1. Introduction
It is generally accepted that the fatty acid composition of membrane phospholipids have a major impact on determining membrane biophysical and, thereby, physiological properties [
1].
Staphylococcus aureus is a Gram-positive bacterial pathogen that shows a significant growth environment-dependent plasticity in its membrane fatty acid composition [
2]. The fatty acids of
S. aureus are generally considered to be a mixture of saturated branched-chain fatty acids (BCFAs) and straight-chain fatty acids (SCSFAs) that are synthesized by fatty acid biosynthesis system II (FAS II). Some bacteriological media such as Mueller-Hinton broth (MHB) promote a high proportion of BCFAs, whereas others such as Tryptic Soy Broth (TSB) result in decreased BCFAs and increased SCSFAs. BCFAs fluidize the membrane and SCSFAs have the opposite effect [
1,
3,
4,
5]. The majority of BCFAs are anteiso- having a methyl group at the antepenultimate position while iso-fatty acids have a methyl group at the penultimate position.
S. aureus lacks a fatty acid desaturase enzyme that converts saturated fatty acids into unsaturated products [
2,
6], and, hence, BCFAs play a critical role in maintaining membrane fluidity. However, when cells are grown ex vivo in serum straight-chain unsaturated fatty acids (SCUFAs) become a significant fraction of the total fatty acid composition [
2], due to direct incorporation of exogenous preformed SCUFAs from serum into the phospholipid biosynthesis pathway [
6]. SCUFAs are known to significantly increase membrane fluidity [
1].
The membrane composition and properties of
S. aureus are further complicated by the unique production of the triterpene staphyloxanthin carotenoid responsible for the typical golden color of
S. aureus colonies. Staphyloxanthin is thought to be important in the life of
S. aureus decreasing membrane fluidity [
2,
7,
8,
9,
10,
11,
12,
13], influencing membrane permeability [
8], susceptibility to oxidative stress and neutrophil killing [
14,
15,
16]. Thus staphyloxanthin biosynthesis has been explored as a potential drug target [
16]. Fatty acid biosynthesis is one of the highly energy demanding biosynthetic pathways and acetyl-CoA is the common precursor for both fatty acid and staphyloxanthin synthesis [
17]. Recently, we found a correlation between a high content of the membrane fluidizing fatty acids (BCFAs and SCUFAs) and an increase in staphyloxanthin content, possibly to counterbalance potential membrane hyperfluidity conferred by BCFAs or SCUFAs [
2]. Similarly, a
S. aureus mutant in
brnQ1 that was defective in transport of BCFA precursor amino acids had higher anteiso-fatty acids and increased pigmentation [
18]. Here, we were interested in probing further the relationship between carotenoid and fatty acid production and membrane biophysical properties in pigmented (Pig1) and non-pigmented (Pig1Δ
crtM and MSHR1132) staphylococcal strains.
S. aureus Pig1Δ
crtM is deleted in the
crtM gene that encodes a key enzyme in staphyloxanthin production [
14]. Strain MSHR1132 is a close relative of
S. aureus that naturally lacks the carotenoid biosynthesis operon (
crtOPQMN) that is known as
Staphylococcus argenteus (the silver
Staphylococcus, [
19,
20].
The level of staphyloxanthin production varies between strains and under different environmental conditions and the role of the pigment in the life of
S. aureus remains somewhat enigmatic. The alternative sigma factor SigB is a positive regulator of expression of the
crtMNOPQ operon [
21], and direct or indirect effects on the expression and activity of SigB regulate pigment production in the bacterium. A cold-shock protein (CspA) [
22] and an aeration sensing response regulator (AirR) [
23] have been shown to positively affect SigB activity and increase pigmentation. Mutations and/or altered activities of some regulators (such as SarA, Agr, argR) and ClpP protease may also affect SigB expression altering pigmentation [
17,
24]. Also, a transfer-messenger antisense RNA has been reported that negatively regulates the
crtMN transcript [
25]. Additionally, a carbohydrate catabolite regulator (CcpE) has been shown to regulate pigment production without affecting
crtM transcription [
26,
27]. On the other hand, addition of fatty acids or mevalonate in the growth medium [
28], blocking polyprenyl synthetase [
24] and inactivation of the tricarboxylic acid (TCA) cycle [
17] have been reported to increase pigmentation. A
S. aureus pyruvate dehydrogenase mutant defective in conversion of pyruvate to acetyl CoA had decreased staphyloxanthin production, whereas a branched-chain α-keto acid dehydrogenase mutant had decrease production of C4 and C5 iso and anteiso CoA precursors of BCFAs showed increased pigmentation [
29].
The membrane lipid composition of S. aureus is complex due to the variable production and incorporation of BCFAs, SCSFAs and SCUFAs, as well as being overlain with the variable production of staphyloxanthin. In earlier work, we observed an association between increased content of membrane fluidizing BCFAs and SCUFAs and increased content of membrane rigidifying staphyloxanthin. In this work we probed this potential relationship further through studies of fatty acid composition, staphyloxanthin content, and membrane fluidity in a carotenoid-deficient S. aureus mutant and a naturally occurring staphylococcal species closely related to S. aureus that had lost the crt carotenoid biosynthesis operon. We found no evidence that carotenoid biosynthesis responded to membrane fatty acid composition. Carotenoid deficiency led to membranes with increased fluidity. However, medium carbon source influenced membrane fatty acid composition and staphyloxanthin content, with evidence that excess acetyl CoA led to increased staphyloxanthin production.
3. Discussion
Fatty acids in phospholipids play major roles in determining membrane biophysical properties.
S. aureus synthesizes and alters the balance of membrane fluidizing BCFAs (mostly anteiso C15:0 and anteiso C17:0) and rigidifying SCSFAs under different growth conditions. The bacterium can take up SCUFAs from environment and incorporate them into phospholipids [
2,
6]. Many
S. aureus strains produce a characteristic membrane component, staphyloxanthin, which is a triterpenoid pigment with an anteiso C15:0 at the C6-position of the glucose residue in glycosyl-4,4′-diaponeurosporenoate [
10,
12,
13,
33,
34]. The pigment has been shown to provide rigidity in the membrane [
2,
7,
8,
9,
10,
11]. Here, we grew
S. aureus strains Pig1 (pigmented), Pig1Δ
crtM (non-pigmented), and
S. argenteus (a close relative of
S. aureus, non-pigmented) in various culture media and analyzed the relationship between fatty acid composition and pigmentation. We observed that production of staphyloxanthin does not influence the fatty acid composition but helps maintain membrane fluidity. Notably, fatty acid composition and pigmentation in the bacterium were markedly affected by supplementation of glucose or acetate in the growth medium. We believe that staphyloxanthin production and fatty acid composition are regulated metabolically.
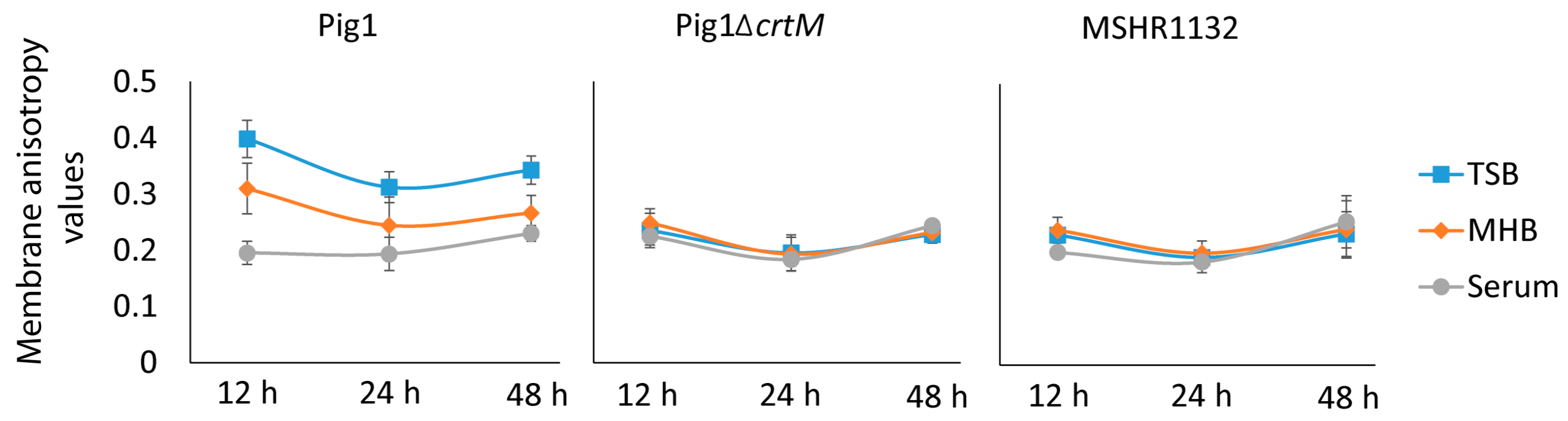
The relationship of fatty acid composition and staphyloxanthin production is complex. As observed previously [
2] in two different strains of
S. aureus, here we observed that
S. aureus Pig1 grown in MHB at log phase had higher BCFAs and higher pigmentation than those grown in TSB. Further, serum-grown cells had a large proportion of SCUFAs and higher pigmentation than the TSB-grown cells. Additionally, we observed that the 12 h-grown cells in TSB and MHB had higher proportion of BCFAs and were more pigmented than the respective log phase cells indicating a direct relationship of BCFAs and pigmentation in
S. aureus. However, despite having lower BCFAs, the TSB-grown cells at 12 h had markedly higher pigmentation than those grown in MHB. Further, despite having a substantial proportion of SCUFAs the serum-grown cells at 12 h had least pigmentation. Additionally, acetate-supplementation in culture broths increased pigmentation and decreased BCFAs in
S. aureus. These observations indicate that there may not always be a direct relationship of fatty acid composition and pigmentation in
S. aureus. We observed that the fatty acid composition in pigmented and non-pigmented strains were similar. Further, the genes for biosynthesis and regulation of fatty acids and staphyloxanthin in the strains were appear to be independently regulated. Taken together our observation suggest that the relationship of fatty acid composition and pigmentation in
S. aureus is complex.
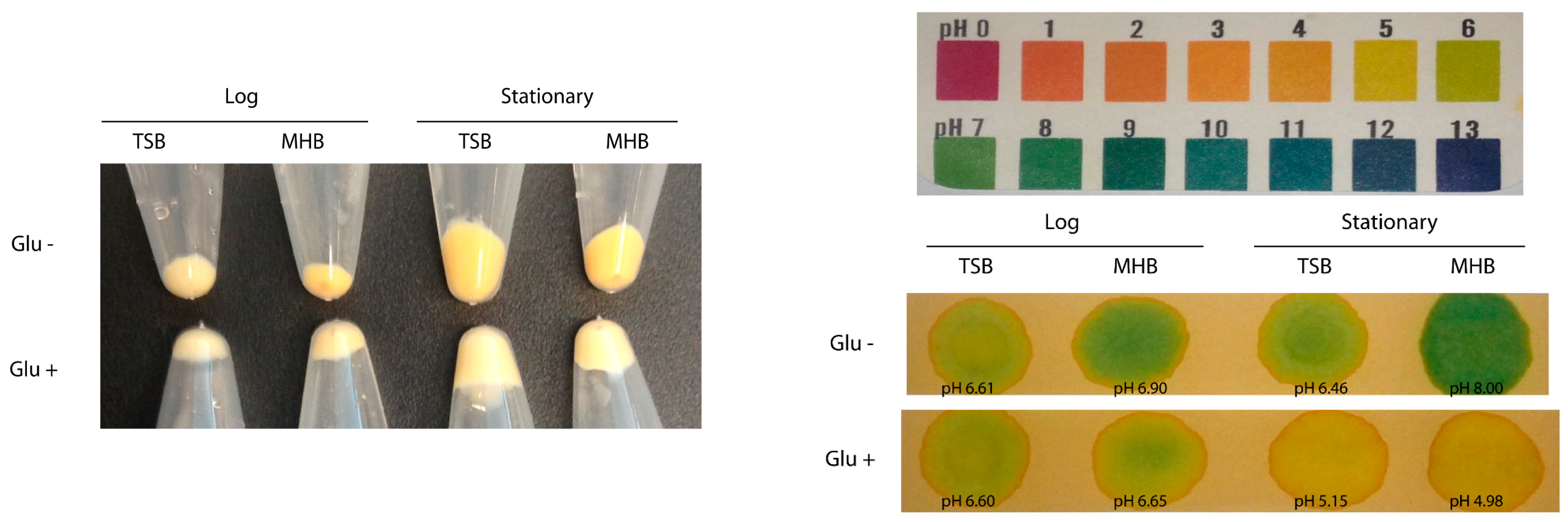
S. aureus acetate metabolism determines the extent of its pigment production.
S. aureus continuously secretes acetate in glucose-containing medium and consumes acetate when glucose is exhausted [
31]. We observed that supplementation of media with glucose decreased pigmentation while acetate-supplementation increased pigmentation. Further, glucose-supplementation increased acetate concentration in the culture supernatants even at stationary phase where the cells preferentially consume acetate otherwise. Since glucose catabolism yields acetyl-CoA and the excess acetyl-CoA is converted into acetate, which is continuously secreted out of cells [
31], the secretion of the precursor molecule might reduce pigmentation in the cells in glucose-supplemented media. On the other hand, supplementation of acetate in the media might provide surplus acetyl CoA in the cells leading to higher pigment production. Hence, these observations indicate that acetate consumption directly correlates to pigmentation in
S. aureus.
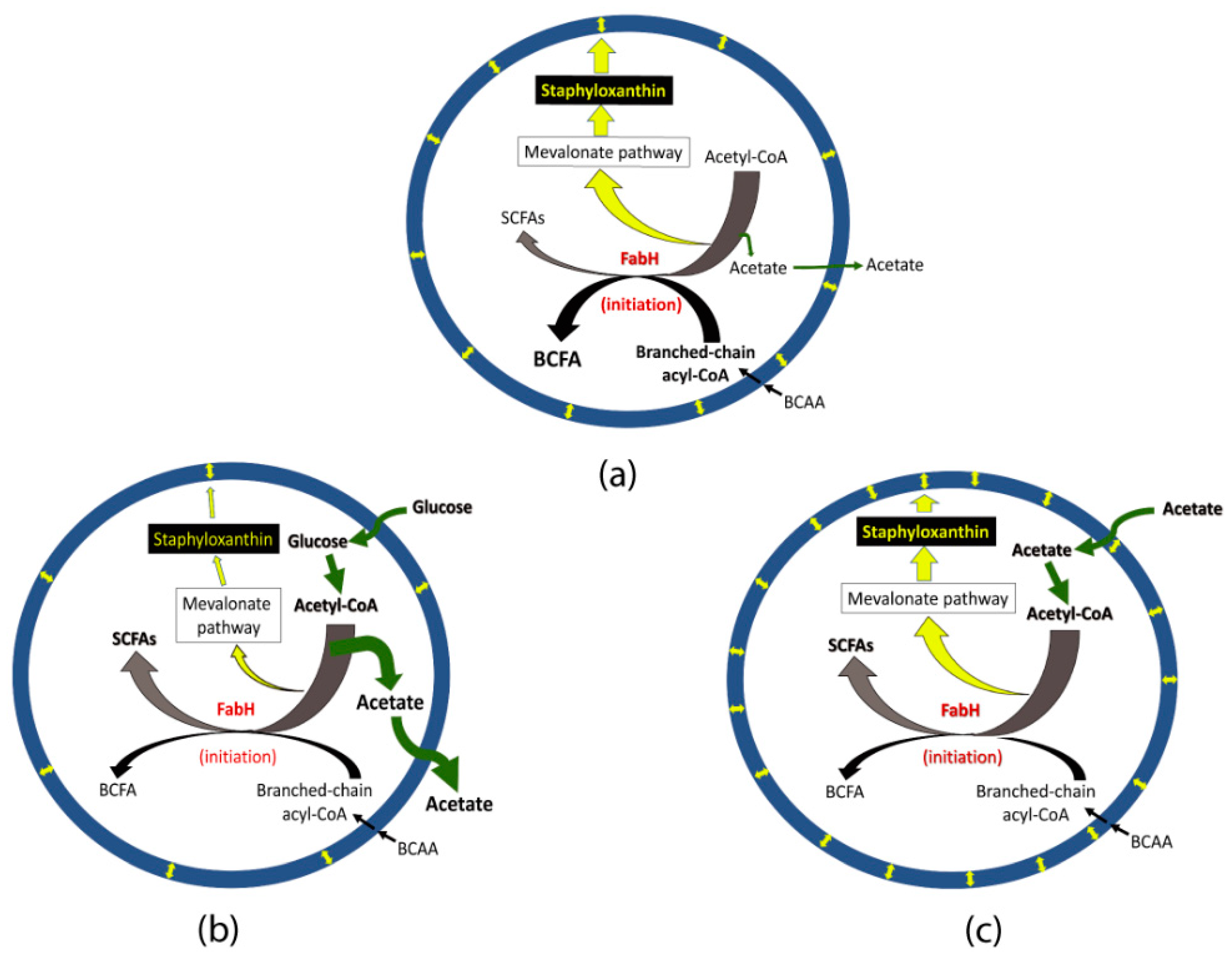
Acetate metabolism may affect initiation of fatty acid biosynthesis leading to alteration in BCFA and staphyloxanthin production. As explained above, increased BCFAs is usually associated with increased pigmentation in glucose-lacking media (
Figure 7a); however, an increased pigmentation may not result in increased BCFAs. This notion is supported by an observation that a
S. aureus pdh mutant impaired in acetyl-CoA production had both significantly reduced staphyloxanthin and significantly higher BCFAs (80%; wild type, 50%) [
29]. Similarly, a
bkd mutant had significantly higher pigmentation but had significantly lower BCFAs (31%) [
29]. Since, BCFA synthesis requires branched-chain acyl-CoA (precursors synthesized by the BKD complex) and fatty acid biosynthesis initiation enzyme (FabH) prefers branched-chain acyl-CoA to acetyl-CoA [
35], the scarcity of acetyl-CoA in the
pdh mutant might be a condition where the FabH enzyme synthesizes more BCFAs. On the other hand, a decreased synthesis of BCFAs in the
bkd mutant might result in FabH enzyme utilizing more acetyl-CoA in the initiation step leading to increased SCSFAs than in the parent strain. Based on this assumption of acetyl-CoA consumption in a
bkd mutant, a continuous production (cellular availability) of acetyl-CoA in glucose-supplemented broths might affect FabH activity leading to increased SCSFAs (
Figure 7b). Similarly, a continuous uptake of acetate and subsequent availability of surplus acetyl-CoA could affect FabH activity leading to increased SCSFAs in the acetate-supplemented medium (
Figure 7c).
An increased flow of acetyl CoA causes a higher pigmentation in
S. aureus Pig1. The notion of increased flow of acetyl-CoA towards the mevalonate pathway leading to increased pigmentation is corroborated by the findings of addition of fatty acids or mevalonate in the growth medium [
28], blocking polyprenyl synthetase [
24] and inactivation of the TCA cycle [
17] increase pigmentation in
S. aureus. Inactivation of the TCA cycle provides more acetyl-CoA in the cytoplasmic pool that can be used in the mevalonate pathway [
17]. Notably, the mevalonate pathway also yields undecaprenyl phosphate that is required to synthesize peptidoglycan, teichoic acid, and capsule [
36] and menaquinone. Interestingly, most (~90%) of the undecaprenyl phosphate is converted and stored as an inactive form, undecaprenol, which is recycled back to undecaprenyl phosphate when necessary [
36,
37]. Therefore, we believe that consumption of most of the surplus acetyl-CoA by the mevalonate pathway is committed to staphyloxanthin production.
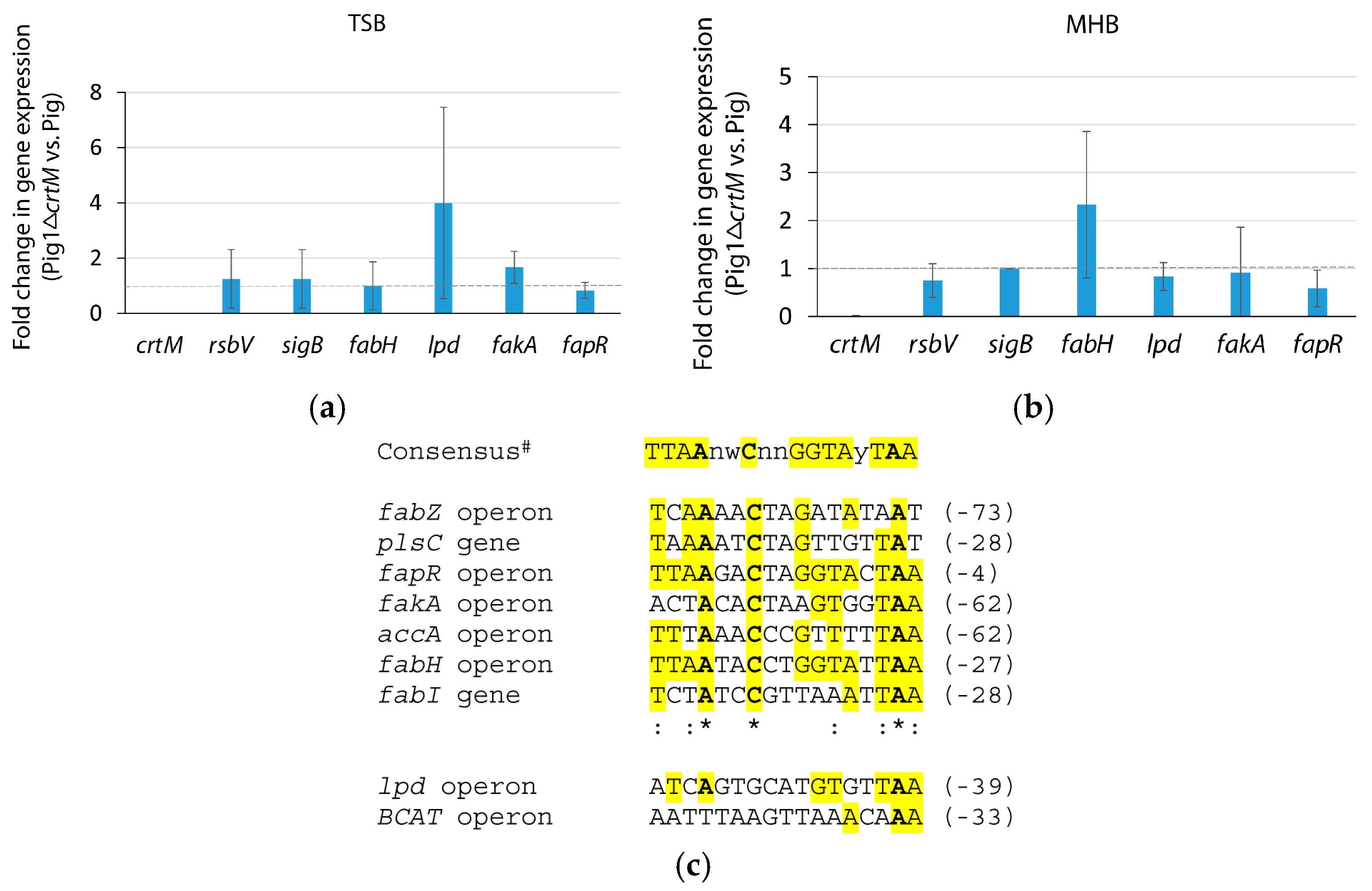
On the other hand, a highly pigmented
bkd mutant had an unaltered level of
crtM transcript compared to the parent strain indicating an unaltered SigB (that regulates the
crt operon) activity in the mutant [
29]. Hence, an increased flow of acetyl-CoA towards the mevalonate pathway could be responsible for increased pigmentation in the
bkd mutant [
29]. In this study, we supplemented broths with glucose (140 mM) and acetate (100 mM) to alter the carbon flow in a clinical isolate Pig1. A decreased pigmentation with glucose-supplementation and an increased pigmentation with acetate-supplementation was observed in both log- and stationary-phase cells indicating that the alterations in pigmentation might not be influenced by SigB (the activity of which changes over growth cycle, [
38,
39,
40]). Taken altogether, an increased flow of carbon to the mevalonate pathway might cause an increase in pigmentation in
S. aureus Pig1 having higher proportion of BCFAs.
4. Materials and Methods
S. aureus strains and growth media. A markedly yellow pigmented clinical
S. aureus isolate (Pig1) and its non-pigmented
crtM-knockout mutant (Pig1Δ
crtM) [
14], kindly supplied by Dr. George Liu, University of California San Diego, La Jolla, CA, and a naturally
crt-operon lacking clinical
S. argenteus strain (Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ), Leibniz-Institut, Germany [
19,
20]) were used in this study (
Table 9). Other
S. aureus strains used were SH1000 [
41], Newman [
42], COL [
43] and MW2 [
44] (
Table 9). The growth media (culture to volume ratio of 1:5) used were Bacto
TM Tryptic soy broth (TSB; Becton, Dickinson and Company, Baltimore, MD, USA), Bacto
TM Mueller-Hinton broth (MHB; Becton, Dickinson and Company, MD), Difco
TM Luria-Bertani (LB, Miller; Becton, Dickinson and Company) medium and whole human serum (BioreclammationIVT, New York, NY, USA). Human serum was heated to 56 °C for 30 min before using.
Determination of growth patterns of S. aureus strains grown in TSB, MHB and human serum. Overnight grown strains Pig1, Pig1ΔcrtM and MHSR1132 were inoculated into TSB, MHB and serum, grown for 48 h and growth was monitored by measuring turbidity (OD600) at intervals.
Determination of fatty acid compositions in
S. aureus strains grown in TSB, MHB and human serum. Strains Pig1, Pig1Δ
crtM and MHSR1132 were grown to an optical density (OD
600) of about 0.8 in TSB, MHB and serum, were harvested (3000×
g for 5 min), washed with cold water, and the pellets were sent as frozen sample with overnight delivery for fatty acid methyl ester (FAME) analysis at MIDI, Inc. (Newark, Delaware, USA). At MIDI, fatty acids were extracted and transesterified to fatty acid methyl esters, which were then separated and identified by gas-chromatography [
45].
Determination of staphyloxanthin production in
S. aureus strains grown in TSB, MHB and human serum. Staphyloxanthin content in the
S. aureus cells was determined as described previously [
23,
46]. Briefly, TSB-, MHB- and serum-grown Pig1 cells were pelleted (3000×
g for 5 min at 4 °C) at OD
600 of ~0.8, and then at 12 h, 24 h and 48 h. Each pellet was washed with water and resuspended in 0.8 mL of 100% methanol and incubated at 65 °C for 10 min with vortexing every three minutes. OD
465 was measured for each methanol extract and the staphyloxanthin-content was determined by dividing OD
465 by cell mass (OD
600 1.0 corresponds to 0.39 mg dry wt./mL; Bieber and Wilkinson, 1984). Three independent assays were performed for each category.s
Determination of membrane fluidity of
S. aureus strains grown in TSB, MHB and human serum. The membrane fluidities of the strains Pig1, Pig1Δ
crtM and MHSR1132 were determined using 1,6-diphenyl-1,3,5-hexatriene (DPH, Sigma-Aldrich, St. Louis, MO, USA) as described previously [
47] with some modifications. Briefly, the strains were grown in TSB, MHB and serum, pelleted, washed, resuspended in phosphate buffered saline (PBS, pH 7.5) to an OD
600 of ~0.8, DPH (5 µM) was added and the suspension was incubated at 30 °C in a water bath. All steps involving DPH were carried out in the dark. Fluorescence polarization emitted by the fluorophore was measured using a PTIModel Quanta Master-4 Scanning Spectrofluorometer at an excitation wavelength of 360 nm and an emission wavelength of 430 nm. The experiments were performed with three separate fresh batch cultures of cells grown to early-exponential phase, 12, 24 and 48 h.
Determination of expression of genes for staphyloxanthin and fatty acid biosynthesis and their regulation. The level of expression of the genes encoding key enzymes for staphyloxanthin and fatty acid biosynthesis and their regulation were determined by quantitative Real-time Polymerase Chain Reaction (qRT-PCR). Briefly, overnight grown cultures of the strains Pig1 and Pig1Δ
crtM were inoculated in TSB or MHB, grown to OD
600 of ~0.8, harvested, and total RNA was extracted and purified using a RNAeasy Mini Kit (Qiagen). cDNA was prepared from 300 ng of each RNA using the High Capacity RNA-to-cDNA kit (Applied Biosystems, Foster City, CA, USA). qRT-PCR reactions were conducted using the DyNAmo Flash SYGR Green Kit (Thermo Scientific, Waltham, MA, USA) with gene-specific primers (
Supplementary Materials Table S1) along with for 16SrRNA as an internal control of gene expression. PCR was run using the ABI 7300 Real-Time PCR system. The levels of expression of the genes were calculated by the Comparative Ct method. The reactions were run in triplicates for each set and each experiment was repeated with three independent biological replicates.
Determination of staphyloxanthin in Pig1 grown in glucose or acetate containing LB. Overnight grown culture was inoculated into (i) LB; (ii) LB with MOPS [3-(N-morpholino)propanesulfonic acid] buffer, pH 7.5; (iii) LB with 140 mM glucose; (iv) LB with MOPS and 140 mM glucose; and (v) LB with MOPS and 100 mM acetate. The cells were grown at 37 °C, harvested at 3 h and 12 h, and staphyloxanthin content was determined as described previously. Each experiment was repeated at least three times.
Determination of acetate content in culture supernatants. Culture supernatants were collected from strain Pig1 grown in TSB, MHB, 140 mM glucose-supplemented TSB and MHB, and harvested (10,000× g for 10 min at 4 °C) at ~3 h (OD600 ~0.8) and at 12 h. The supernatants were filtered through 0.2 µ filter (Millipore) and acetate concentration was determined with an acetate standard calibration curve prepared using an Acetic Acid Assay Kit (Megazyme International Ireland, Wicklow, Ireland). Additionally, staphyloxanthin content in the cell pellets were determined. Each experiment was repeated three times.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}