Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice

College of Horticulture, China Agricultural University, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Molecules 2018, 23(3), 521; https://doi.org/10.3390/molecules23030521
Submission received: 12 January 2018
/
Revised: 21 February 2018
/
Accepted: 23 February 2018
/
Published: 27 February 2018
(This article belongs to the Special Issue Melatonin as an Antioxidant and a Functionally Pleiotropic Molecule: Synthesis, Metabolism and Activities in Organisms)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Synthetic melatonin (N-acetyl-5-methoxytryptamine, MT) is popular in the US and Asian markets as a health supplement. Here, we identified a naturally occurring melatonin source in apple juice. Melatonin was present in all 18 apple cultivars tested. The highest melatonin level of the edible part of apple was detected in the apple peel. The melatonin content in ‘Fuji’ apple juice is comparable to the level of its flesh. Melatonin was consumed during the process of juicing due to its interaction with the oxidants. Melatonin addition significantly reduced the juice color change to brown (browning). The mechanism is that melatonin scavenges the free radicals, which was indicated by the ASBT analysis; therefore, inhibiting the conversion of o-diphenolic compounds into quinones. Most importantly, melatonin exhibited powerful anti-microorganism activity in juice. The exact mechanisms of this action are currently unknown. These effects of melatonin can preserve the quality and prolong the shelf life of apple juice. The results provide valuable information regarding commerciall apple juice processing and storage.
1. Introduction
Apple is one of the top four favorite fruits in the world. A total of 63,407,407 metric tons of apple worth approximately $23.5 billion are consumed yearly [1,2,3]. Apple is very popular as a healthy fruit and it contains multiple minerals, vitamins, and phytochemicals, including carotenoids, flavonoids, polyphenolic compounds, and phenolic acids [4,5,6,7,8,9,10]. Apple juices are also important dietary sources of these bioactive compounds [10,11]. It was reported that consuming apples and apple juice effectively reduces the risk of several cancers, cardiovascular disease, and asthma [12]. However, the underlying mechanisms remain elusive. Since melatonin is a potent antioxidant, in the current study, the antioxidant capacity of melatonin in apple and apple juice was systemically investigated.
Melatonin was first identified in plants in 1995; thereafter, it was found that melatonin existed in variety of plants and plant products [13,14,15,16,17,18]. In 2013, we firstly reported that melatonin was also present in apple fruit [19]. As a potent reactive oxygen species (ROS) and antioxidant, melatonin may be a major contributor regarding the health effects of apple, including its antioxidant, anticancer, antitumor, anti-inflammatory, anti-aging, anti-diabetic, antiviral and neuroprotective activities [20]. It also has therapeutic potential for respiratory, colonic, autoimmune, adult-onset chronic non-communicable diseases, ischemic brain and cardiovascular injury, circadian disturbance, stroke, steatohepatitis, etc. [21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41]. Melatonin can efficiently scavenge a variety of ROS including hydrogen peroxide (H2O2), hydroxyl radical (HO•), nitric oxide (NO•), singlet oxygen (1O2) and superoxide anion (O2–•) [42]. Compared with other classical antioxidants such as vitamin C, vitamin E and glutathione, melatonin is amphiphilic and can freely diffuse into different organelles. In addition, its metabolites, such as N1-acetyl-N2-formyl-5-methoxykynuramine (AFMK), N-acetyl-5-methoxykynuramine (AMK) and 6-hydroxymelatonin also possess the antioxidant activities, which amplify the capacity of melatonin as an antioxidant. The collective effects of melatonin and its metabolites are referred as a cascade reaction of melatonin [42,43,44,45,46,47].
Compared to synthetic melatonin, phyto-melatonin [48] may be more welcome by consumers. One of the natural melatonin sources are the fruits and their juices. Compared to the popularity of apple and its juice there is little information regarding the melatonin content in apple fruit of different cultivars and their juice. Especially, in ‘Fuji’ fruits, melatonin may be lost during the processing procedure or be consumed by the exposure to oxidants in air. No report is available on the natural melatonin level left after apple juicing. Therefore, it is difficult to know whether exogenous melatonin needs to be added to preserve the quality of the juice.
During the juicing process, o-diphenolic compounds are converted to quinones in the damaged cells and this results in the color of juice changing to brown. Inhibition of this oxidative reaction reduces the juice browning. Numerous studies on reducing color changes in apple juice have been reported [49,50,51,52,53,54]. Inhibitors of apple juice browning are divided into six groups: reducing molecules, chelating agents, complexing substances, acidulants, enzyme inhibitors, and enzyme treatments [49,55,56]. However, very few of these inhibitors are practically used in the food industry due to concerns about off-flavour and odours, food safety, and economic feasibility [57]. Therefore there is an increasing interest in identifying additional inhibitors which are safe, cheap, and abundant to enhance the stability of juice which is susceptible to browning.
As a powerful antioxidant, melatonin may be a first choice for healthy food supplements. In the current study, ‘Fuji’ apple was selected for apple juicing, since in China 70% of the apple production is ‘Fuji’ [58]. Melatonin levels in the apple peel and flesh of 18 cultivars were tested and also exogenous melatonin was supplemented to ‘Fuji’ apple juice during its processing to explore its ability to reduce the browning and inhibit microbial growth. Its antioxidant capacity was also checked. Our research should contribute to the application of melatonin as an anti-browning molecule and health care component in the apple juice industry.
2. Results
2.1. The Content of Melatonin and Total Phenols in ‘Fuji’ and ’Granny Smith’ Apple Fruit
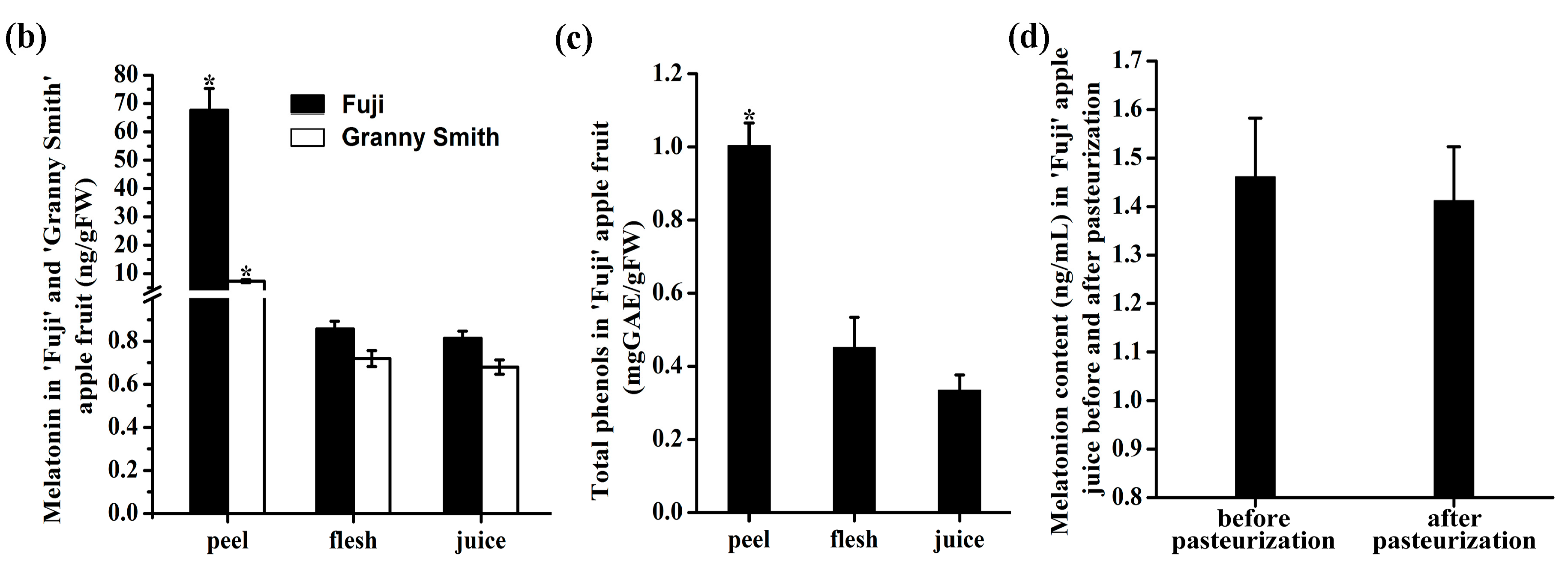
All the juice melatonin comes from the flesh since the melatonin from the peel may not be incorporated into the juice. The results showed that the highest melatonin level was in the peel of ‘Fuji’ apple (67.627 ng/gFW) and ’Granny Smith’ apple (7.37 ng/gFW), which was nearly 79- and 10- fold higher than the levels in their flesh (0.857 ng/gFW and 0.719 ng/gFW) and 83-times and 11-fold higher than that in their juices (0.814 ng/gFW and 0.680 ng/gFW), respectively (Figure 1b).
The content of total phenols in the ‘Fuji’ apple peel (1.00 mgGAE/gFW) was nearly 2-times and 3-fold higher than that in their flesh (0.45 mgGAE/gFW) and fruit juice (0.33 mgGAE/gFW), respectively (Figure 1c). Obviously, both the melatonin and total phenol in apple peel were significantly higher than that in their flesh and juice, but there was no significant difference in the melatonin and total phenol content between the flesh and fruit juice.
2.2. The Effect of Pasteurization on the Melatonin Content in Apple Juice
Pasteurization seems not influence melatonin levels in apple juice. Its levels were 1.46 and 1.41 ng/mL before and after pasteurization, respectively. There was no significant difference (p < 0.05) (Figure 1d).
2.3. The Melatonin Content in Different Apple Cultivars
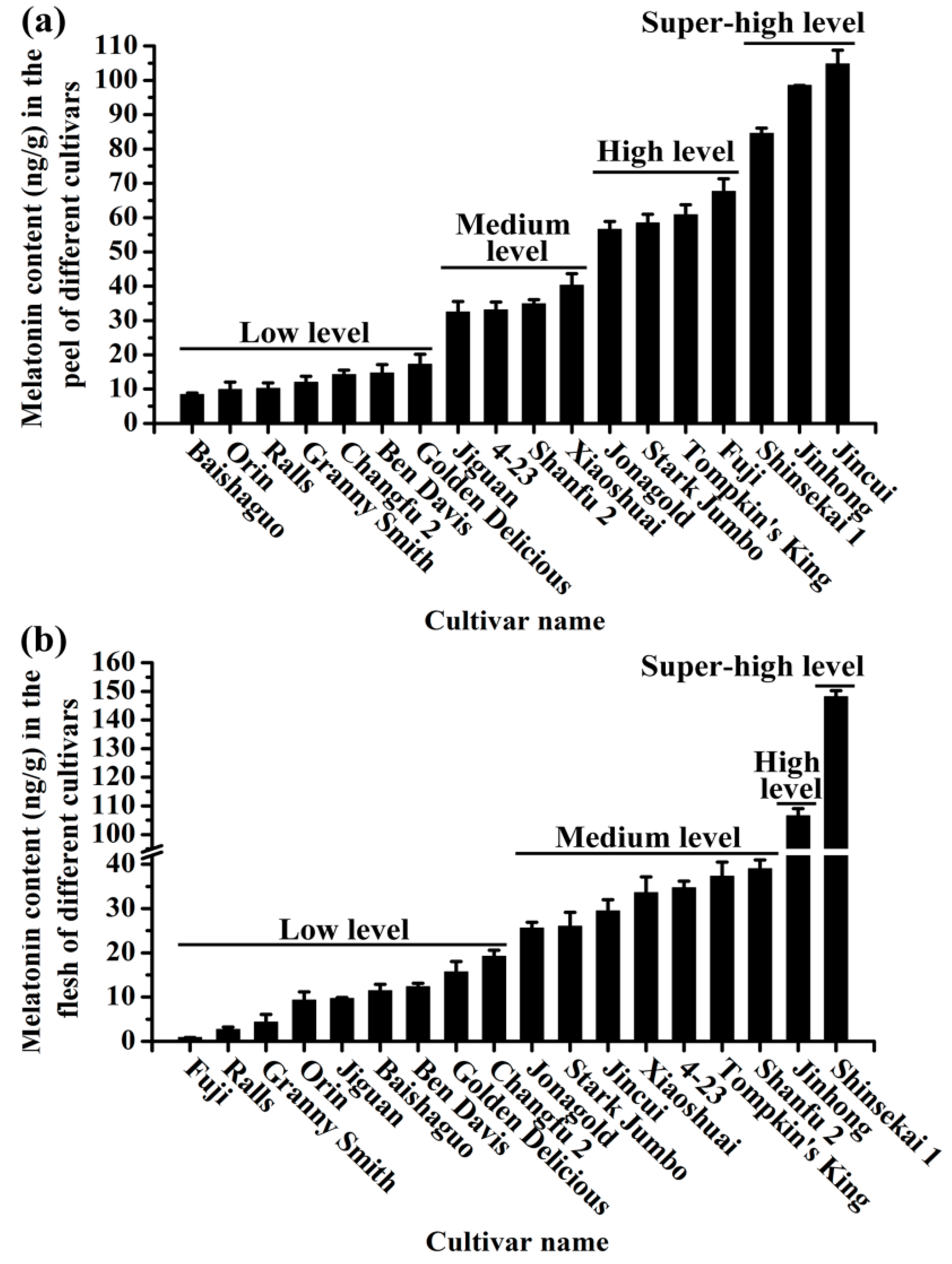
The melatonin contents in apple peel and flesh among the 18 commercial cultivars varied from 0.86 ng/g to 148.11 ng/g (Figure 2). For most of the cultivars, there was a good correlation between the melatonin in the peel and the flesh. If there was a high level in the peel, generally speaking, there was a high level in the flesh.
Based on the melatonin levels in apple peel, these cultivars were divided into four groups including super-high, high, medium and low level groups (Table 1 in the Supplementary Data). The cultivars ‘Baishaguo’, ‘Orin’, ‘Ralls’, ‘Granny Smith’, ‘Changfu 2’, ‘Ben Davis’ and ‘Golden Delicious’ with melatonin ranging from 8.37 to 17.18 ng/g were classified as the low melatonin group; ‘Jiguan’, ‘Shanfu 2’, ‘4-23’ and ‘Xiaoshuai’ with melatonin levels from 32.43 to 40.3 ng/g were the medium level group; ‘Jonagold’, ‘Stark Jumbo’, ‘Tompkin’s King’ and ‘Fuji’ with melatonin from 56.55 to 67.63 ng/g were the high level group and in the super-high level group, the melatonin levels were from 87.28 to 105.97 ng/g. For example, the melatonin level in the peel of ‘Shinsekai 1’ was 87.28 ng/g, in ‘Jinhong’ it was 102.64 ng/g and in ‘Jincui’ it was 105.97 ng/g, which were 1.25-, 1.46- and 1.55-fold higher than that in the most popular ‘Fuji’ apple (67.63 ng/g), respectively (Figure 2a).
Based on the apple flesh melatonin levels, these cultivars were also classified into four groups (Table 2 in the Supplementary Data) like the peel. In the low level group, the melatonin content in the flesh of nine cultivars (‘Fuji’, ‘Ralls’, ‘Granny Smith’, ‘Orin’, ‘Jiguan’, ‘Baishaguo’, ‘Ben Davis’, ‘Golden Delicious’ and ‘Changfu 2’) ranged from 0.86 ng/g to 19.22 ng/g; in the medium level group, which included seven apple cultivars (‘Jonagold’, ‘Stark Jumbo’, ‘Jincui’, ‘Xiaoshuai’, ‘4-23’, ‘Tompkin’s King’ and ‘Shanfu 2’), melatonin ranged from 25.55 ng/g to 39.0 ng/g; ‘Jinhong’ and ‘Shinsekai 1’ were both the only cultivar in the high and super-high level groups, with 106.54 ng/g and 148.11 ng/g melatonin, respectively (Figure 2b).
2.4. The Melatonin Content Reduced during Juicing Procedure and Storage Time
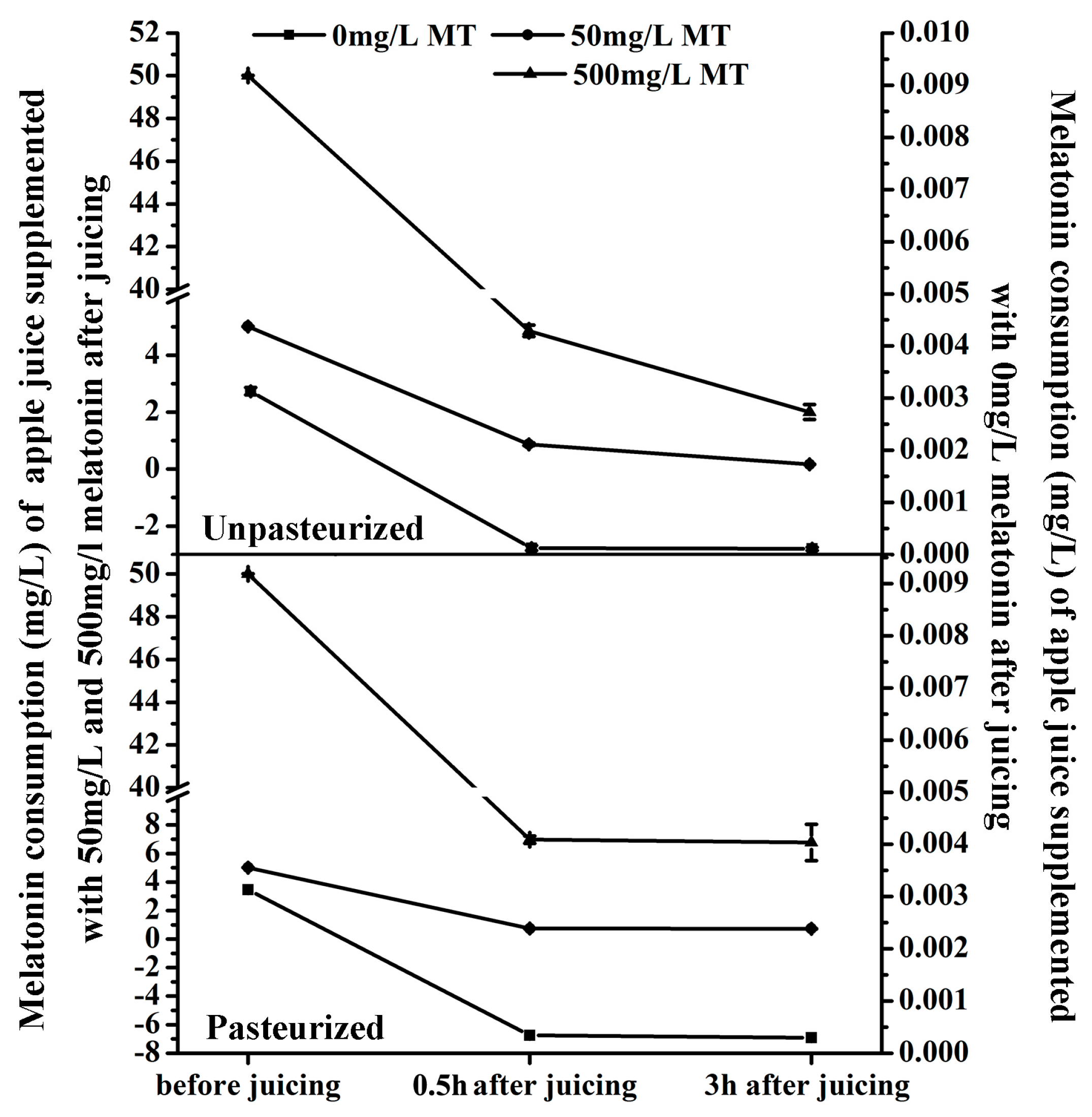
The melatonin content in ‘Fuji’ apple was detected at three time points: just after juicing, just after pasteurization (only for the pasteurized juice, pasteurization was carried out just after juicing) and three hours after juicing (for both the pasteurized juice and unpasteurized juice). The melatonin level was reduced sharply during the juicing process and then declined gradually during storage, whether the juice was pasteurized or not (Figure 3).
For ‘Fuji’ apple, its juice yield (the ratio of final apple juice volume and the weight of apple fruit) is around 70%. The melatonin solution was added into the juice in 7:100 (v/w) before juicing. It means the supplemented melatonin in the apple juice was diluted nearly 10 times. Even without any consumption or loss of the supplemented melatonin, the supplemented melatonin is only 5 mg/L or 50 mg/L left after juicing, respectively.
The average weight of ‘Fuji’ fruit was about 150 g including 3.0 g peel with an average melatonin content of 67.63 ng/g (total melatonin was around 202.89 ng) and 147.0 g flesh with an average melatonin level of 0.857 ng/g (total melatonin was about 125.98 ng) (Figure 1b). Therefore the average total melatonin content of one ‘Fuji’ apple (including peel and flesh) was about 329 ng. The juice yield of an apple fruit was 105 mL and the average melatonin content of apple juice before juicing was about 3.13 ng/mL (0.00313 mg/L).
In this experiment, the average water for each apple fruit was 10 mL, which was used as blank control for the melatonin supplementation. As shown in Figure 3, 0.5 h after juicing, the melatonin content in the unpasteurized juice of the control decreased from 3.13 ng/mL to 0.125 ng/mL, a reduction of 96% (Figure 3a). When 50 mg/L and 500 mg/L melatonin were supplemented before juicing, the melatonin levels also reduced from 5 mg/L and 50 mg/L to 0.870 mg/L (reduction by about 83%) and 4.858 mg/L (reduction by about 90%), respectively. In the pasteurized juice, the decrease in melatonin content was similar to the control (Figure 3b). It decreased from 3.13 ng/mL to 0.344 ng/mL (reduction by about 89%) in the control, and from 5 mg/L and 50 mg/L to 0.739 mg/L (reduction by about 85%) and 6.971 mg/L (reduction by about 86%) in 50 mg/L and 500 mg/L melatonin supplemented juice, respectively. Obviously, juicing resulted in a significant melatonin decrease in apple juice, but pasteurization didn’t make a significant difference compared with the unpasteurized apple juice.
After 3 hours’ storage, in the unpasteurized juice the melatonin levels were reduced from 0.125 ng/mL, 0.870 mg/L and 4.858 mg/L to 0.109 ng/mL, 0.166 mg/L and 2.002 mg/L (reductions by about 13%, 80% and 59%) in the blank control and groups treated with 50 mg/L and 500 mg/L melatonin supplementation, respectively (Figure 3a). However, in the pasteurized juice, melatonin levels were only reduced from 0.344 ng/mL, 0.739 mg/L and 6.971 mg/L to 0.296 ng/mL, 0.726 mg/L and 6.780 mg/L (reductions by about 14%, 2%, and 3%) in control and the two groups supplemented with melatonin, respectively (Figure 3b).
2.5. The Effect of Exogenous Melatonin on the Browning for ‘Fuji’ Apple Juicing
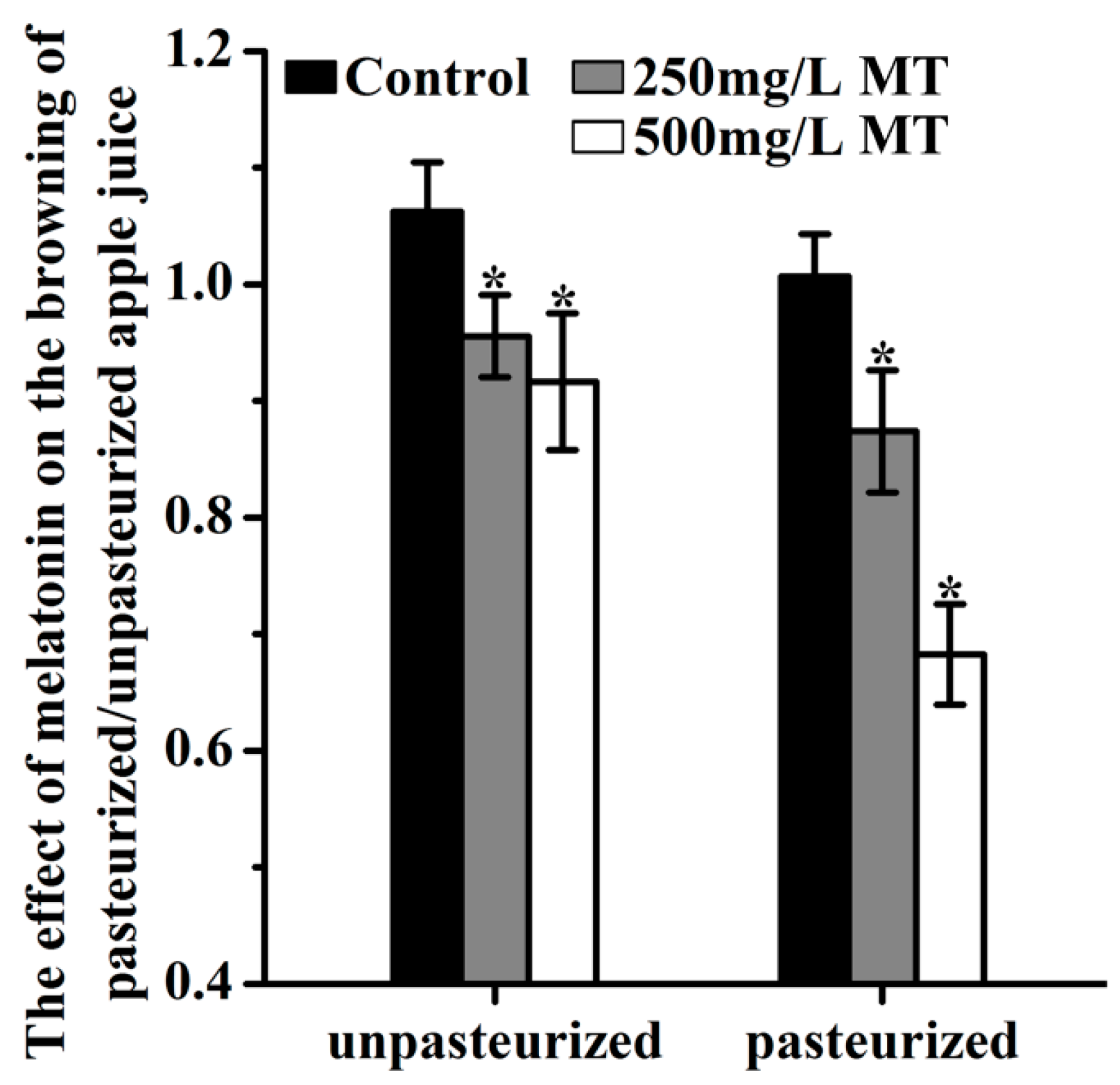
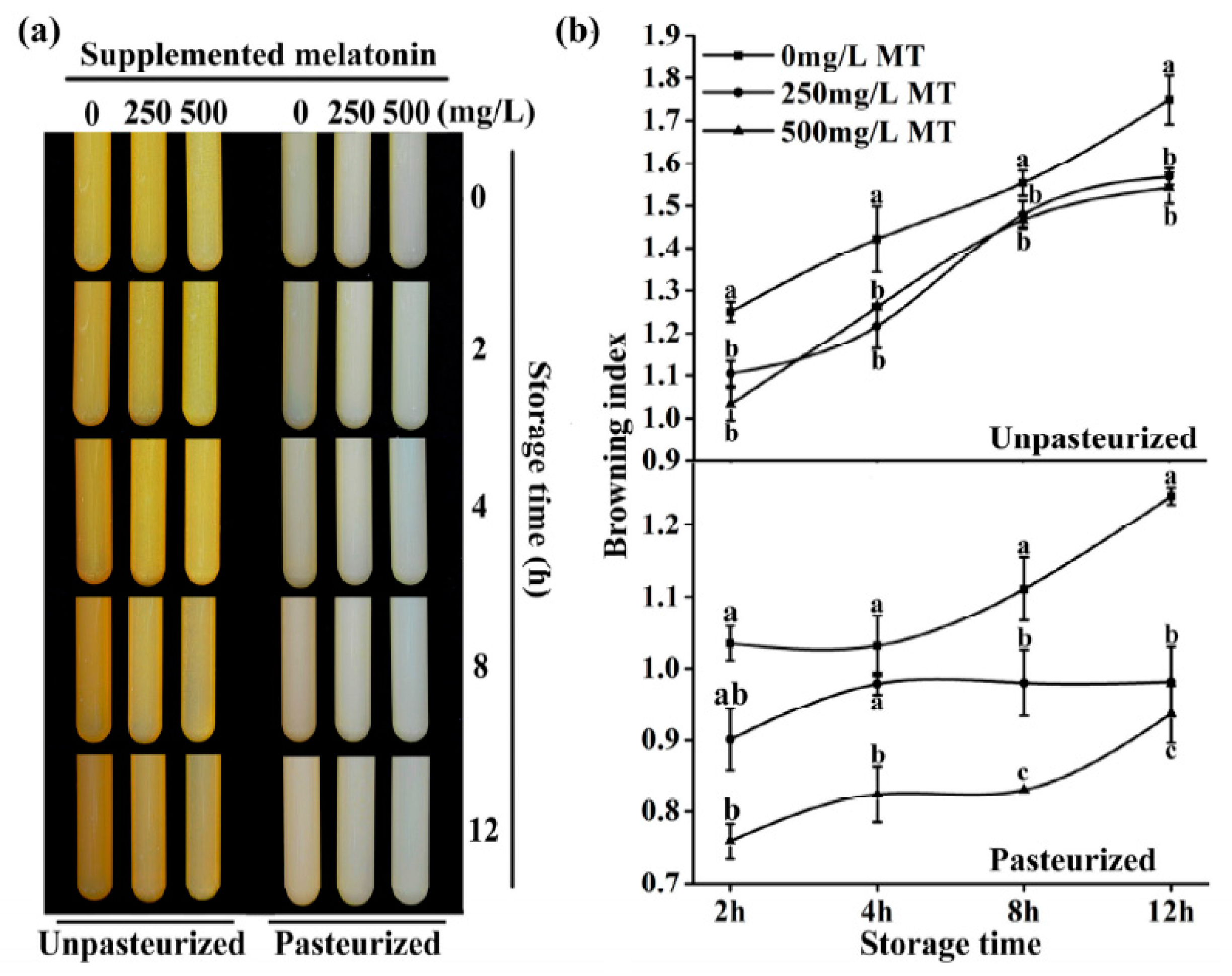
Addition of 250 mg/L and 500 mg/L melatonin before juicing significantly reduced the browning. This was indicated by the changes of browning index and observed whether the juice was pasteurized or not (Figure 4).
The browning index for controls was 1.062 and 1.007 in unpasteurized and pasteurized juices, respectively. In the 250 mg/L added melatonin group, they were 0.956 and 0.874 in the unpasteurized and pasteurized juices respectively. In the 500 mg/L added melatonin group, the indexes were 0.917 and 0.683, respectively, in unpasteurized and pasteurized juices. After 12 hours of storage, the browning was further increased in all groups. Obviously, the browning of unpasteurized juice was more severe than that of pasteurized juice. Both doses of melatonin treatment significantly decreased the juice browning at this time point too (Figure 5)
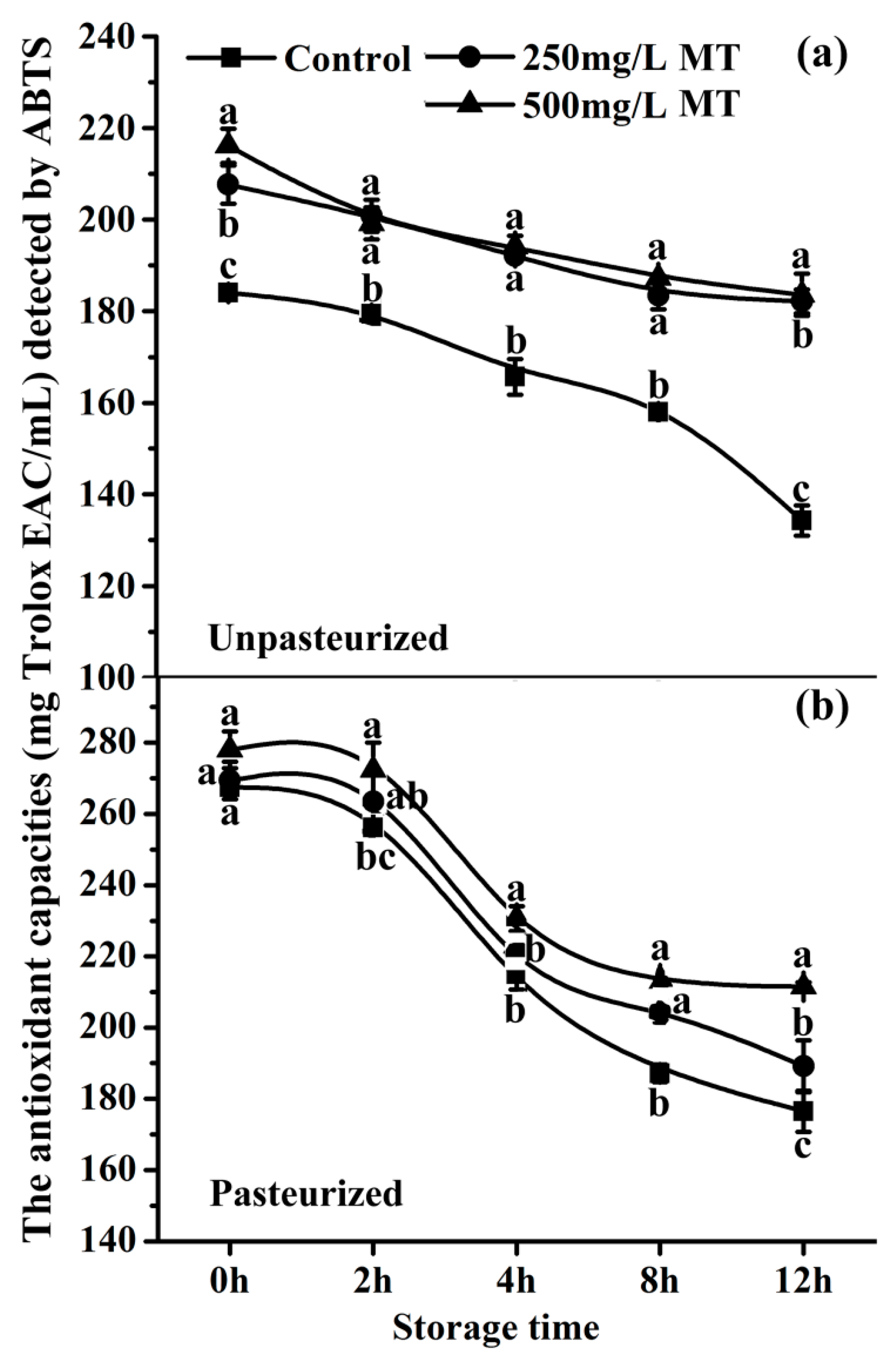
2.7. The Antioxidant Capacity of Exogenous Melatonin in Unpasteurized and Pasteurized ‘Fuji’ Apple Juice during Storage
The ABTS method was applied to detect the antioxidant capacities in unpasteurized and pasteurized ‘Fuji’ apple juice with or without 250 mg/L and 500 mg/L melatonin treatment. During storage, the antioxidant capacity of the unpasteurized control juice gradually decreased during the first 8 hours. Thereafter, this decrease was accelerated from 8 (162.09 mg Trolox EAC/mL) to 12 h (134.29 mg Trolox EAC/mL) (Figure 6a). A different pattern was observed in pasteurized control and 250 and 500 mg/L melatonin treatment juices. The antioxidant capacities of these juices had a sharp decrease from 2 (266.07, 275.59 and 284.52 mg Trolox EAC/mL respectively) to 4 h (208.06, 210.25 and 222.95 mg Trolox EAC/mL respectively), after that, the changes were very slow (Figure 6b). Melatonin supplementation at 500 mg/L significantly increased the antioxidant capacities in both pasteurized and unpasteurized juices, compared to the control or 250 mg/L melatonin groups (Figure 6).
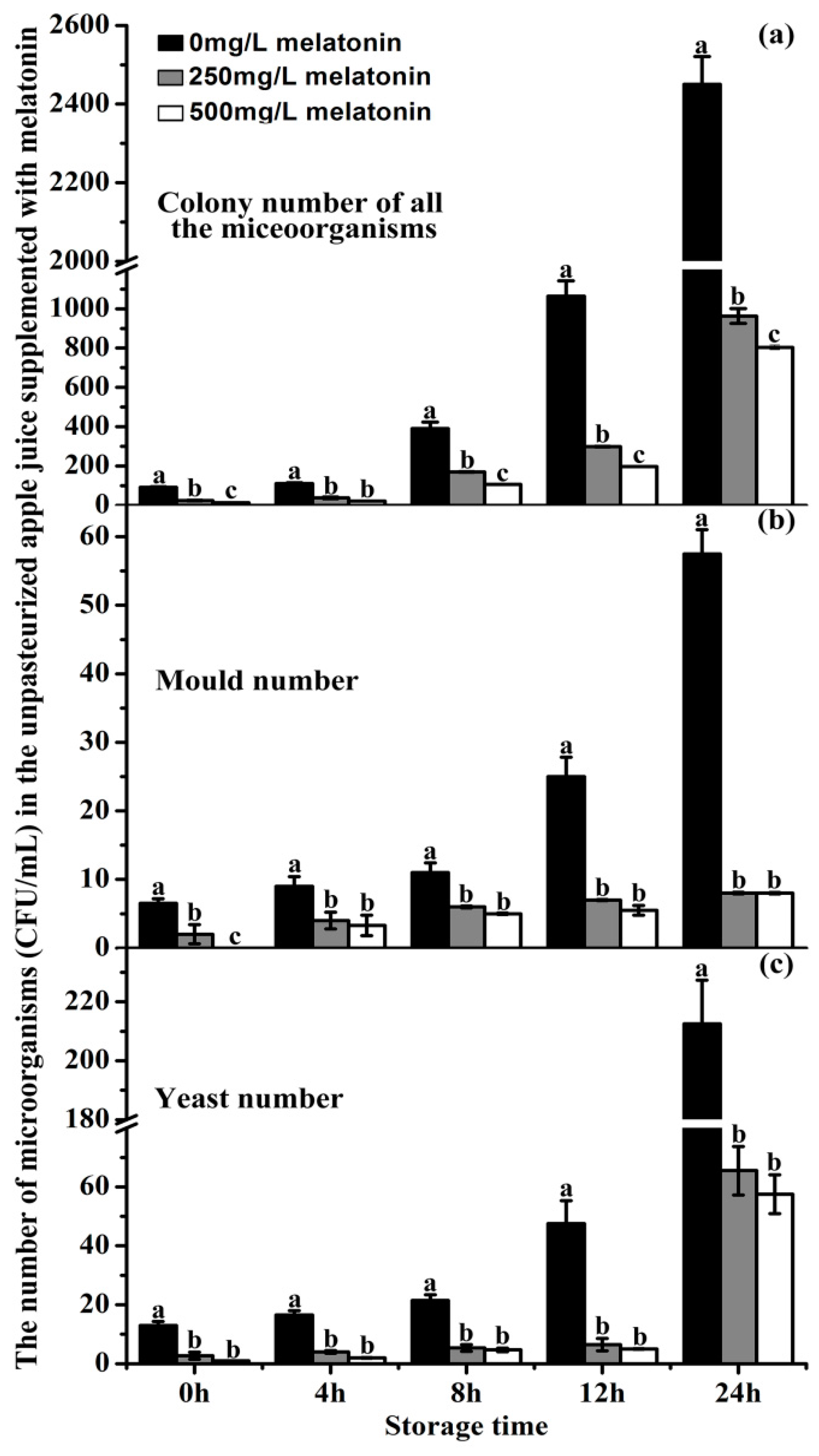
2.8. The Effects of Exogenous Melatonin on the Microorganism Growth in ‘Fuji’ Apple Juice during Storage
The capacity of melatonin to inhibit microorganism growth was tested in unpasteurized ‘Fuji’ apple juice by counting the colony numbers of yeast, moulds and other microorganisms. As expected, the microorganisms proliferated rapidly during the 24 h of juice storage. In the control juice, at 12 h storage, the number of microorganisms had already exceeded its quality guarantee limits. In details, its total colony number of microorganisms at 12 h storage were 1064 CFU/mL which was roughly 12-fold higher than that at 0 h (91 CFU/mL) (Figure 7a). Both 250 and the 500 mg/L melatonin treatments significantly inhibited microorganism growth (including yeasts, moulds and bacteria), in juice. The total microorganisms in 250 and 500 mg/L melatonin-treated juices were 299 CFU/mL and 197 CFU/mL, respectively, which were only 28 and 19% of the control regarding the number of microorganisms.
At 12 h storage, the mould colony number and yeast number in the control juice were roughly 4-fold higher than those at time 0 (Figure 7b,c). In contrast, in both added melatonin groups, the mould colony numbers were reduced to 28% (7 CFU/mL) and 22% (5.5 CFU/mL) compared to the control (25 CFU/mL), respectively (Figure 7b); the yeast numbers were decreased to 14% (6.5 CFU/mL) and 11% (5 CFU/mL) related to the control (47.5 CFU/mL), respectively (Figure 7c). The inhibition of exogenous melatonin on microorganism growth was detected at all time points analyzed. It appeared that the inhibitory effect of 500 mg/L melatonin on microorganism growth was more potent than that of 250 mg/L (Figure 7).
3. Discussion
Melatonin is a popular food supplement with human health benefits [59]. It was first reported to be present in apple fruit in 2013 [19]; therefore, it is expected that apple juice should be rich in melatonin. If so, apple juice could serve as an alternative source of naturally occurring melatonin which may be more welcome by the consumers compared to the synthetic version. In the present study, the melatonin content in apple peel and flesh were tested in variety of cultivars (18 in total) (Figure 2). The apple cultivars rich in melatonin apple could be selected to produce ‘apple juice rich in melatonin’ in the future. Melatonin content in these cultivars exhibited substantial variations. Two cultivars (‘Jinhong’ and ‘Shinsekai 1’) had the highest melatonin content among others (Figure 2). These two apple cultivars will be candidates for producing ‘melatonin-rich juice’ in the future. The content of melatonin as well as the total phenols in ‘Fuji’ apple peel was significantly higher than that in apple flesh and juice (Figure 1b,c). It is well known that apple peel contains higher levels of antioxidant chemicals such as phenols and flavonoids than flesh and therefore has higher antioxidant activity [60,61,62,63]. During the coloration in the final stage of apple development, the high intensity of light during the daytime and the low temperature at night probably contribute to the accumulation of these antioxidants. Melatonin levels in the peel are likely induced by environmental stressors. This is supported by the previous reports that melatonin content increased when fruits are subject to stressors like cold or high light intensity [64,65,66,67,68,69,70,71].
Compared to the melatonin level in peel (67.63 ng/gFW), the melatonin content in Fuji apple juice (0.814 ng/gFW) is comparable to that in flesh (0.857 ng/gFW), (Figure 1b). A similar result was also observed in ‘Granny Smith’ apple juice. It appears that the juice melatonin level was decided by the flesh melatonin. However, the possibility of melatonin losses during the processing of juice could not be ignored. Initially, we thought that only the free melatonin was left in juice and the bound melatonin was lost. In addition, incomplete cell breakage might also result in melatonin being lost. Our results indicated that melatonin addition (250 mg/L and 500 mg/L) before the juicing process also resulted in 80~90% melatonin loss. It is obvious that this melatonin loss during juice processing is due to the reactions of melatonin with oxidants. This was further proved by the protective effects of melatonin on juice browning. When the juice was exposed to the air it rapidly changed color to brown (referred to as browning). The majority of clinical trials on the therapeutic usefulness of melatonin in different fields of medicine have shown very low toxicity of melatonin over a wide range of doses [72], and melatonin is also recommended to be used as a health food supplement in China [73]. In addition, the commercial melatonin tablets contain 1 mg, 3 mg or 5 mg melatonin. Therefore, addition of synthetic melatonin as a healthy supplement to apple juice would be legal.
This browning process is probably via the conversion to quinones of o-diphenolic compounds which is the main oxidative reaction occurring during fruit color change [74] Melatonin addition at the concentrations of 250 and 500 mg/L significantly reduced the browning in unpasteurized apple juice, during juicing and storage, respectively (Figure 4 and Figure 5). This was due to the potent antioxidant capacity of melatonin indicated by the free radical scavenging ABTS analysis (Figure 6a). Likewise, for the pasteurized apple juice, 500 mg/L supplemented melatonin also significantly inhibited the browning, which was also consistent with the antioxidant capacities detected (Figure 6). The effect of 250 mg/L melatonin on inhibiting juice browning was not obvious as in the pasteurized juice. The pasteurization causes Maillard reactions which create more anti-oxidant chemicals and possibly consume less melatonin [51]. Most importantly, a powerful anti-microorganism activity of melatonin was uncovered during juice storage, although melatonin is a molecule frequently reported to reduce the biotic stress and inhibit the growth of microorganisms in plants [75,76,77,78,79,80,81,82,83,84,85], this effect on food storage was not known, especially in the fresh juice with a shelf life of less than 4 hours. Our results showed that melatonin significantly inhibited the total microorganisms, including bacteria, moulds and yeasts during the juice storage during the tested time from 4 to 12 h (Figure 7). According to the national guidelines regarding the number of microorganisms in juice [86,87], fresh apple juice can’t be sold eight hours after juicing at room temperature (25 °C ± 2 °C). However, when melatonin (250 or 500 mg/L) was added, judging from its microorganism number, the apple juice was still safe to be consumed. We observed that melatonin content gradually declined in juice with time. This is consistent with its effect on the juice browning, i.e., melatonin consumption by interaction with oxidants reduces the juice browning. The exact mechanisms of how melatonin exhibits its powerful anti-microorganism effect are currently unknown and they are our future research subject.
In summary, melatonin was detected in the apple juice and may serve as a naturally occurring melatonin supplement in the future. Melatonin not only reduces the apple juice browning by its potent antioxidant capacity, but also inhibits the growth of a wide spectrum of microorganisms including yeasts, moulds and bacteria. These effects of melatonin would preserve the appearance and prolong the shelf life of apple juice.
4. Materials and Methods
4.1. Chemicals and Reagents
Melatonin (≥98%) was purchased from Sigma (St. Louis, MO, USA). Folin-Ciocalteau phenol reagent, gallic acid, ABTS, glucose and chloramphenicol were supplied by Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China). Methanol, ethyl alcohol, sodium carbonate and potassium persulfate were purchased from Beijing Guangda Smirnov Technology Co., Ltd. (Beijing, China). Trolox, tryptone and yeast powder were obtained from Beijing Jingrijindian Science and Technology Ltd. (Beijing, China). Agar was purchased from Beijing AUKS Technology Co., Ltd. (Beijing, China). All reagents were of analytical grade, except the methanol which was of chromatographic grade.
4.2. Growth Conditions
‘Fuji’ apple (Malus pumila Mill.) fruits were collected from 9-year-old ‘Fuji’ apple trees which were grafted onto ‘Rehd’ (Malus robusta Carr.) stocks and planted at the Jinguoshu Science and Technology Center in Beijing, located at 39°96’ N and 116°22’ W, on October 25th in 2016 and 2017. Apple fruits of the other cultivars (‘Baishaguo’, ‘Orin’, ‘Ralls’, ‘Granny Smith’, ‘Changfu 2’, ‘Ben Davis’, ‘Golden Delicious’, ‘Jiguan, Shanfu 2’, ‘4-23’, ‘Xiaoshuai’, ‘Jonagold’, ‘Stark Jumbo’, ‘Tompkin’s King’, ‘Shinsekai 1’, ‘Jinhong’ and ‘Jincui’) were collected from the 5~8-year-old apple trees planted in the Bakou Apple Orchard in Beijing, on September 16th in 2017.
4.3. Preparation of Apple Peel, Flesh and Juice
The apple fruit was peeled and the skin and flesh was prepared after removing the apple core and slicing; the samples were quick-frozen with liquid nitrogen, and then were kept at −80 °C; (shown in Figure 1a). Apple juice was made by juicing for 30–40 s (220 V, 50 Hz, 240 W) in a juicer (Fissler-7100, Ida-oberstein, Germany), and then it was collected into 10 mL centrifuge tubes followed by quick-freezing with liquid nitrogen. Two apples were used for the preparation of each sample of the peel, flesh and juice for the melatonin detection among the different cultivars. For ‘Fuji’ cultivar, total phenols were also detected. Three duplicate experiments were independently conducted in each cultivar.
4.4. The Melatonin Supplementation and Juice Pasteurization
Three apples were weighed and used for ‘Fuji’ apple juicing. To detect the melatonin reduction after juicing and storage, 50 and 500 mg/L melatonin were added in the juice in 7:100 (v/w), respectively. Based on the result of this first melatonin supplementation, in the following experiment to check the juice browning the antioxidant ability of melatonin in juice, before juicing, 250 or 500 mg/L melatonin dissolved into water was added in 7:100 (v/w) [88]. The process of juicing lasted 30–40 s (220 V, 50 Hz, 240 W) in the juicer (Fissler-7100). After juicing, half of the juice of the control and the two samples supplemented with melatonin were immediately pasteurized at 85–90 °C for 20 min. After the juicing, it took a total of 30 min for the treatment before the juice was frozen for melatonin detection. The other unpasteurized half of the juice was also left at room temperature for about half hour as well. After pasteurization, all the unpasteurized/pasteurized apple juice was frozen immediately. Three duplicate experiments were independently conducted with or without pasteurization. In addition, the final concentrations of melatonin in all samples were analyzed after juicing to compare anti-oxidative capacity of different melatonin levels in apple juice.
4.5. Melatonin and Total Phenols Detection of Apple Fruit
4.5.1. Melatonin Detection
A total of 1.5–2.0 g frozen apple peel or flesh, respectively, were ground to a fine powder in liquid nitrogen with a mortar and pestle and suspended in 5 mL methanol and ultrasonicated (at 80 Hz) for 35 min at 45 °C. After centrifugation at 10,000 g at 4 °C for 15 min, the supernatants were collected and dried under nitrogen gas. The extracts were dissolved in 2 mL 5% methanol and transferred to a C18 solid phase extraction (SPE) cartridges (Waters, Milford, MA, USA), which were activated with 2 mL methanol and 2 mL water respectively, for the purification of melatonin. Then the cartridges were washed with 2 mL 5% methanol and melatonin was eluted at a low flow rate with 2 mL 80% methanol, and filtered through 0.2 μm nylon membrane filters (Titan Syringe Filters, Sun Sri, Rockwood, TN, USA). The purified extracts were subjected to HPLC analysis [89,90]. The fruit juice (1 mL) was diluted with 5 mL methanol and then melatonin was detected as the method described above. The melatonin detection of each sample was independently repeated three times.
The recovery rate was determined by adding 100 μL 1 μg/mL standard melatonin (Sigma) solution into the suspension of samples before melatonin extraction using the methods described by Zhao [89]. The melatonin-spiked plant extracts were then analyzed by HPLC.
4.5.2. Total Phenols Detection
The total phenolic content (TPC) was assessed using the Folin-Ciocalteau phenol reagent method [91,92] with a slight modification. A total of 3.0 g frozen apple peel or 5.0 g frozen fruit flesh, were ground to a fine powder in liquid nitrogen with a mortar and pestle and suspended in 20 mL 80% (v/v) methanol for 15 min at room temperature, respectively. Then they were diluted with 30 mL 50% (v/v) methanol and centrifuged for 15 min at 6000 rpm at room temperature. Then 4.8 mL distilled water and 300 μL of Folin-Ciocalteau reagent were added to 300 μL of the extracted sample. After 8 minutes of vortex oscillation at room temperature, 600 μL of sodium carbonate (1 N) were added. The extracts were mixed and kept in the dark at room temperature for 2 h before measuring the absorbance at 725 nm using a UV–visible spectrophotometer (Thermo Scientific Technologies, Madison, WI, USA). A mixture of water and reagents was used as a blank. Gallic acid standard curve (0.02−0.10 mg/mL) was used and the total phenolic content was expressed as milligram gallic acid equivalents per gram fresh weight (mgGAE/gFW) of samples. Each experiment was independently repeated three times.
4.6. Browning Observation and the Browning Index Detection of Apple Juice
4.6.1. Browning Observation
The browning of unpasteurized/pasteurized apple juice was observed in 10 mL glass tubes. Photos were taken at hour 0, 2, 4, 8, 12 after juicing using camera (Nikon D7000, Tokyo, Japan).
4.6.2. Browning Index Detection
The browning index (BI) was measured using the following method [93,94,95]. A total of 10 mL of apple juice was centrifuged for 2 min at 8000 rpm and at room temperature 25 °C ± 2 °C to remove coarse particles, and then 5 mL of absolute ethyl alcohol was added to 5mL of supernatant before the absorbance of the obtained supernatant measured at 420 nm using a UV–visible spectrophotometer (Thermo Scientific Technologies). The browning index of pasteurized and unpasteurized apple juice was both determined at hour 0, hour 2, 4, 8 and 12 after juicing. Each measurement was repeated three times independently.
4.7. Detection of Antioxidant Capacity of Apple Juice by the ABTS Assay
The free radical scavenging activity of the apple juice with different concentrations of melatonin was measured using ABTS reagent, which was prepared by mixing 5 mL 7.4 mM ABTS solution and 5 mL 2.6 mM potassium persulfate solution together to make them react for 16 h at room temperature 25 °C ± 2 °C; in the dark [96]. Then a total of 20 µL apple juice was added to a 1950 µL ABTS reagent to react for 6mins. The absorbance was determined at 734 nm using a UV–visible spectrophotometer (Thermo Scientific Technologies). The free radical scavenging activity was compared to a standard curve of prepared Trolox solutions. Each experiment was independently repeated three times.
4.8. The inhibition of Melatonin on Microorganisms
Total colonies were counted in on plate count agar (PCA) [86,97], following inoculation with 100 μL of apple juice supplemented with 0, 250 and 500 mg/L melatonin or its serial dilutions in physiological saline incubated at 36 °C ± 1 °C for 48 h ± 2 h. Mould and yeast counts were calculated in potato dextrose agar (PDA) [88], following inoculation with 100 μL of apple juice with different concentrations of melatonin or its serial dilutions in sterile water incubated at 28 °C ± 1 °C for 5 days. The results were expressed as colony forming units per milliliter of juice (CFU/mL). Three duplicates were independently conducted.
4.9. Statistical Analysis
The data were subjected to ANOVA followed by Fisher’s least significant difference (LSD) or Student’s t-test analysis. Statistically significant differences were indicated at p < 0.05. Statistical computations were carried out using the SPSS 20.0 software (IBM, Armonk, NY, USA).
Supplementary Materials
Supplementary Materials are available online.
Acknowledgments
This research was supported by National Natural Science Fund (No. 3050216135).
Author Contributions
Designed the studies: Jin Kong. Undertook the experimental work: Haixia Zhang, Xuan Liu, Ting Chen, Yazhen Ji. Contributed to figures and manuscript preparation: Haixia Zhang, Kun Shi, Lin Wang, Xiaodong Zheng and Jin Kong. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no competing financial interests.
References
- Food and Agriculture Organization of the United Nations. Available online: http://faostat.fao.org/default.aspx?alias=faostatclassic (accessed on 23 February 2018).
- US Apple Statistics, US Apple Association. Available online: http://www.usapple.org/industry/applestats/index.cfm (accessed on 23 February 2018).
- Ibanez, A.M.; Dandekar, A.M. Apple. In Transgenic Crops V; Pua, E.C., Davey, M.R., Eds.; Springer: Berlin and Heidelberg, Germany, 2007; p. 60. [Google Scholar]
- Hecke, K.; Herbinger, K.; Veberič, R.; Trobec, M.; Toplak, H.; Štampar, F.; Keppel, H.; Grill, D. Sugar-, acid- and phenol contents in apple cultivars from organic and integrated fruit cultivation. Eur. J. Clin. Nutr. 2006, 60, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Biedrzycka, E.; Amarowicz, R. Diet and health: apple polyphenols as antioxidants. Food Rev. Int. 2008, 24, 235–251. [Google Scholar] [CrossRef]
- Petkovsek, M.M.; Stampar, F.; Veberic, F. Changes in the inner quality parameters of apple fruit from technological to edible maturity. Acta Agric. Slov. 2009, 93, 17–29. [Google Scholar]
- Radunić, M.; Klepo, T.; Strikić, F.; Lukić, D.; Maretić, M. Karakteristike sorata jabuka (Malus x domestica Borkh.) uzgajanih na području Žrnovnice. Pomologia Croat. 2011, 17, 11–17. [Google Scholar]
- Balík, J.; Rop, O.; Mlček, J.; Híc, P.; Horák, M.; Řezníček, V. Assessment of nutritional parameters of native apple cultivars as new gene sources. Acta Univ. Agric. Silvic. Mendelianae Brun. 2012, 60, 27–38. [Google Scholar] [CrossRef]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Marinoni, D.T.; Cerutti, A.K.; Canterino, S.; Bounous, G. Application of sensory, nutraceutical and genetic techniques to create a quality profile of ancient apple cultivars. J. Food Qual. 2012, 35, 169–181. [Google Scholar] [CrossRef]
- Candrawinata, V.I.; Golding, J.B.; Roach, P.D.; Stathopoulos, C.E. From apple to juice—the fate of polyphenolic compounds. Food Rev. Int. 2013, 29, 276–293. [Google Scholar] [CrossRef]
- Biegańska-Marecik, R.; Radziejewska-Kubzdela, E.; Marecik, R. Characterization of phenolics, glucosinolates and antioxidant activity of beverages based on apple juice with addition of frozen and freeze-dried curly kale leaves (Brassica oleracea L. var. acephala L.). Food Chem. 2017, 230, 271–280. [Google Scholar]
- Hyson, D.A. A comprehensive review of apples and apple components and their relationship to human health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Hattori, A.; Migitaka, H.; Iigo, M.; Itoh, M.; Yamamoto, K.; Ohtani-Kaneko, R.; HARA, M.; Suzuki, T.; Reiter, R.J. Identification of melatonin in plants and its effects on plasma melatonin levels and binding to melatonin receptors in vertebrates. Biochem. Mol. Biol. Int. 1995, 35, 627–634. [Google Scholar] [PubMed]
- Afzalia, D.; Azadmehra, F.; Torkzadeha, M. Vortex-assisted dispersive liquid–liquid microextraction trace amounts of melatonin prior to HPLC determination in fruit juice samples. Sep. Sci. Technol. 2016, 51, 1509–1514. [Google Scholar] [CrossRef]
- Howatson, G.; Bell, P.G.; Tallent, J.; Middleton, B.; McHugh, M.P.; Ellis, J. Effect of tart cherry juice (Prunus cerasus) on melatonin levels and enhanced sleep quality. Eur. J. Nutr. 2012, 51, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pachón, M.S.; Medina, S.; Herrero-Martín, G.; Cerrillo, I.; Berna, G.; Escudero-López, B.; Ferreres, F.; Martín, F.; García-Parrilla, M.C.; Gil-Izquierdo, A. Alcoholic fermentation induces melatonin synthesis in orange juice. J. Pineal Res. 2014, 56, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, Y.; Herrera, T.; Benítez, V.; Arribas, S.M.; López de Pablo, A.L.; Esteban, R.M.; Martín-Cabrejas, M.A. Estimation of scavenging capacity of melatonin and other antioxidants: Contribution and evaluation in germinated seeds. Food Chem. 2015, 170, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.F.; Shi, T.C.; Song, S.; Zhang, Z.W.; Fang, Y.L. Melatonin in grapes and grape-related foodstuffs: A review. Food Chem. 2017, 231, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Wang, L.; Tan, D.X.; Zhao, Y.; Zheng, X.D.; Chen, H.; Li, Q.T.; Zuo, B.X.; Kong, J. Identification of genes for melatonin synthetic enzymes in ‘Red Fuji’ apple (Malus domestica Borkh.cv. Red) and their expression and melatonin production during fruit development. J. Pineal Res. 2013, 55, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S.; Daglia, M.; Sureda, A.; Selamoglu, Z.; Gulhan, M.F.; Nabavi, S.M. Melatonin and Respiratory Diseases: A Review. Curr. Top. Med. Chem. 2017, 17, 467–488. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, Z.; Zheng, H.Y.; Ho, J.; Chan, M.T.V.; Wu, W.K.K. Protective roles of melatonin in central nervous system diseases by regulation of neural stem cells. Cell Prolif. 2017, 50, e12323. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; RosalesCorral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2012, 63, 577–597. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Zubero, E.; López-Pingarrón, L.; Alatorre-Jiménez, M.A.; Ochoa-Moneo, P.; Buisac-Ramón, C.; Rivas-Jiménez, M.; Castán-Ruiz, S.; Antoñanzas-Lombarte, Á.; Tan, D.X.; García, J.J.; et al. Melatonin’s role as a co-adjuvant treatment in colonic diseases: A review. Life Sci. 2017, 170, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Asghari, M.H.; Abdollahi, M.; De Oliveira, M.R.; Nabavi, S.M. A review of the protective role of melatonin during phosphineinduced cardiotoxicity: Focus on mitochondrial dysfunction, oxidative stress and apoptosis. J. Pharm. Pharmacol. 2017, 69, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sci. 2017, 173, 94–106. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, G.; Marseglia, L.; Manti, S.; Colavita, L.; Cuppari, C.; Impellizzeri, P.; Arena, S.; Arrigo, T.; Salpietro, C.; Gitto, E. Atopy and autoimmune thyroid diseases: Melatonin can be useful? Ital. J. Pediatr. 2016, 42, 95. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Hsieh, M.J.; Yang, W.E.; Chung, W.H.; Reiter, R.J.; Yang, S.F. Cancer metastasis: Mechanisms of inhibition by melatonin. J. Pineal Res. 2017, 62, e12370. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N. Developmental Programming of Adult Disease: Reprogramming by Melatonin? Int. J. Mol. Sci. 2017, 18, 426. [Google Scholar] [CrossRef] [PubMed]
- Ramos, E.; Patiño, P.; Reiter, R.J.; Gil-Martín, E.; Marco-Contelles, J.; Parada, E.; De Los Rios, E.; Romero, A.; Egea, J. Ischemic brain injury: New insights on the protective role of melatonin. Free Radic. Biol. Med. 2017, 104, 32–53. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Ojike, N.I.; Akinseye, O.A.; Kendzerska, T.; Buttoo, K.; Dhandapany, P.S.; Brown, G.M.; Cardinali, D.P. Melatonin and Human Cardiovascular Disease. Pharmacol. Ther. 2017, 22, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and the pathologies of weakened or dysregulated circadian oscillators. J. Pineal Res. 2017, 62, e12377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, S.; Li, Y.; Liu, Y. Melatonin as a promising agent of regulating stem cell biology and its application in disease therapy. Pharmacol. Res. 2017, 117, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Watson, N.; Diamandis, T.; Gonzales-Portillo, C.; Reyes, S.; Borlongan, C.Y. Melatonin as an Antioxidant for Stroke Neuroprotection. Cell Transplant. 2016, 25, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Yin, T.C.; Sung, P.H.; Chiang, J.Y.; Sun, C.K.; Yip, H.K. Melatonin enhances survival and preserves functional integrity of stem cells: A review. J. Pineal Res. 2017, 62, e12372. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Zubero, E.; García-Gil, F.A.; López-Pingarrón, L.; Alatorre-Jiménez, M.A.; Ramírez, J.M.; Tan, D.X.; García, J.J.; Reiter, R.J. Melatonin role preventing steatohepatitis and improving liver transplantation results. Cell Mol. Life Sci. 2016, 73, 2911–2927. [Google Scholar] [CrossRef] [PubMed]
- Nooshinfar, E.; Safaroghli-Azar, A.; Bashash, D.; Akbari, M.E. Melatonin, an inhibitory agent in breast cancer. Breast Cancer 2017, 24, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.W.; Zhang, X.; Huang, W.J. Pain control by melatonin: Physiological and pharmacological effects. Exp. Ther. Med. 2016, 12, 1963–1968. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Ma, Z.Q.; Di, S.; Jiang, S.; Li, Y.; Fan, C.X.; Yang, Y.; Wang, D.J. Snapshot: Implications for melatonin in endoplasmic reticulum homeostasis. Br. J. Pharmacol. 2016, 173, 3431–3442. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Cordaro, M.; Esposito, E.; Cuzzocrea, S. The antioxidative property of melatonin against brain ischemia. Expert Rev. Neurother. 2016, 16, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.A.; Garde, A.H.; Kristiansen, J.; Nabe-Nielsen, K.; Hansen, A.M. The effect of the number of consecutive night shifts on diurnal rhythms in cortisol, melatonin and heart rate variability (HRV): A systematic review of field studies. Int. Arch. Occup. Environ. Health 2016, 89, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Campa, C.; Menéndez-Menéndez, J.; Alonso-González, C.; González, A.; Álvarez-García, V.; Cos, S. What is known about melatonin, chemotherapy and altered gene expression in breast cancer. Oncol. Lett. 2017, 13, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchster, L.C.; Qi, W.B.; Karbownik, M.; Calvo, J.R. Significance of melatonin in antioxidative defense system: Reactions and products. Biol. Signals Recept. 2000, 9, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchster, L.C.; Terron, M.P.; Flores, L.J.; Reiter, R.J. One molecule, many derivatives: a never-ending interaction of melatonin with reactive oxygen and nitrogen species? J. Pineal Res. 2007, 42, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchster, L.C.; Di Mascio, P.; Martinez, G.R.; Prado, F.M.; Reiter, R.J. Novel rhythms of N1-acetyl-N2-formyl-5-methoxykynuramine and its precursor melatonin in water hyacinth:importance for phytoremediation. Faseb. J. 2007, 21, 1724–1729. [Google Scholar] [CrossRef] [PubMed]
- Guenther, A.L.; Schmidt, S.I.; Laatsch, H.; Fotso, S.; Ness, H.; Ressmeyer, A.R.; Poeggeler, B.; Hardeland, R. Reactions of the melatonin metabolite AMK (N1-acetyl-5-methoxykynuramine) with reactive nitrogen species: formation of novel compounds, 3-acetamidomethyl-6-methoxycinnolinone and 3-nitro-AMK. J. Pineal Res. 2005, 39, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: a physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Blask, D.E.; Dauchy, R.T.; Sauer, L.A.; Krause, J.A. Melatonin uptake and growth prevention in rat hepatoma 7288CTC in response to dietary melatonin: Melatonin receptor-mediated inhibition of tumor linoleic acid metabolism to the growth signaling molecule 13-hydroxyoctadecadienoic acid and the potential role of phytomelatonin. Carcinogenesis 2004, 25, 951–960. [Google Scholar] [PubMed]
- Billaud, C.; Brun-Mérimée, S.; Louarme, L.; Nicolas, J. Effect of glutathione and Maillard reaction products prepared from glucose or fructose with glutathione on polyphenoloxidase from apple—I: Enzymatic browning and enzyme activity inhibition. Food Chem. 2004, 84, 223–233. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Bioche. 1976, 72, 248–257. [Google Scholar] [CrossRef]
- Burdurlu, H.S.; Karadeniz, F. Effect of storage on nonenzymatic browning of apple juice concentrates. Food Chem. 2003, 80, 91–97. [Google Scholar] [CrossRef]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Hellens, R.P. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, C.Y.; Park, I. Prevention of enzymatic browning of pear by onion extract. Food Chem. 2005, 89, 181–184. [Google Scholar] [CrossRef]
- Sudha, G.; Priya, M.S.; Shree, R.I.; Vadivukkarasi, S. In vitro free radical scavenging activity of raw pepino fruit (Solanum muricatum aiton). Int. J. Curr. Pharm. Res. 2011, 3, 137–140. [Google Scholar]
- Lu, S.M.; Luo, Y.G.; Turner, E.; Feng, H. Efficacy of sodium chlorite as an inhibitor of enzymatic browning in apple slices. Food Chem. 2007, 104, 824–829. [Google Scholar] [CrossRef]
- López-Nicolás, J.M.; Núñez-Delicado, E.; Sánchez-Ferrer, Á.; García-Carmona, F. Kinetic model of apple juice enzymatic browning in the presence of cyclodextrins: The use of maltosyl-β-cyclodextrin as secondary antioxidant. Food Chem. 2007, 101, 1164–1171. [Google Scholar] [CrossRef]
- Luo, Y.; Lu, S.; Zhou, B.; Feng, H. Dual effectiveness of sodium chlorite for enzymatic browning inhibition and microbial inactivation on fresh-cut apples. LWT-Food Sci. Technol. 2011, 44, 1621–1625. [Google Scholar] [CrossRef]
- Zhao, D.Y.; Yuan, J.C.; Xu, K.; Cheng, C.G.; Yan, S. Analysis of apple production and marketing at home and abroad in the past 10 years. Chinese Tree 2016, 3, 87–93. [Google Scholar]
- Cerezo, A.B.; Leal, A.; Alvarez-Fernandez, M.A.; Hornedo-Ortega, R.; Troncoso, A.M.; Garcia-Parrilla, M.C. Quality control and determination of melatonin in food supplements. J. Food Compos. Anal. 2016, 45, 80–86. [Google Scholar] [CrossRef]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.; Kim, Y.J.; Kim, D.O.; Lee, H.J.; Lee, C.Y. Major phenolics in apple and their contribution to the total antioxidant capacity. J. Agric. Food Chem. 2003, 51, 6516–6520. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant activity of apple peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Petkovšek, M.M.; Štampar, F.; Veberič, R. Parameters of inner quality of the scab resistant and susceptible apple in organic and integrated production. Sci. Hortic. 2007, 114, 37–44. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Growth conditions determine different melatonin levels in Lupinus albus L. J. Pineal Res. 2013, 55, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.R.; Fan, J.B.; Xie, Y.; Amombo, E.; Liu, A.; Gitau, M.M.; Khaldun, A.B.M.; Chen, L.; Fu, J.M. Comparative photosynthetic and metabolic analyses reveal mechanism of improved cold stress tolerance in bermudagrass by exogenous melatonin. Plant Physio. Biochem. 2016, 100, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.H.; Huang, B.; Ding, C.B.; Zhang, Z.W.; Chen, Y.E.; Hu, C.; Zhou, L.J.; Huang, Y.; Liao, J.Q.; Yuan, S.; et al. Effects of melatonin on anti-oxidative systems and photosystem II in cold-stressed rice seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Yao, J.F.; Wang, X.N.; You, C.X.; Wang, X.F.; Hao, Y.J. MdHY5 positively regulates cold tolerance via CBF-dependent and CBF independent pathways in apple. J. Plant Physiol. 2017, 218, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, S.V.; Shukla, R.M.; Sherif, M.S.; Murch, J.S.; Saxena, K.P. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Turk, H.; Erdal, S.; Genisel, M.; Atici, O.; Demir, Y.; Yanmis, D. The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul. 2014, 74, 139–152. [Google Scholar] [CrossRef]
- Zheng, X.D.; Tan, D.X.; Allan, C.A.; Zuo, B.X.; Zhao, Y.; Reiter, J.R.; Wang, L.; Wang, Z.; Guo, Y.; Zhou, J.Z.; et al. Chloroplastic biosynthesis of melatonin and its involvement in protection of plants from salt stress. Sci. Rep. 2017, 7, 41236. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.D.; Zhou, J.Z.; Tan, D.X.; Wang, N.; Wang, L.; Shan, D.Q.; Kong, J. Melatonin improves waterlogging tolerance of Malus baccata (Linn.) Borkh. seedlings by maintaining aerobic respiration, photosynthesis and ROS migration. Front. Plant Sci. 2017, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Tan, D.X.; Reiter, R.J. Clinical uses of melatonin: Evaluation of human trials. Curr Med. Chem. 2010, 17, 2070–2095. [Google Scholar] [CrossRef] [PubMed]
- SFDA. State Food and Drug Administration. Available online: http://www.sfda.gov.cn/WS01/CL0055/10396_7.html China (accessed on 20 May 2005).
- Martínez, M.V.; Whitaker, J.R. The biochemistry and control of enzymatic browning. Trends Food Sci. Tech. 1995, 6, 195–200. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Zhou, Z.; Cruz, M.H.; Fuentes-Broto, L.; Galano, A. Phytomelatonin: Assisting plants to survive and thrive. Molecules 2015, 20, 7396–7437. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Zheng, X.; Kong, J.; Manchester, L.C.; Hardeland, R.; Kim, S.J.; Xu, X.; Reiter, R.J. Fundamental issues related to the origin of melatonin and melatonin isomers during evolution: relation to their biological functions. Int. J. Mol. Sci. 2014, 15, 15858–15890. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, Y.; Tan, D.X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin induces nitric oxide and the potential mechanisms relate to innate immunity against bacterial pathogen infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Qian, Y.; Tan, D.X.; Reiter, R.J.; He, C. Melatonin induces the transcripts of CBF/DREBs and their involvement in abiotic and biotic stresses in Arabidopsis. J. Pineal Res. 2015, 59, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Wiid, L.; Helden, E.H.V.; Hon, D.; Lombard, C.; Helden, A.P.V. Potentiation of Isoniazid Activity against Mycobacterium tuberculosis by Melatonin. Antimicrob. Agents Chemother. 1999, 4, 975–977. [Google Scholar]
- Atroshia, F.; Rizzob, A.; Westermarck, A.; Ali-vehmas, T. Effects of tamoxifen, melatonin, coenzyme Q, and 10 L-carnitine supplementation on bacterial growth in the presence of mycotoxins. Pharmacol. Res. 1998, 38, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Tekbas, O.F.; Ogur, R.; Korkmaz, A.; Kilic, A.; Reiter, R.J. Melatonin as an antibiotic: new insights into the actions of this ubiquitous molecule. J. Pineal Res. 2008, 44, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Sumaira; Khan, T.; Abbasi, B.H.; Afridi, M.S.; Tanveer, F.; Ullah, I.; Bashira, S.; Hanod, C. Melatonin-enhanced biosynthesis of antimicrobial AgNPs by improving the phytochemical reducing potential of a callus culture of Ocimum basilicum L. var. thyrsiflora. Rsc. Adv. 2017, 7, 38699. [Google Scholar] [CrossRef]
- Romić, M.D.; Klarić, M.Š.; Lovrić, J.; Pepić, I.; Čižmek, B.C.; Grčić, J.F.; Hafner, A. Melatonin-loaded chitosan/Pluronic® F127 microspheres as in situ forming hydrogel: An innovative antimicrobial wound dressing. Eur. J. Pharm. Biopharm. 2016, 107, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Deng, C.; Ma, Z.Q.; Wang, D.J.; Fan, C.X.; Li, T.; Di, S.Y.; Gong, B.; Reiter, R.J.; Yang, Y. Utilizing melatonin to combat bacterial infections and septic injury. Br. J. Pharmacol. 2017, 174, 754–768. [Google Scholar] [CrossRef] [PubMed]
- GB 4789.2-2010. National food safety standard. Food microbiological examination: Aerobic plate count. Ministry of Health: Beijing, China, 2010.
- GB 4789.15-2010. National food safety standard. Food microbiological examination: Enumeriation of moulds and yeasts. Ministry of Health: Beijing, China, 2010.
- Beveridge, T. Haze and cloud in apple juice. Crit. Rev. Food Sci. Nutr. 1997, 37, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tan, D.X.; Lei, Q.; Chen, H.; Wang, L.; Li, Q.T.; Gao, Y.A.; Kong, J. Melatonin and its potential biological functions in the fruits of sweet cherry. J. Pineal Res. 2012, 55, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Y.; Reiter, R.J.; He, C.J.; Liu, G.S.; Lei, Q.; Zuo, B.X.; Zheng, X.D.; Li, Q.T.; Kong, J. Changes in melatonin levels in transgenic ‘Micro-Tom’ tomato overexpressing ovine AANAT and ovine HIOMT genes. J. Pineal Res. 2014, 56, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colourimetry of total phenolics with phosphomolybdic-phospholungistic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Valentina, U.; Jerneja, F.; Franci, Š. Sugars, organic acids, phenolic composition and antioxidant activity of sweet cherry (Prunus avium L.). Food Chem. 2008, 107, 185–192. [Google Scholar]
- Meydav, S.; Saguy, I.; Kopelman, I.J. Browning determination in citrus products. J. Agric. Food Chem. 1977, 25, 602–604. [Google Scholar] [CrossRef]
- Cruz-Cansino, N.D.; Ramirez-Moreno, E.; Leon-Rivera, J.E.; Delgado-Olivares, L.; Alanis-Garcia, E.; Ariza-Ortega, J.A.; Manriquez-Torres, J.D.; Jaramillo-Bustos, D.P. Shelf life, physicochemical, microbiological and antioxidant properties of purple cactus pear (Opuntia ficus indica) juice after thermoultrasound treatment. Ultrason. Sonochem. 2015, 27, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.H.; Kim, Y.M.; Nam, S.H.; Yim, S.H.; Eun, J.B. Enzymatic browning inhibition and antioxidant activity of pear juice from a new cultivar of asian pear (Pyrus pyrifolia Nakai cv. Sinhwa) with different concentrations of ascorbic acid. Food Sci. Biotechnol. 2016, 25, 153–158. [Google Scholar] [CrossRef]
- Lee, B.; Seo, J.D.; Rhee, J.K.; Kim, C.Y. Heated apple juice supplemented with onion has greatly improved nutritional quality and browning index. Food Chem. 2016, 201, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Valero, M.; Recrosio, N.; Saura, D.; Munoz, N.; Marti, N.; Lizama, V. Effects of ultrasonic treatments in orange juice processing. J. Food Eng. 2007, 80, 509–516. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |
Figure 1.
Melatonin and total phenol content in apple. (a) The ‘Fuji’ apple peel and flesh samples. (b) Melatonin content in peel, flesh, and juice of the ‘Fuji’ and ’Granny Smith’ apple. (c) The total phenol content in the peel, flesh, and juice of ‘Fuji’ apple. (d) The effect of pasteurization on the content of melatonin in ‘Fuji’ apple juice. Data are means ± SD of three independent studies, and three duplicates for each analysis. * indicates significant difference among treatments based on LSD multiple comparison (p < 0.05).
Figure 1.
Melatonin and total phenol content in apple. (a) The ‘Fuji’ apple peel and flesh samples. (b) Melatonin content in peel, flesh, and juice of the ‘Fuji’ and ’Granny Smith’ apple. (c) The total phenol content in the peel, flesh, and juice of ‘Fuji’ apple. (d) The effect of pasteurization on the content of melatonin in ‘Fuji’ apple juice. Data are means ± SD of three independent studies, and three duplicates for each analysis. * indicates significant difference among treatments based on LSD multiple comparison (p < 0.05).


Figure 2.
The melatonin levels in the peel and flesh of different apple cultivars. (a) The melatonin content in the peel of different cultivars. (b) The melatonin content in the flesh of different cultivars. Data are means ± SD of three independent studies, and three duplicates for each analysis.
Figure 2.
The melatonin levels in the peel and flesh of different apple cultivars. (a) The melatonin content in the peel of different cultivars. (b) The melatonin content in the flesh of different cultivars. Data are means ± SD of three independent studies, and three duplicates for each analysis.

Figure 3.
Effects of pasteurization on melatonin levels in apple juice. Melatonin content of unpasteurized and pasteurized ‘Fuji’ apple juice supplemented with 50 or and 500 mg/L MT (melatonin) respectively. Data are means ± SD of three independent studies, and three duplicates for each analysis.
Figure 3.
Effects of pasteurization on melatonin levels in apple juice. Melatonin content of unpasteurized and pasteurized ‘Fuji’ apple juice supplemented with 50 or and 500 mg/L MT (melatonin) respectively. Data are means ± SD of three independent studies, and three duplicates for each analysis.
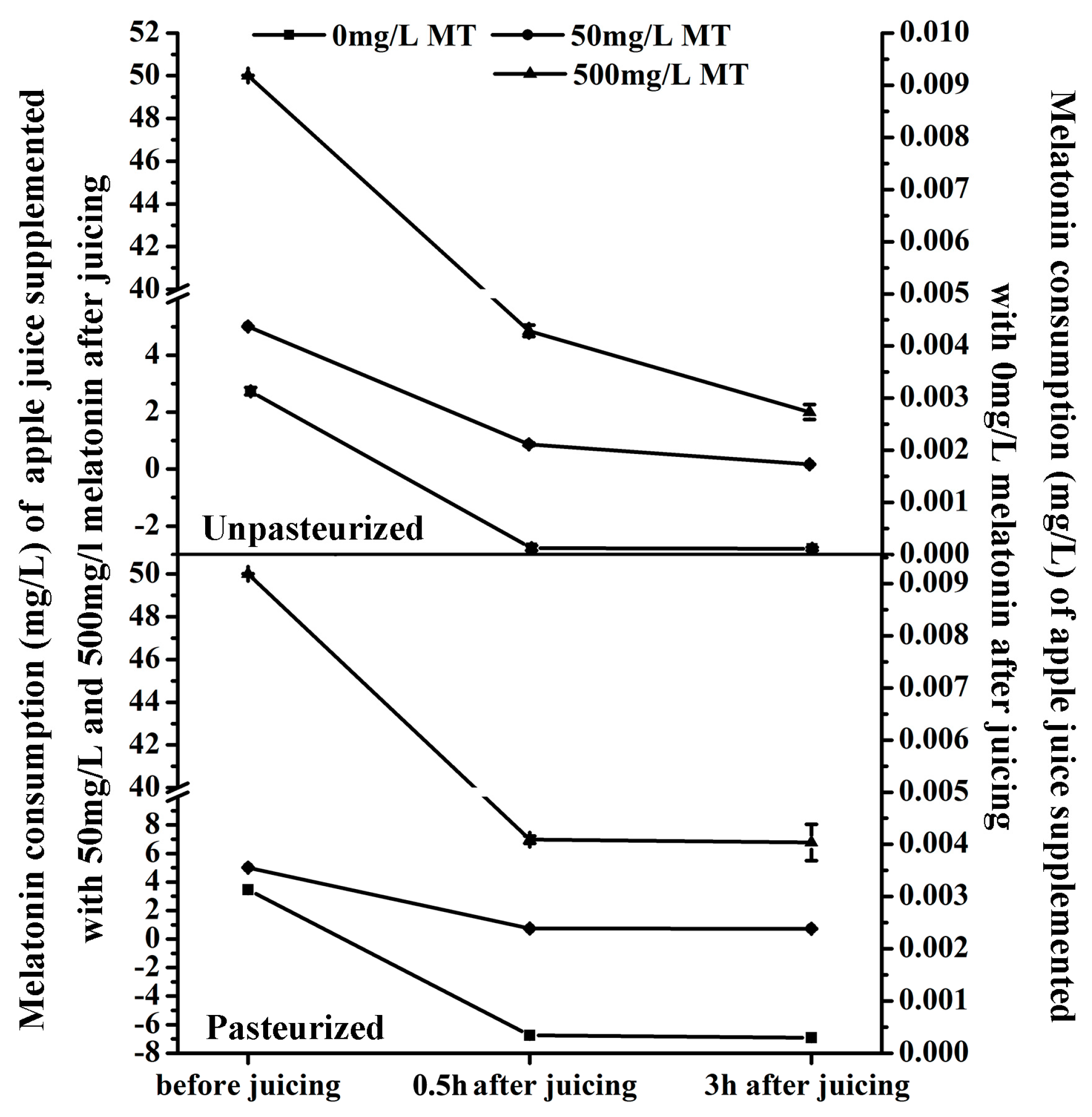
Figure 4.
The effect of melatonin on the browning of unpasteurized or pasteurized ‘Fuji’ apple juice. MT was for melatonin. Data are means ± SD of three independent studies, and three duplicates for each sample. * indicates significant differences among treatments, based on LSD multiple comparison (p < 0.05).
Figure 4.
The effect of melatonin on the browning of unpasteurized or pasteurized ‘Fuji’ apple juice. MT was for melatonin. Data are means ± SD of three independent studies, and three duplicates for each sample. * indicates significant differences among treatments, based on LSD multiple comparison (p < 0.05).
Figure 5.
The effect of melatonin on the browning of ‘Fuji’ apple juice at different storage time. (a) Photo of the ‘Fuji’ apple juice supplemented with 250 or 500 mg/L MT (melatonin) during storage with or without pasteurization. (b) The browning index of the ‘Fuji’ apple juice supplemented with 250 mg/L or 500 mg/L MT (melatonin) during storage with or without pasteurization. Data are means ± SD of three independent studies, and three duplicates for each analysis. Lower-case letters indicate significant differences among treatments based on LSD multiple comparison (p < 0.05).
Figure 5.
The effect of melatonin on the browning of ‘Fuji’ apple juice at different storage time. (a) Photo of the ‘Fuji’ apple juice supplemented with 250 or 500 mg/L MT (melatonin) during storage with or without pasteurization. (b) The browning index of the ‘Fuji’ apple juice supplemented with 250 mg/L or 500 mg/L MT (melatonin) during storage with or without pasteurization. Data are means ± SD of three independent studies, and three duplicates for each analysis. Lower-case letters indicate significant differences among treatments based on LSD multiple comparison (p < 0.05).
Figure 6.
The effect of melatonin of 250 or 500 mg/L on the antioxidant capacities of ‘Fuji’ apple juice. (a) and (b) The antioxidant capacities of ‘Fuji’ apple juice with 250 or 500 mg/L MT (melatonin) with or without pasteurization compared to the control respectively. Antioxidant capacity was evaluated by ABTS analysis. Data are means ± SD of three independent studies, and three duplicates for each analysis. Lower-case letters indicate significant differences among treatments based on LSD multiple comparison (p < 0.05).
Figure 6.
The effect of melatonin of 250 or 500 mg/L on the antioxidant capacities of ‘Fuji’ apple juice. (a) and (b) The antioxidant capacities of ‘Fuji’ apple juice with 250 or 500 mg/L MT (melatonin) with or without pasteurization compared to the control respectively. Antioxidant capacity was evaluated by ABTS analysis. Data are means ± SD of three independent studies, and three duplicates for each analysis. Lower-case letters indicate significant differences among treatments based on LSD multiple comparison (p < 0.05).
Figure 7.
Effects of melatonin on microorganism growth in the unpasteurized ‘Fuji’ apple juice (a), (b) and (c) The number of total colonies, moulds and yeasts in the unpasteurized ‘Fuji’ apple juice with 250 or 500 mg/L MT (melatonin), respectively. Data are means ± SD of three independent studies, and three duplicates for each analysis. Lower-case letters indicate significant differences among treatments based on LSD multiple comparison (p < 0.05).
Figure 7.
Effects of melatonin on microorganism growth in the unpasteurized ‘Fuji’ apple juice (a), (b) and (c) The number of total colonies, moulds and yeasts in the unpasteurized ‘Fuji’ apple juice with 250 or 500 mg/L MT (melatonin), respectively. Data are means ± SD of three independent studies, and three duplicates for each analysis. Lower-case letters indicate significant differences among treatments based on LSD multiple comparison (p < 0.05).
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, H.; Liu, X.; Chen, T.; Ji, Y.; Shi, K.; Wang, L.; Zheng, X.; Kong, J. Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice. Molecules 2018, 23, 521. https://doi.org/10.3390/molecules23030521
AMA Style
Zhang H, Liu X, Chen T, Ji Y, Shi K, Wang L, Zheng X, Kong J. Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice. Molecules. 2018; 23(3):521. https://doi.org/10.3390/molecules23030521
Chicago/Turabian StyleZhang, Haixia, Xuan Liu, Ting Chen, Yazhen Ji, Kun Shi, Lin Wang, Xiaodong Zheng, and Jin Kong. 2018. "Melatonin in Apples and Juice: Inhibition of Browning and Microorganism Growth in Apple Juice" Molecules 23, no. 3: 521. https://doi.org/10.3390/molecules23030521