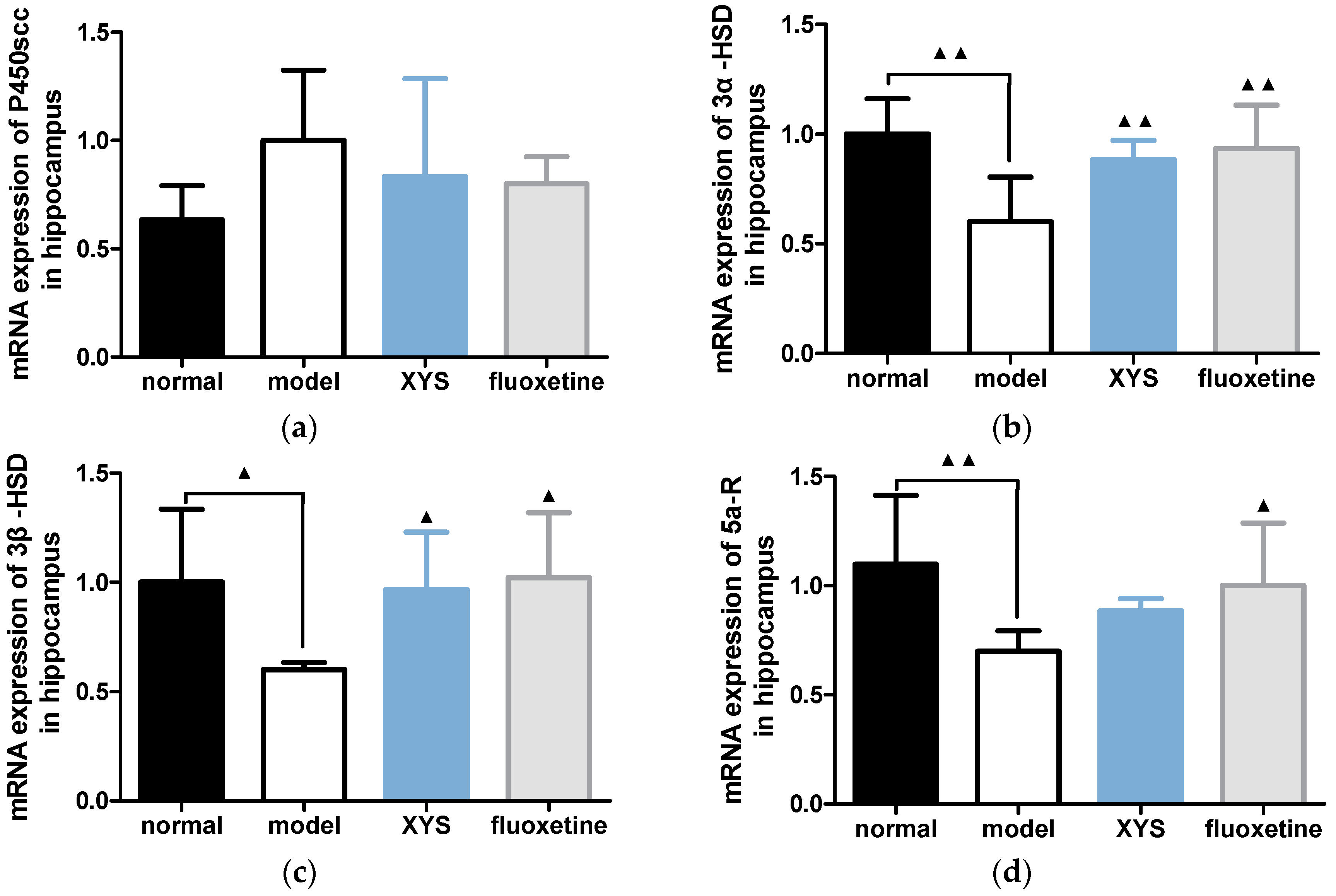
2.6. Effect of XYS on the mRNA Expressions of 3α-HSD, 3β-HSD, P450scc and 5α-R of CUMS Rats
2.6.1. Effect of XYS on the mRNA Expressions of 3α-HSD, 3β-HSD, P450scc and 5a-R in Hippocampus of CUMS Rats
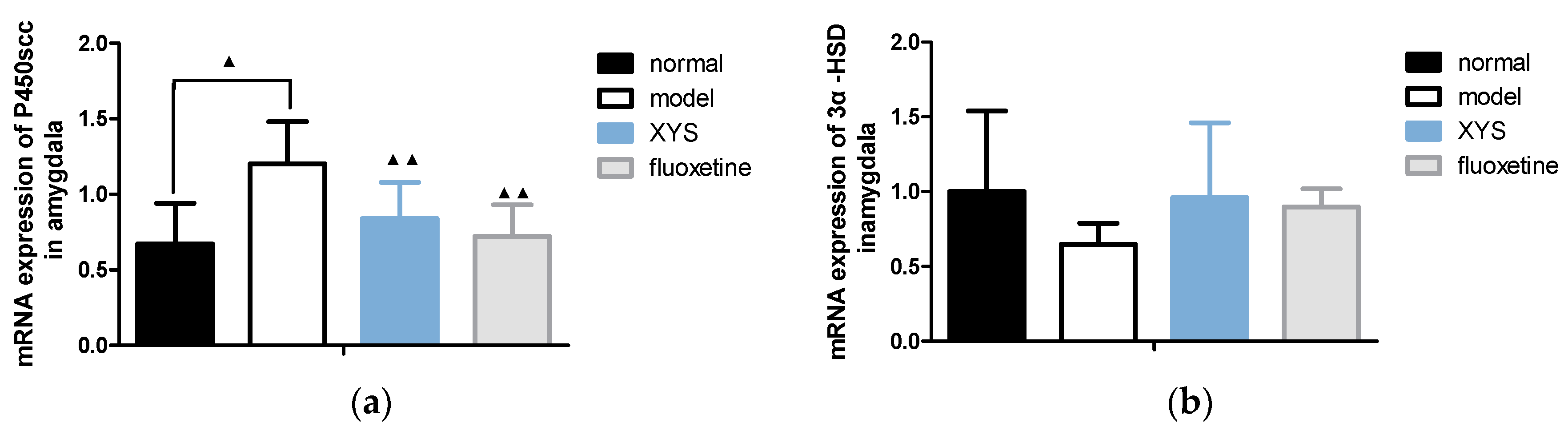
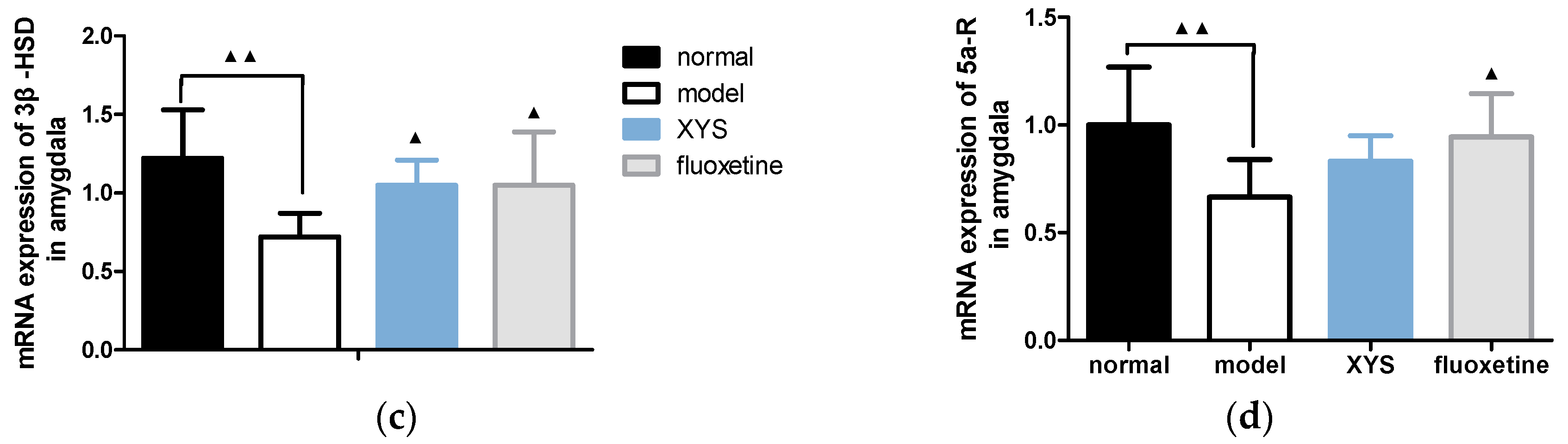
2.6.2. Effect of XYS on the mRNA Expressions of 3α-HSD, 3β-HSD, P450scc and 5a-R in Amygdala of CUMS Rats
3. Discussion
4. Materials and Methods
4.1. Animals and Grouping
4.2. Drug Preparation and Intervention
4.3. CUMS Model Preparation
4.4. Body Weight, Sugar Consumption Rate, Novelty Suppressed Feeding Test, Open Field Test
4.5. ELISA Analysis
4.6. LC-MS/MS Analysis
4.7. RT-qPCR Analysis
4.8. Statistical Methods
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zemlan, F.P.; Garver, D.L. Depression and antidepressant therapy: Receptor dynamics. Prog. Neuropsychopharmacol. Biol. Psychiatry 1990, 14, 503–523. [Google Scholar] [CrossRef]
- Belmaker, R.H.; Agam, G. Major depressive disorder. N. Engl. J. Med. 2008, 358, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.R.; Zhang, J.; Shi, Q.; Song, Z.; Ding, Z.; Pang, S.; Li, X.; Zhang, Y.; Wang, Z. Prevalence, treatment, and associated disability of mental disorders in four provinces in China during 2001–05: An epidemiological survey. Lancet 2009, 373, 2041–2053. [Google Scholar] [CrossRef]
- Berton, O.; Nestler, E.J. New approaches to antidepressant drug discovery: Beyond monoamines. Nat. Rev. Neurosci. 2006, 7, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Nissen, C.; Holz, J.; Blechert, J.; Feige, B.; Riemann, D.; Voderholzer, U.; Normann, C. Learning as a model for neural plasticity in major depression. Biol. Psychiat 2010, 68, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M. Why stress is bad for your brain. Science 1996, 273, 749–750. [Google Scholar] [CrossRef] [PubMed]
- Bremner, J.D. Does stress damage the brain? Biol. Psychiatry 1999, 45, 797–805. [Google Scholar] [CrossRef]
- Bowley, M.P.; Drevets, W.C.; Ongur, D.; Price, J.L. Low glial numbers in the amygdala in major depressive disorder. Biol. Psychiatry 2002, 52, 404–412. [Google Scholar] [CrossRef]
- Rajkowska, G. Depression: What we can learn from post-mortem studies. Neuroscientist 2003, 9, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P. Neuroactive steroid regulation of neurotransmitter release in the CNS: Action, mechanism and possible significance. Prog. Neurobiol. 2009, 89, 134–152. [Google Scholar] [CrossRef] [PubMed]
- Agis-Balboa, R.C.; Pinna, G.; Pibiri, F.; Kadriu, B.; Costa, E.; Guidotti, A. Down-regulation of neurosteroid biosynthesis in corticolimbic circuits mediates social isolation-induced behavior in mice. Proc. Natl. Acad. Sci. USA 2007, 104, 18736–18741. [Google Scholar] [CrossRef] [PubMed]
- Marx, C.E.; Stevens, R.D.; Shampine, L.J.; Uzunova, V.; Trost, W.T.; Butterfield, M.I.; Massing, M.W.; Hamer, R.M.; Morrow, A.L.; Lieberman, J.A. Neuroactive steroids are altered in schizophrenia and bipolar disorder: Relevance to pathophysiology and therapeutics. Neuropsychopharmacology 2006, 31, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Genud, R.; Merenlender, A.; Gispan-Herman, I.; Maayan, R.; Weizman, A.; Yadid, G. DHEA lessens depressive-like behavior via GABA-ergic modulation of the mesolimbic system. Neuropsychopharmacology 2009, 34, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.X.; Huang, Q.F.; Chen, J.X. Antidepressant effects of Xiaoyao powder and its different modified combinations. World J. Integr. Tradit. Western Med. 2007, 6, 326–328. [Google Scholar]
- Song, C.L.; Wang, H.W. Observation of curative effect of Xiaoyaosan on depression. J. Clin. Psychosom. Dis. 2007, 13, 54–57. [Google Scholar]
- Li, W.; Chen, J.X.; Yang, J.X. The effect of compounds of soothing liver, invigorating spleen, tonifying kidney on the praxeology and immunological function of chronic immobilization stressed rats. Acta Laboratorium Anim. Sci. Sin. 2003, 11, 251–252. [Google Scholar]
- Wang, Y.; Cui, X.L.; Liu, Y.F.; Gao, F.; Wei, D.; Li, X.W.; Wang, H.N.; Tan, Q.R.; Jiang, W. LPS inhibits the effects of fluoxetine on depression-like behavior and hippocampal neurogenesis in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1831–1835. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Li, M.X.; Xu, C.; Chen, H.B.; An, S.C.; Ma, X.M. Dendritic spines in depression: What we learned from animal models. Neural Plast 2016, 2016, 8056370. [Google Scholar] [CrossRef] [PubMed]
- Magarinos, A.M.; McEwen, B.S. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: Comparison of stressors. Neuroscience 1995, 69, 83–88. [Google Scholar] [CrossRef]
- Magarinos, A.M.; McEwen, B.S. Stress-induced atrophy of apical dendrites of hippocampal ca3c neurons: Involvement of glucocorticoid secretion and excitatory amino acid receptors. Neuroscience 1995, 69, 89–98. [Google Scholar] [CrossRef]
- Cao, X.; Li, L.P.; Wang, Q.; Wu, Q.; Hu, H.H.; Zhang, M.; Fang, Y.Y.; Zhang, J.; Li, S.J.; Xiong, W.C.; et al. Astrocyte-derived APT modulates depressive-like behaviors. Nat. Med. 2013, 19, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Jaggi, A.S.; Singh, N. Studies on effect of stress preconditioning in restraint stress-induced behavioral alterations. Yakugaku Zasshi 2010, 130, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.X.; Wang, Z.F.; Zhang, Q.L. Changes of AMPA receptors and related protein in immobilization stress rats and effect of Xiaoyao Powder. J. Beijing Univ. TCM 2007, 30, 603–607. [Google Scholar]
- Eser, D.; Schule, C.; Baghai, T.C.; Romeo, E.; Uzunov, D.P.; Rupprecht, R. Neuroactive steroids and affective disorders. Pharmacol. Biochem. Behav. 2006, 84, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Zhang, Y.F.; Liu, Y.Y. Effects of refined Xiaoyao Powder on behavior and protein expression of P450scc in hippocampus of depression rats. Chin. J. Tradit. Chin. Med. 2013, 2013, 1253–1258. [Google Scholar]
- Ma, K.; Xu, A.; Cui, S.; Sun, M.R.; Xue, Y.C.; Wang, J.H. Impaired GABA synthesis, uptake and release are associated with depression-like behaviors induced by chronic mild stress. Transl. Psychiatry 2016, 6, e910. [Google Scholar] [CrossRef] [PubMed]
- Pytka, K.; Dziubina, A.; Mlyniec, K.; Dziedziczak, A.; Zmudzka, E.; Furgala, A.; Olczyk, A.; Sapa, J.; Filipek, B. The role of glutamatergic, GABA-ergic, and cholinergic receptors in depression and antidepressant-like effect. Pharmacol. Rep. 2016, 68, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.X.; Chen, J.X.; Yue, G.X.; Bai, M.H.; Kou, M.J.; Jin, Z.Y. Xiaoyaosan decoction regulates changes in neuropeptide y and leptin receptor in the rat arcuate nucleus after chronic immobilization stress. Evid. Based Complement. Alternat. Med. 2012, 2012, 381278. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.Z.; Chen, J.X.; Jiang, Y.M.; Zhang, H.T. Effect of xiaoyaosan decoction on learning and memory deficit in rats induced by chronic immobilization stress. Evid. Based Complement. Alternat. Med. 2013, 2013, 297154. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, Q.; Li, X.J.; Bai, X.H.; Liu, Y.Y.; Zhao, H.B.; Jin, Z.Y.; Jing, Y.X.; Yan, Z.Y.; Chen, J.X. TCM formula Xiaoyaosan decoction improves depressive-like behaviors in rats with type 2 diabetes. Evid. Based Complement. Alternat. Med. 2015, 2015, 415243. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Lu, L.; Li, Z.; Gao, X.; Tian, J.; Zhang, L.; Wu, B.; Qin, X. Antidepressant-like effects of the fractions of Xiaoyaosan on rat model of chronic unpredictable mild stress. J. Ethnopharmacol. 2011, 137, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Katz, R.J.; Roth, K.A.; Carroll, B.J. Acute and chronic stress effects on open field activity in the rat: Implications for a model of depression. Neurosci. Biobehav. Rev. 1981, 5, 247–251. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Nicholson, J.K.; Hylands, P.J.; Sampson, J.; Holmes, E. A metabonomic strategy for the detection of the metabolic effects of chamomile (Matricaria recutita L.) ingestion. J. Agric. Food Chem. 2005, 53, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xue, R.; Yu, N.J. Effects of the total flavonoids of Xiaobuxin Decoction on the antidepressant effect of the acquired helpless mice and its effect on the HPA axis. Chin. J. Pharmacol. Bull. 2015, 2015, 815–821. [Google Scholar]
- Vogel, H.G.; Vogel, W.H. Pharmacological Experiment Guide—New Drug Discovery and Pharmacological Evaluation; Science Press: Beijing, China, 2001; pp. 309–310. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |









© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Qiu, W.; Liu, Y.; Zhang, Y.; Zhao, H.; Chen, J. Effects of Refined Xiaoyaosan on Depressive-Like Behaviors in Rats with Chronic Unpredictable Mild Stress through Neurosteroids, Their Synthesis and Metabolic Enzymes. Molecules 2017, 22, 1386. https://doi.org/10.3390/molecules22081386
Guo X, Qiu W, Liu Y, Zhang Y, Zhao H, Chen J. Effects of Refined Xiaoyaosan on Depressive-Like Behaviors in Rats with Chronic Unpredictable Mild Stress through Neurosteroids, Their Synthesis and Metabolic Enzymes. Molecules. 2017; 22(8):1386. https://doi.org/10.3390/molecules22081386
Chicago/Turabian StyleGuo, Xiaoling, Wenqi Qiu, Yueyun Liu, Yifang Zhang, Hongbo Zhao, and Jiaxu Chen. 2017. "Effects of Refined Xiaoyaosan on Depressive-Like Behaviors in Rats with Chronic Unpredictable Mild Stress through Neurosteroids, Their Synthesis and Metabolic Enzymes" Molecules 22, no. 8: 1386. https://doi.org/10.3390/molecules22081386