Calycophyllum spruceanum (Benth.), the Amazonian “Tree of Youth” Prolongs Longevity and Enhances Stress Resistance in Caenorhabditis elegans

 , , , and
, , , and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Extract
2.2. DPPH
2.3. Total Phenolic Content
2.4. Chemical Characterization
2.5. C. elegans Strains and Maintenance
2.6. Survival Assay under Oxidative Stress
2.7. Intracellular ROS Accumulation
2.8. Quantification of Sod-3::GFP and Hsp-16::GFP Expression
2.9. Longevity Assay
2.10. Pharyngeal Pumping Rate
2.11. Body Length and Brood Size
2.12. Carbonyl Content
2.13. Antimicrobial Activity
2.14. Statistical Analyses
3. Results
3.1. Antioxidant Activity In Vitro and Total Phenolic Content
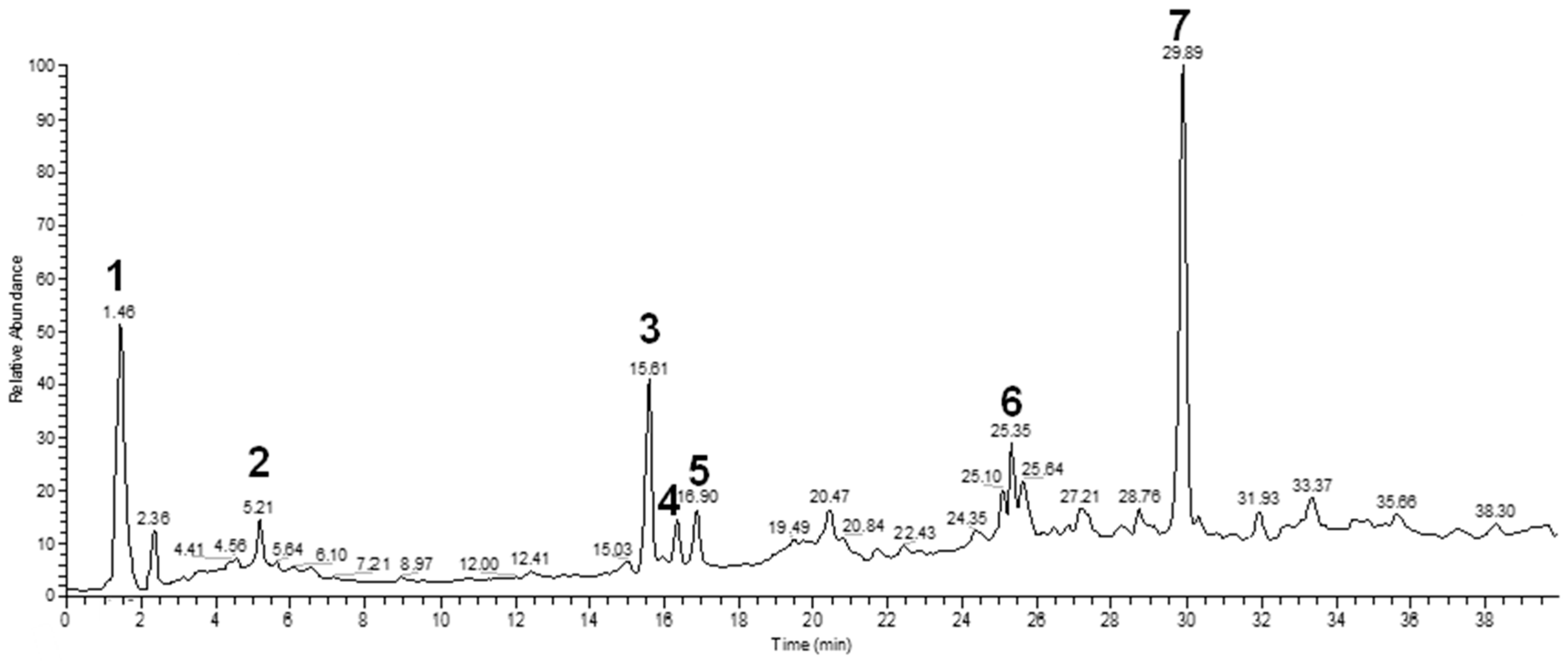
3.2. Chemical Characterization
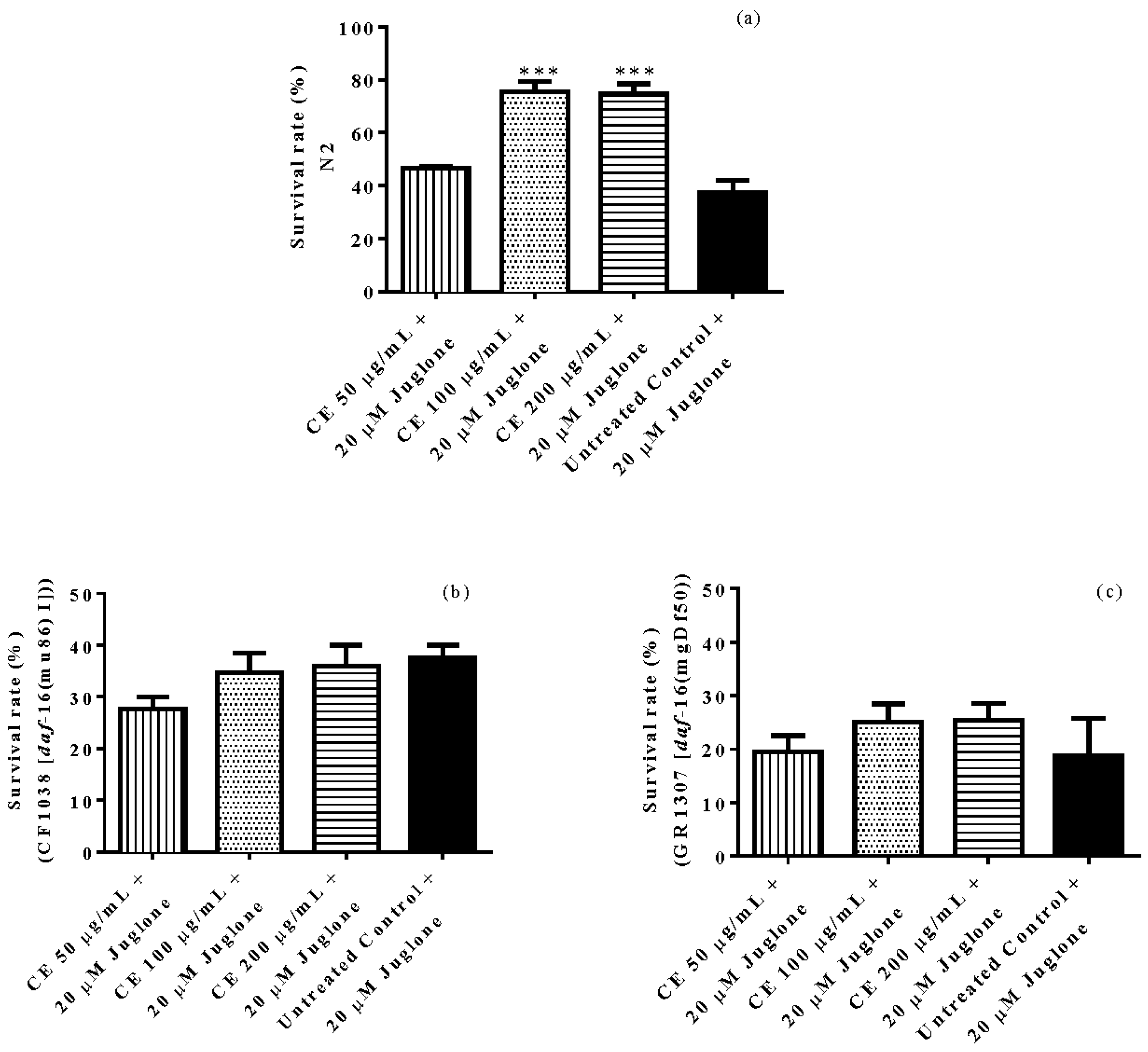
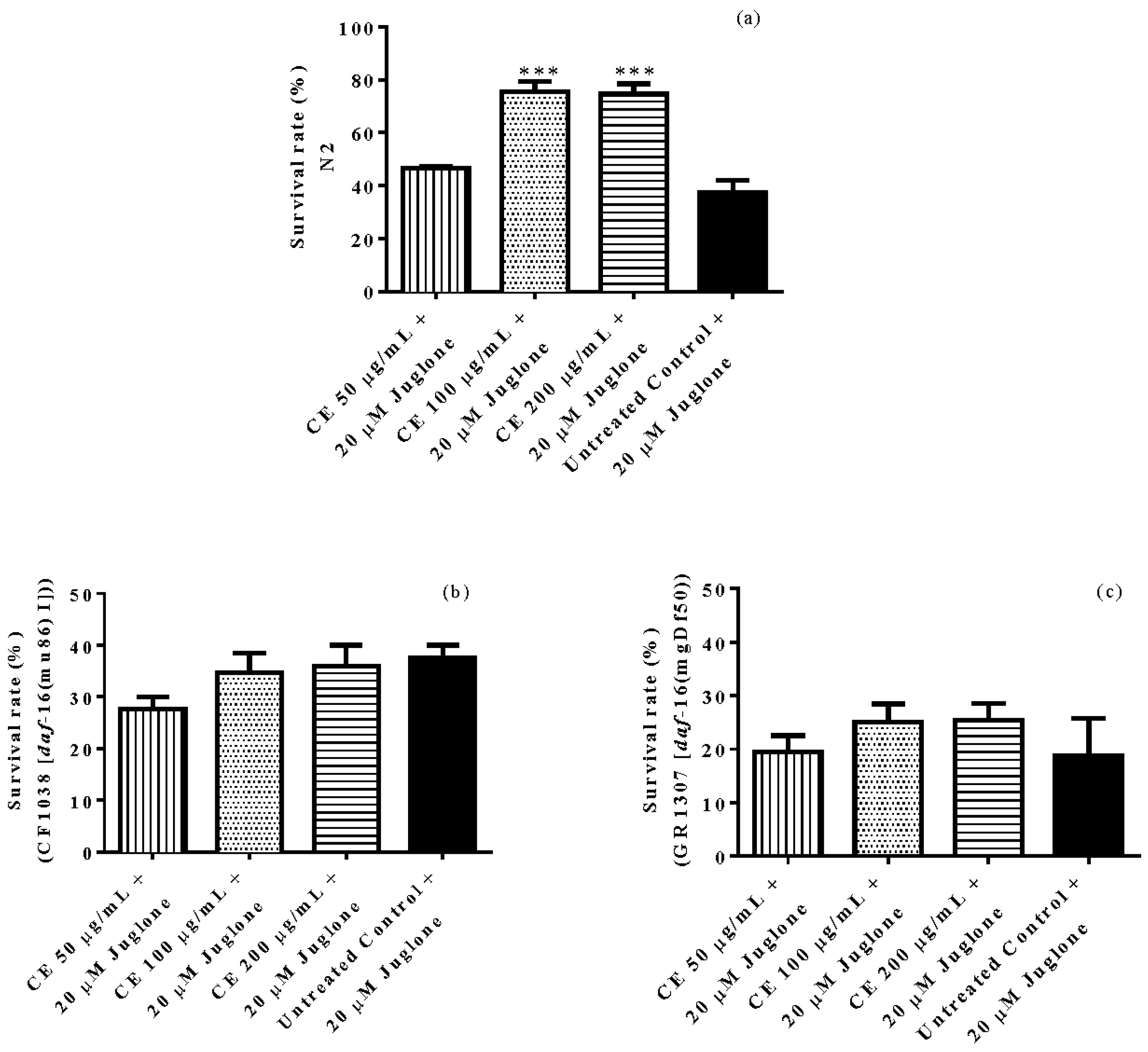
3.3. In Vivo Antioxidant Activity
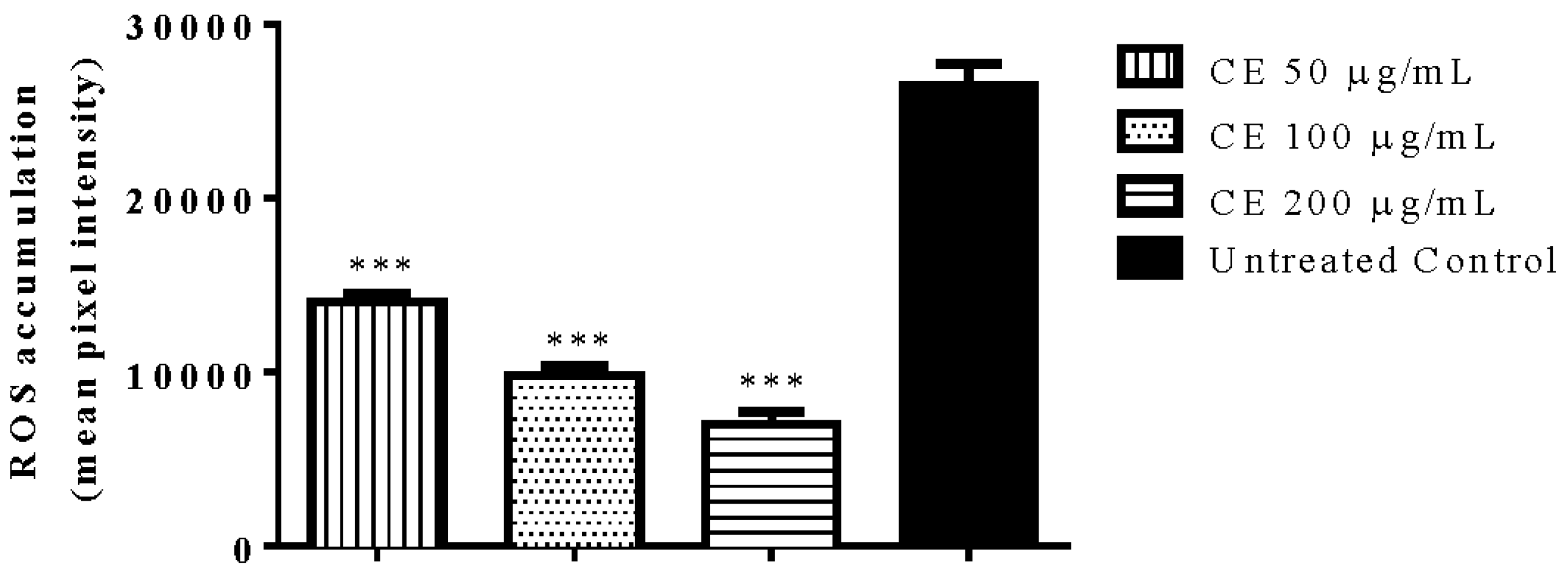
3.4. Quantification of Intracellular ROS Accumulation
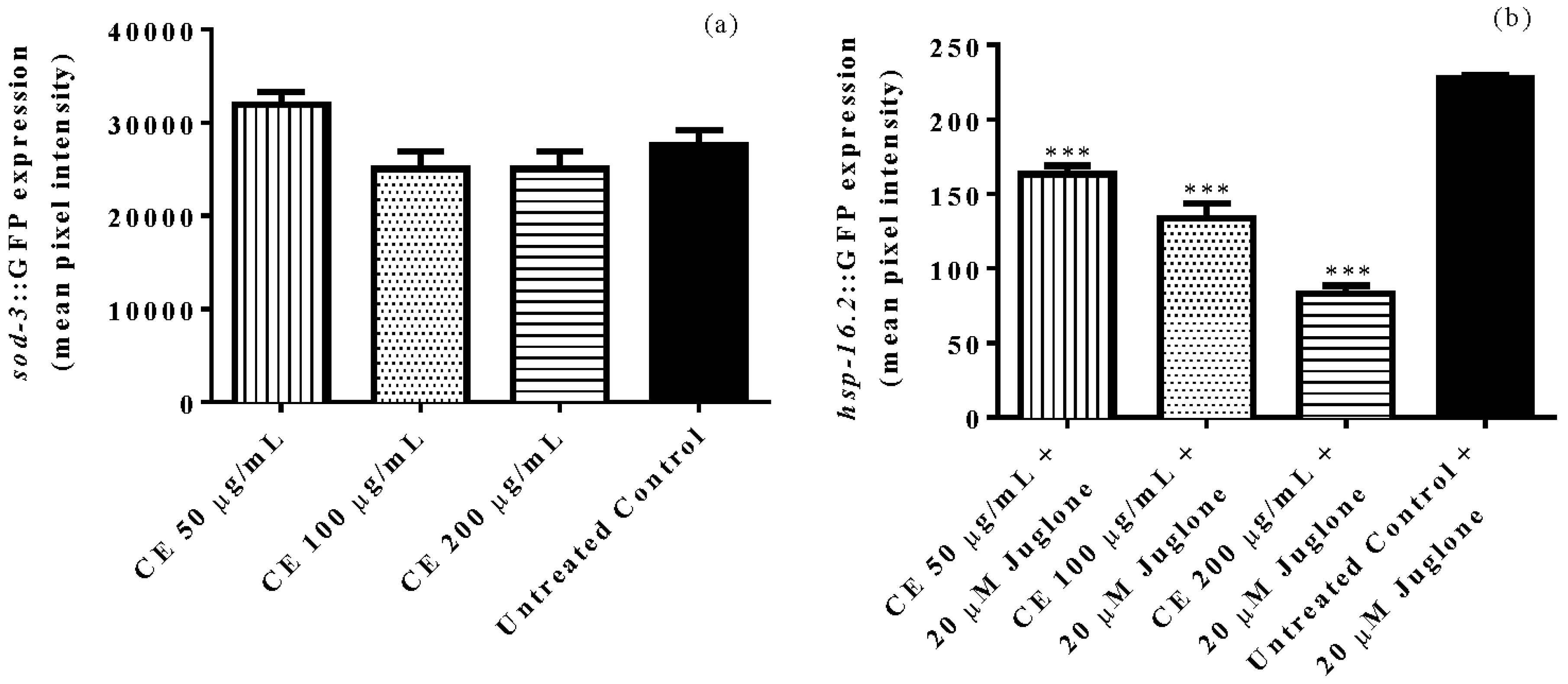
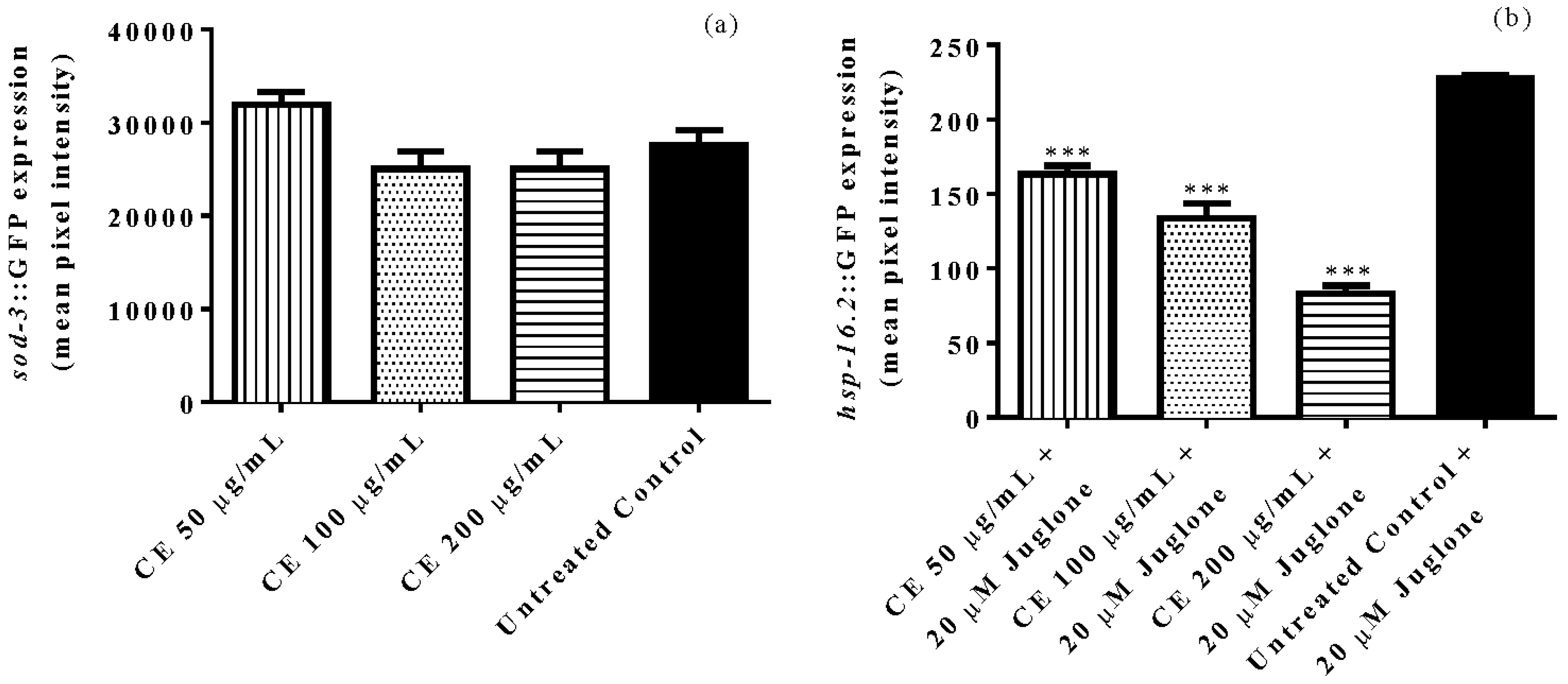
3.5. Quantification of Gene Expression of Sod-3 and Hsp-16.2
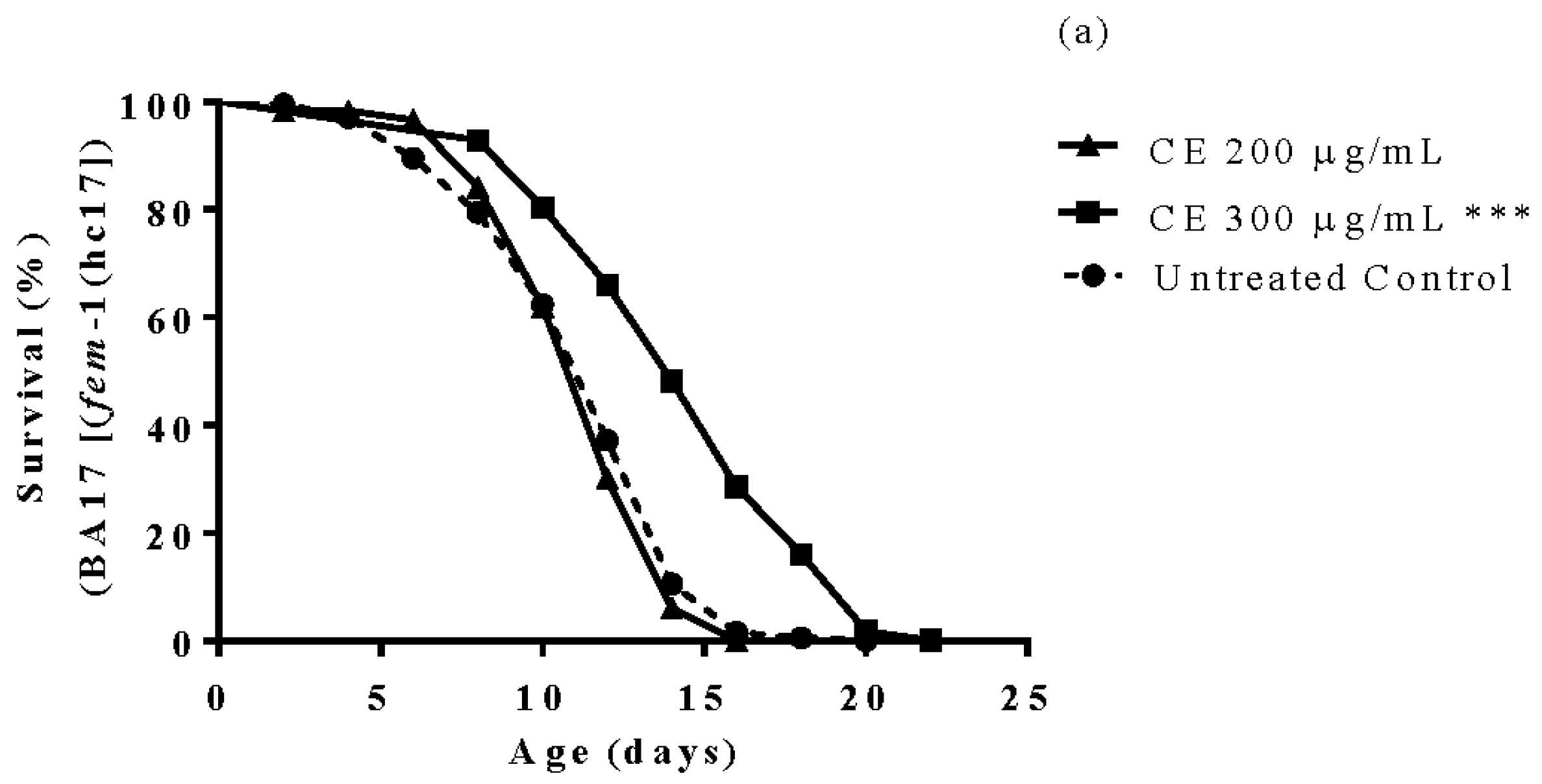
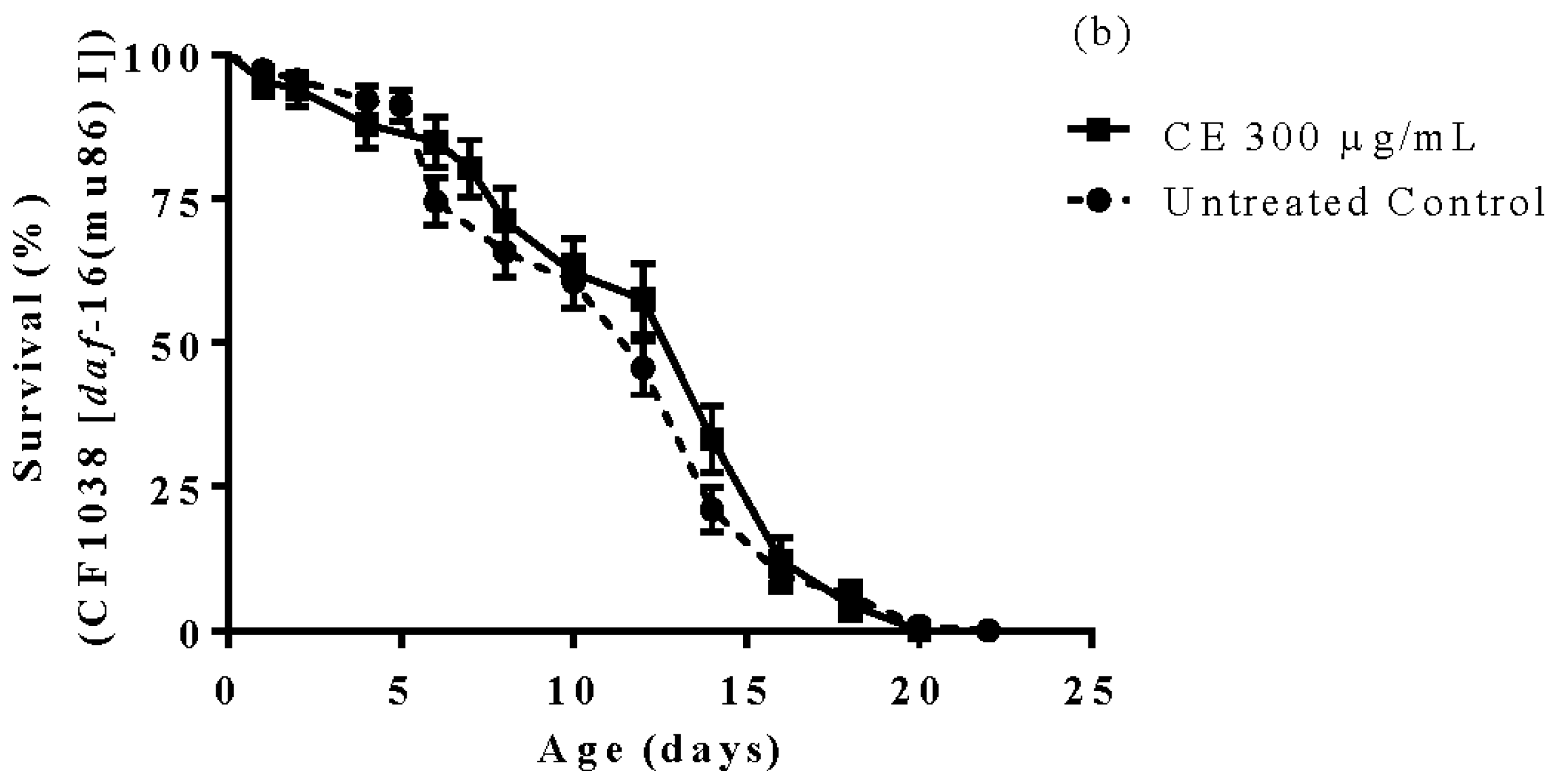
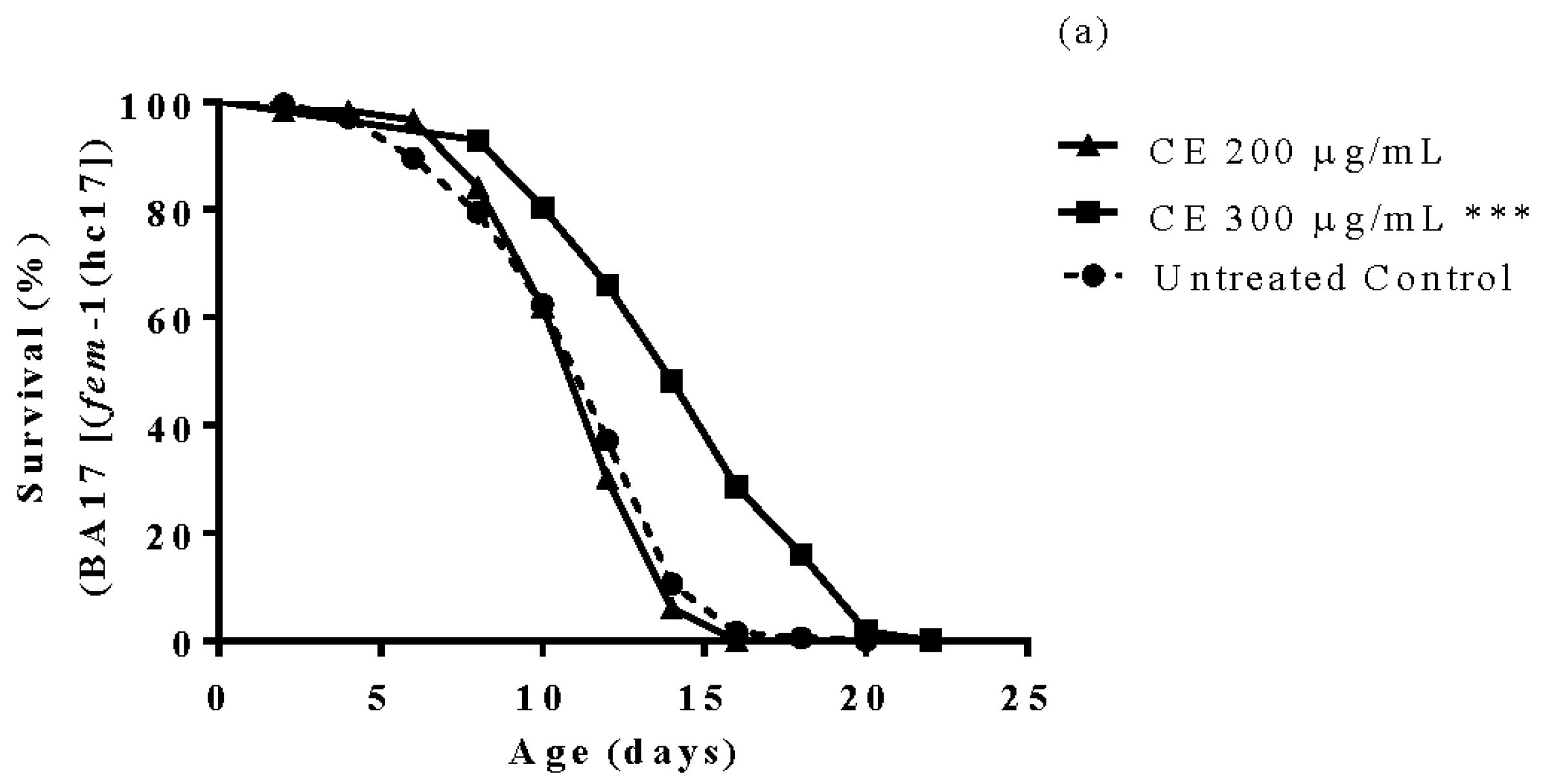
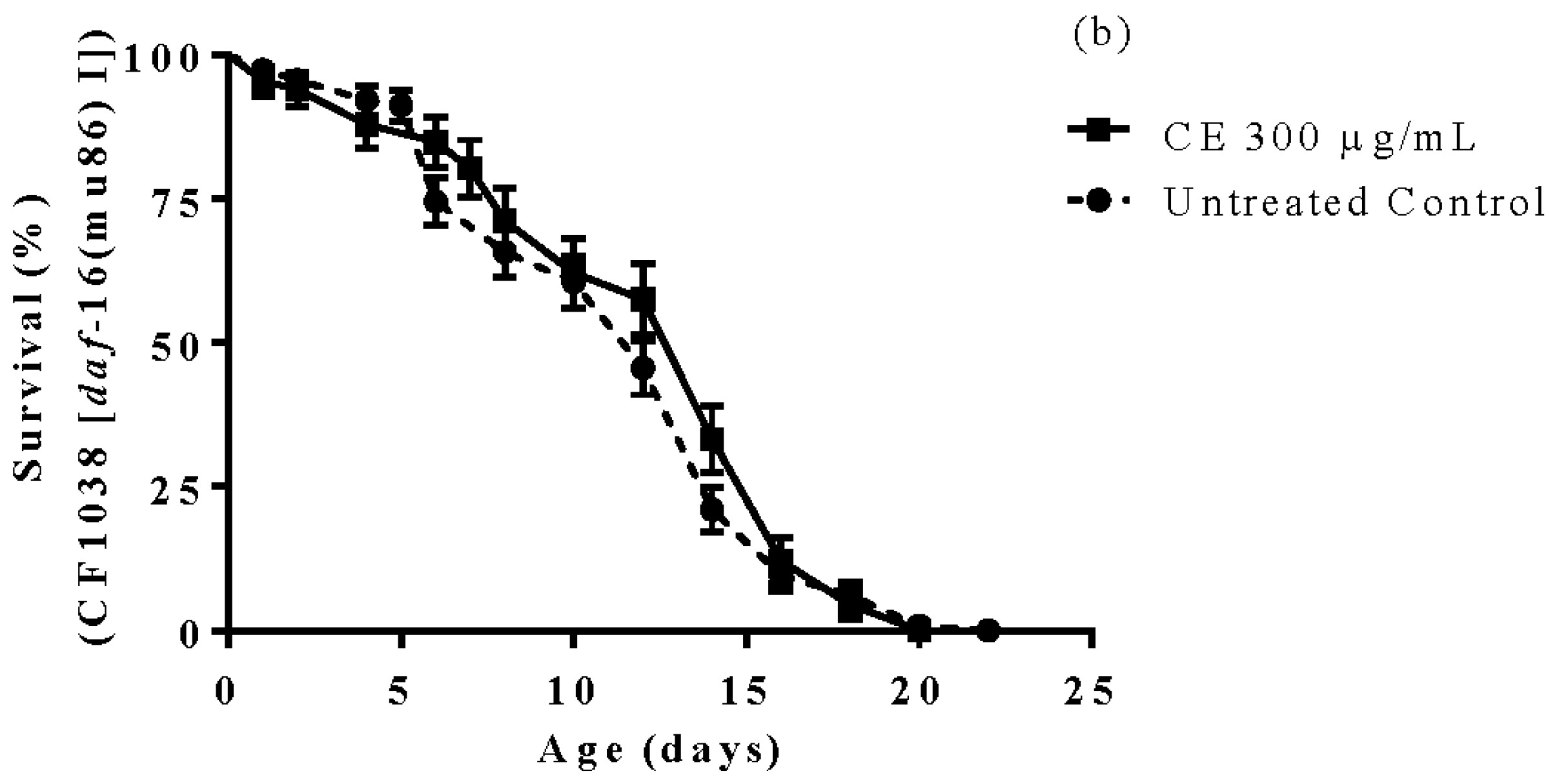
3.6. Longevity Study
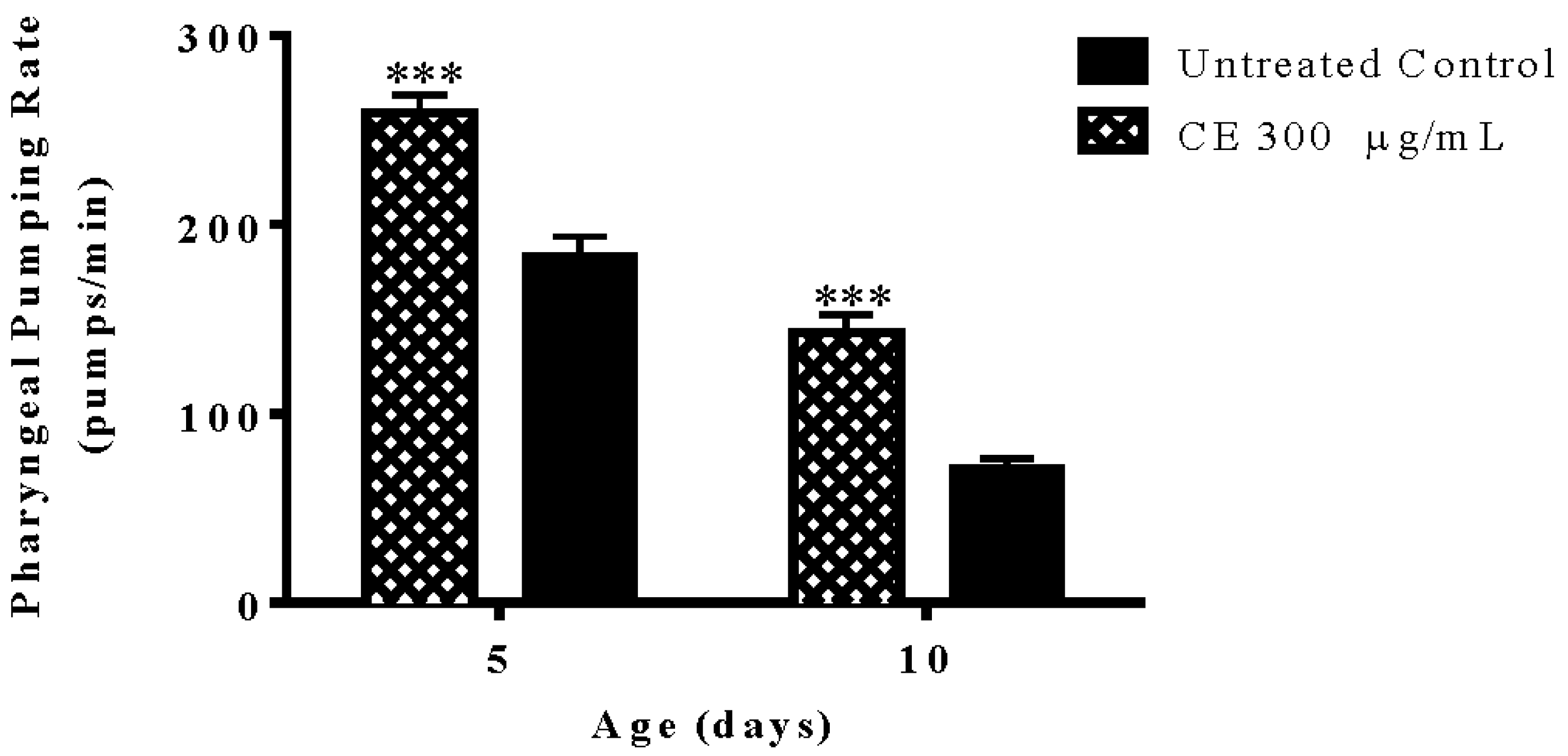
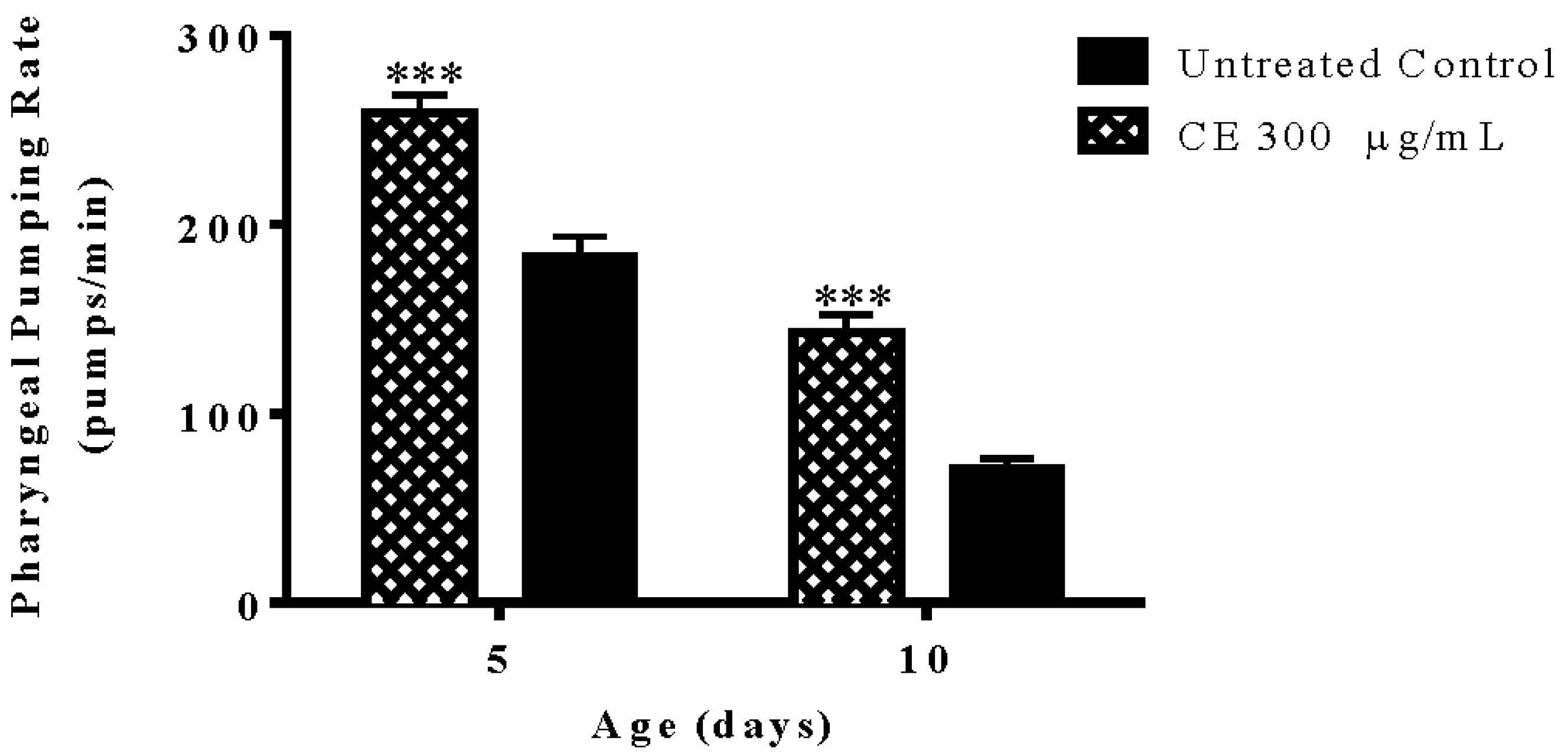
3.7. Pharyngeal Pumping Rate
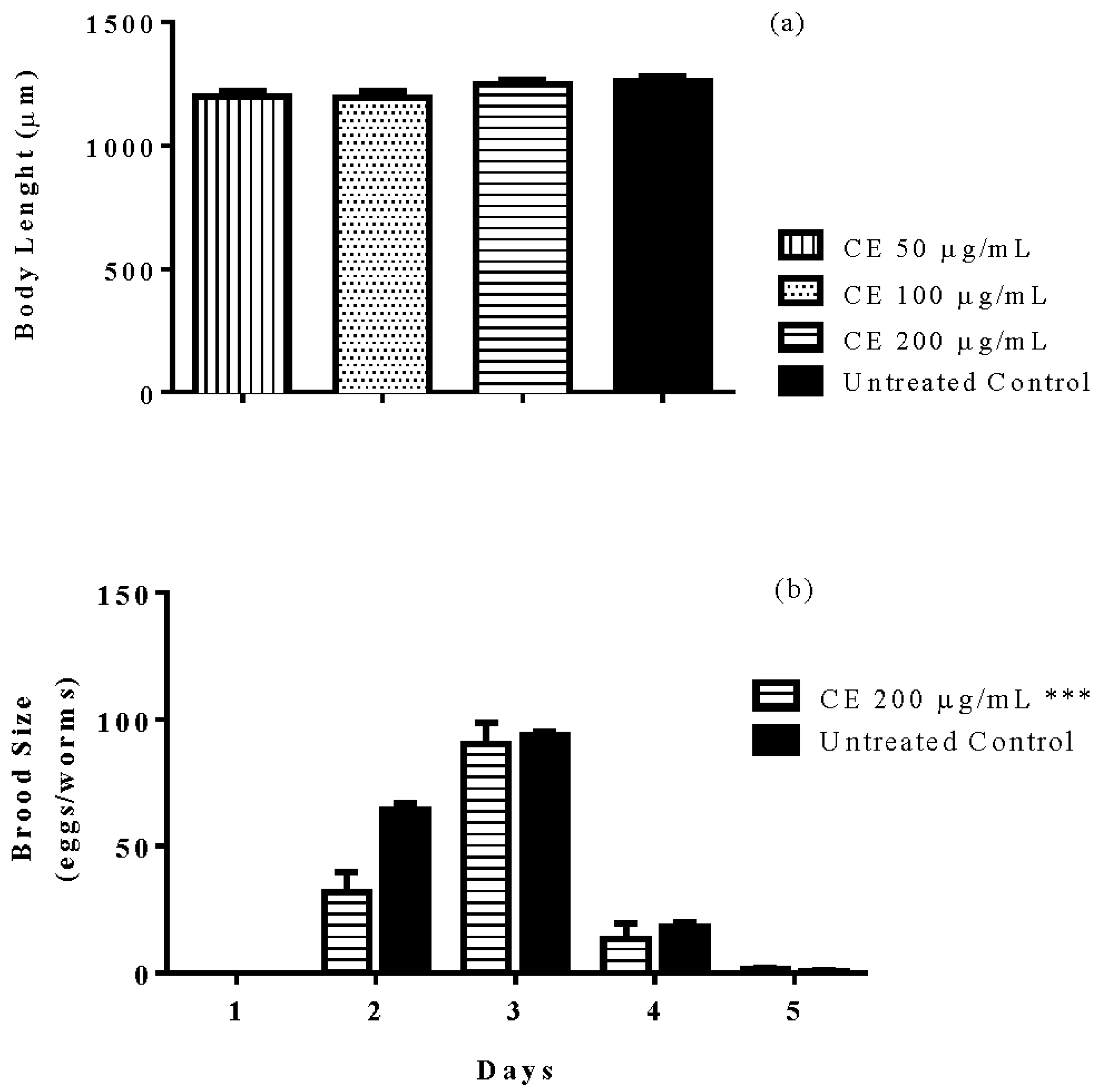
3.8. Brood Size and Body Length
3.9. Protein Carbonyl Content
3.10. Antimicrobial Activity
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duke, J.A.; Godwin, M.; Ottesen, A. Duke’s Handbook of Medicinal Plants of Latin America; CRQ Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Kvist, L.P.; Andersen, M.K.; Stagegaard, J.; Hesselsøe, M.; Llapapasca, C. Extraction from woody forest plants in flood plain communities in amazonian peru: Use, choice, evaluation and conservation status of resources. For. Ecol. Manag. 2001, 150, 147–174. [Google Scholar] [CrossRef]
- Weber, J.C.; Montes, C.S.; Ugarte, J.; Simons, T. Phenotypic selection of Calycophyllum spruceanum on farms in the Peruvian amazon: Evaluating a low-intensity selection strategy. Silvae Genet. 2009, 58, 172. [Google Scholar] [CrossRef]
- Tauchen, J.; Lojka, B.; Hlasna-Cepkova, P.; Svobodova, E.; Dvorakova, Z.; Rollo, A. Morphological and genetic diversity of Calycophyllum spruceanum (Benth) k. Schum (Rubiaceae) in Peruvian amazon. Agric. Trop. Subtrop. 2011, 44, 212–218. [Google Scholar]
- Brioso, P.S.T. Chanker in Calycophyllum spruceanum in Rio de Janeiro state. Summa Phytopathol. 2012, 38, 97. [Google Scholar] [CrossRef]
- Santos, A.; Ribeiro-Oliveira, J.; Carvalho, C. On the botany, ethnopharmacology, and chemistry of Calycophyllum spruceanum (Benth.) Hook. F. Ex k. Schum. Rev. Bras. Plantas Med. 2016, 18, 383–389. [Google Scholar] [CrossRef]
- Alexiades, M.N. Ethnobotany of the Ese Eja: Plants, Health, and Change in an Amazonian Society. Ph.D. Thesis, The City University of New York, New York, NY, USA, 1999. [Google Scholar]
- Wen, L.; Haddad, M.; Fernández, I.; Espinoza, G.; Ruiz, C.; Neyra, E.; Bustamante, B.; Rojas, R. Actividad antifúngica de cuatro plantas usadas en la medicina tradicional peruana: Aislamiento de 3′-formil-2′, 4′, 6′-trihidroxidihidrochalcona, principio activo de Psidium acutangulum. Revista de la Sociedad Química del Perú 2011, 77, 199–204. (In Spanish) [Google Scholar]
- Zuleta, L.M.C.; Cavalheiro, A.J.; Silva, D.H.S.; Furlan, M.; Young, M.C.M.; Albuquerque, S.; Castro-Gamboa, I.; da Silva Bolzani, V. Seco-iridoids from Calycophyllum spruceanum (Rubiaceae). Phytochemistry 2003, 64, 549–553. [Google Scholar] [CrossRef]
- de Vargas, F.S.; Almeida, P.D.; de Boleti, A.P.A.; Pereira, M.M.; de Souza, T.P.; de Vasconcellos, M.C.; Nunez, C.V.; Pohlit, A.M.; Lima, E.S. Antioxidant activity and peroxidase inhibition of amazonian plants extracts traditionally used as anti-inflammatory. BMC Complement. Altern. Med. 2016, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Portillo, A.; Vila, R.; Freixa, B.; Adzet, T.; Cañigueral, S. Antifungal activity of Paraguayan plants used in traditional medicine. J. Ethnopharmacol. 2001, 76, 93–98. [Google Scholar] [CrossRef]
- Ameen, M. Epidemiology of superficial fungal infections. Clin. Dermatol. 2010, 28, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Funasaki, M.; Barroso, H.d.S.; Fernandes, V.L.A.; Menezes, I.S. Amazon rainforest cosmetics: Chemical approach for quality control. Química Nova 2016, 39, 194–209. [Google Scholar] [CrossRef]
- Chen, W.; Sudji, I.R.; Wang, E.; Joubert, E.; van Wyk, B.-E.; Wink, M. Ameliorative effect of aspalathin from rooibos (Aspalathus linearis) on acute oxidative stress in Caenorhabditis elegans. Phytomedicine 2013, 20, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Wink, M. Green tea extract induces the resistance of Caenorhabditis elegans against oxidative stress. Antioxidants 2014, 3, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.; Roxo, M.; Krstin, S.; Röhrig, T.; Richling, E.; Wink, M. An anthocyanin-rich extract of acai (Euterpe precatoria Mart.) increases stress resistance and retards aging-related markers in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. Elegans; WormBook; Posted 11 February 2006; Available online: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 24 February 2018).
- Ashour, M.L.; El-Readi, M.Z.; Hamoud, R.; Eid, S.Y.; El Ahmady, S.H.; Nibret, E.; Herrmann, F.; Youns, M.; Tahrani, A.; Kaufmann, D. Anti-infective and cytotoxic properties of Bupleurum marginatum. Chin. Med. 2014, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wyk, B.-E.; Wink, M. (Eds.) Phytomedicines, Herbal Drugs, and Poisons; University of Chicago Press: Chicago, IL, USA, 2015. [Google Scholar]
- Strayer, A.; Wu, Z.; Christen, Y.; Link, C.D.; Luo, Y. Expression of the small heat-shock protein hsp 16–2 in Caenorhabditis elegans is suppressed by Ginkgo biloba extract EGb 761. FASEB J. 2003, 17, 2305–2307. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R. Heat shock proteins in long-lived worms and mice with insulin/insulin-like signaling mutations. Aging 2009, 1, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, H.; Roxo, M.; Röhrig, T.; Richling, E.; Wang, X.; Wink, M. Anti-aging and antioxidant potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans indicate a new utilization for roasted seeds of guarana. Medicines 2017, 4, 61. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Johnson, I.T.; Saltmarsh, M. Polyphenols: Antioxidants and beyond. Am. J. Clin. Nutr. 2005, 81, 215S–217S. [Google Scholar] [CrossRef] [PubMed]
- Pant, A.; Pandey, R. Bioactive phytomolecules and aging in Caenorhabditis elegans. Healthy Aging Res. 2015, 4, 1–15. [Google Scholar]
- Rothman, J.H.; Singson, A. Caenorhabditis Elegans: Cell Biology and Physiology; Academic Press: Waltham, MA, USA, 2012; Volume 107. [Google Scholar]
- Candore, G.; Balistreri, C.R.; Listi, F.; Grimaldi, M.P.; Vasto, S.; Colonna-Romano, G.; Franceschi, C.; Lio, D.; Caselli, G.; Caruso, C. Immunogenetics, gender, and longevity. Ann. N. Y. Acad. Sci. 2006, 1089, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, T.C. The Evolution of Aging: How New Theories will Change the Future of Medicine; Azinet: Crownsville, MD, USA, 2006. [Google Scholar]
- Harman, D. The biologic clock: The mitochondria? JAGS 1972, 20, 145–147. [Google Scholar] [CrossRef]
- Morselli, E.; Maiuri, M.; Markaki, M.; Megalou, E.; Pasparaki, A.; Palikaras, K.; Criollo, A.; Galluzzi, L.; Malik, S.; Vitale, I. Caloric restriction and resveratrol promote longevity through the sirtuin-1-dependent induction of autophagy. Cell Death Dis. 2010, 1, e10. [Google Scholar]
- Borra, M.T.; Smith, B.C.; Denu, J.M. Mechanism of human sirt1 activation by resveratrol. Int. J. Biol. Chem. 2005, 280, 17187–17195. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Rezaizadehnajafi, L.; Wink, M. Influence of resveratrol on oxidative stress resistance and life span in Caenorhabditis elegans. J. Pharm. Pharmacol. 2013, 65, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jie, G.; Zhang, J.; Zhao, B. Significant longevity-extending effects of egcg on Caenorhabditis elegans under stress. Free Radic. Biol. Med. 2009, 46, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, M.; Bonafè, M.; Franceschi, C.; Paolisso, G. Insulin/igf-i-signaling pathway: An evolutionarily conserved mechanism of longevity from yeast to humans. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1064–E1071. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, R.; Schaffitzel, E.; Hertweck, M. Endocrine signaling in Caenorhabditis elegans controls stress response and longevity. J. Endocrinol. 2006, 190, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.; Chang, J.; Gensch, E.; Rudner, A.; Tabtiang, R.A. C. Elegans mutant that lives twice as long as wild type. Nature 1993, 366, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Tissenbaum, H.A.; Ruvkun, G. An insulin-like signaling pathway affects both longevity and reproduction in Caenorhabditis elegans. Genetics 1998, 148, 703–717. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IC50 * |
|---|---|
| CE | 3.00 ± 0.07 |
| EGCG | 1.03 ± 0.06 |
| Vitamin C | ±0.04 |
| Peak | Rt | m/z [M + H]+ | Fragment Ions | Tentative of Identification | Chemical Structure |
|---|---|---|---|---|---|
| 1 | 1.48 | 329 | 311, 293, 275, 251, 209 | unkown | - |
| 2 | 5.21 | 405 | 387, 355, 323, 193, 167 | gardenoside |  |
| 3 | 15.61 | 319 | 301, 273, 259 | 5-hydroxymorin |  |
| 4 | 10.64 | 287 | 269 | unknown | - |
| 5 | 11.52 | 321 | 285, 257, 247, 221 | cyanidin |  |
| 6 | 11.78 | 305 | 287, 259, 221, 191, 175, 163 | taxifolin |  |
| 7 | 12.01 | 371 | 259, 241, 147, 129 | 5-hydroxy-6-methoxycoumarin 7-glucoside |  |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peixoto, H.; Roxo, M.; Koolen, H.; Da Silva, F.; Silva, E.; Braun, M.S.; Wang, X.; Wink, M. Calycophyllum spruceanum (Benth.), the Amazonian “Tree of Youth” Prolongs Longevity and Enhances Stress Resistance in Caenorhabditis elegans. Molecules 2018, 23, 534. https://doi.org/10.3390/molecules23030534
Peixoto H, Roxo M, Koolen H, Da Silva F, Silva E, Braun MS, Wang X, Wink M. Calycophyllum spruceanum (Benth.), the Amazonian “Tree of Youth” Prolongs Longevity and Enhances Stress Resistance in Caenorhabditis elegans. Molecules. 2018; 23(3):534. https://doi.org/10.3390/molecules23030534
Chicago/Turabian StylePeixoto, Herbenya, Mariana Roxo, Hector Koolen, Felipe Da Silva, Emerson Silva, Markus Santhosh Braun, Xiaojuan Wang, and Michael Wink. 2018. "Calycophyllum spruceanum (Benth.), the Amazonian “Tree of Youth” Prolongs Longevity and Enhances Stress Resistance in Caenorhabditis elegans" Molecules 23, no. 3: 534. https://doi.org/10.3390/molecules23030534