
Antifungal Activity of an Abundant Thaumatin-Like Protein from Banana against Penicillium expansum, and Its Possible Mechanisms of Action

Abstract
:1. Introduction
2. Results and Discussion
2.1. Purification and Identification of Thaumatin-Like Protein from Banana (Ban TLP)
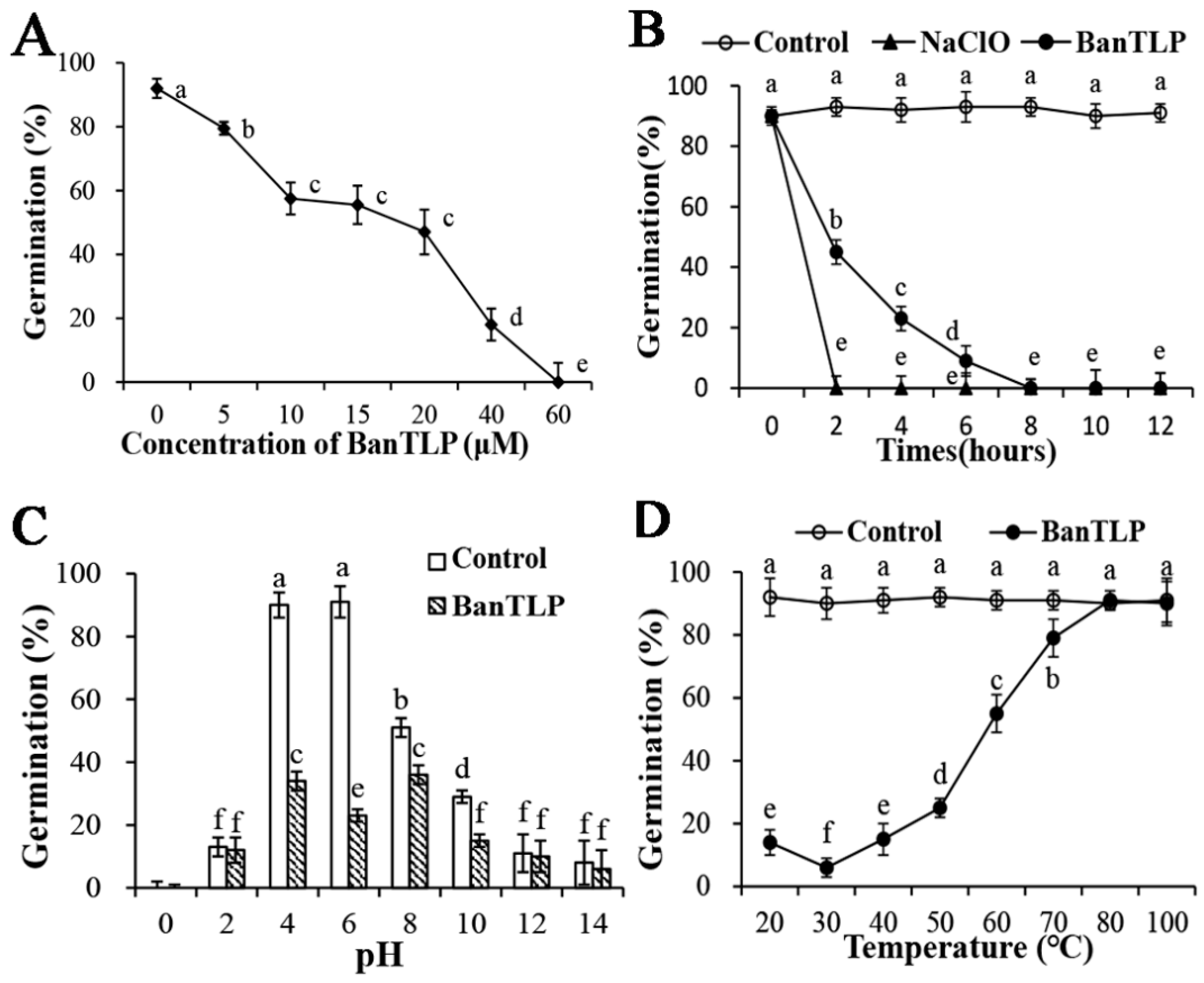
2.2. Antifungal Properties of BanTLP against P. expansum
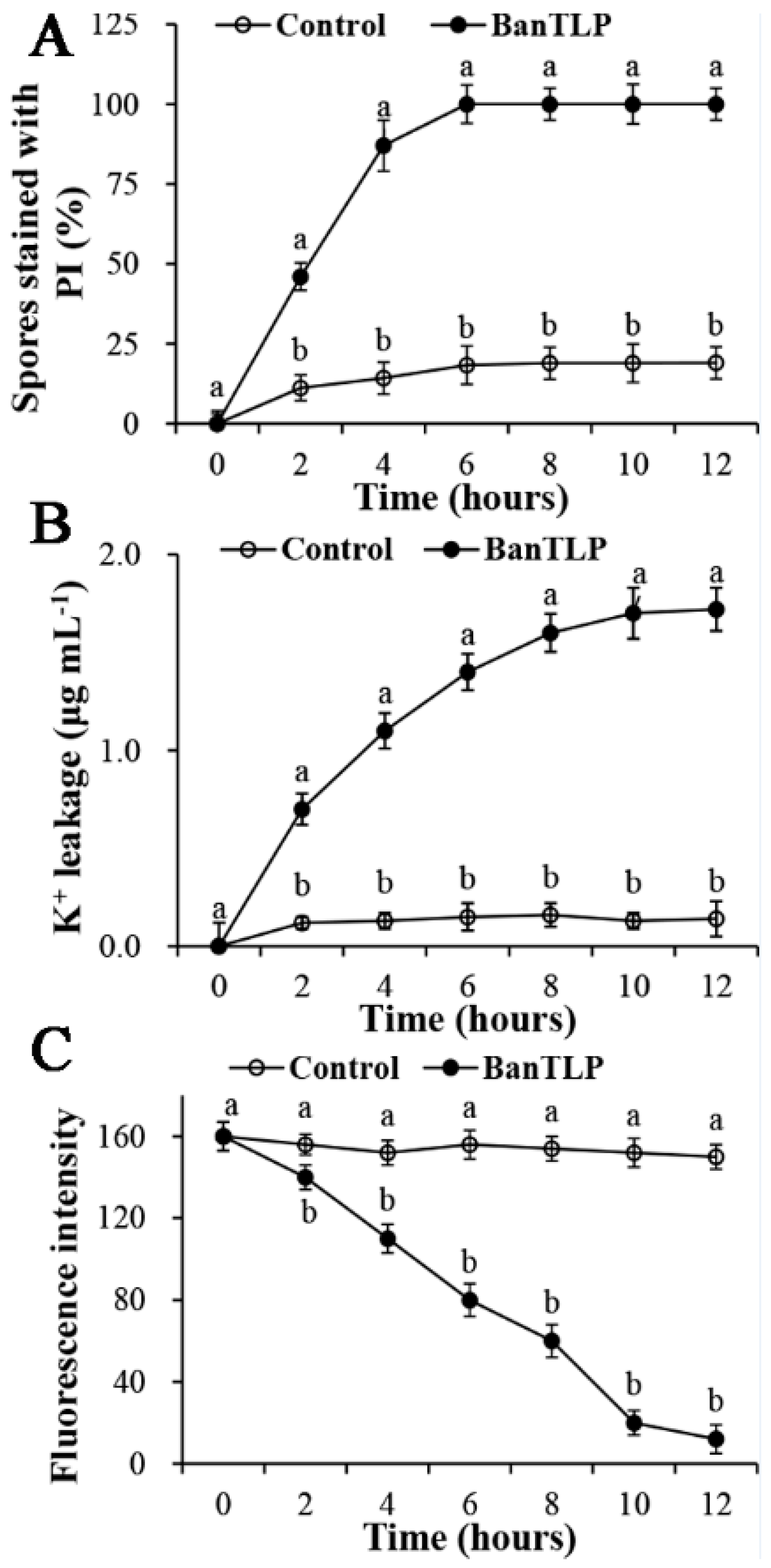
2.3. Effect of BanTLP on Membrane Permeability and Disturbance in P. expansum
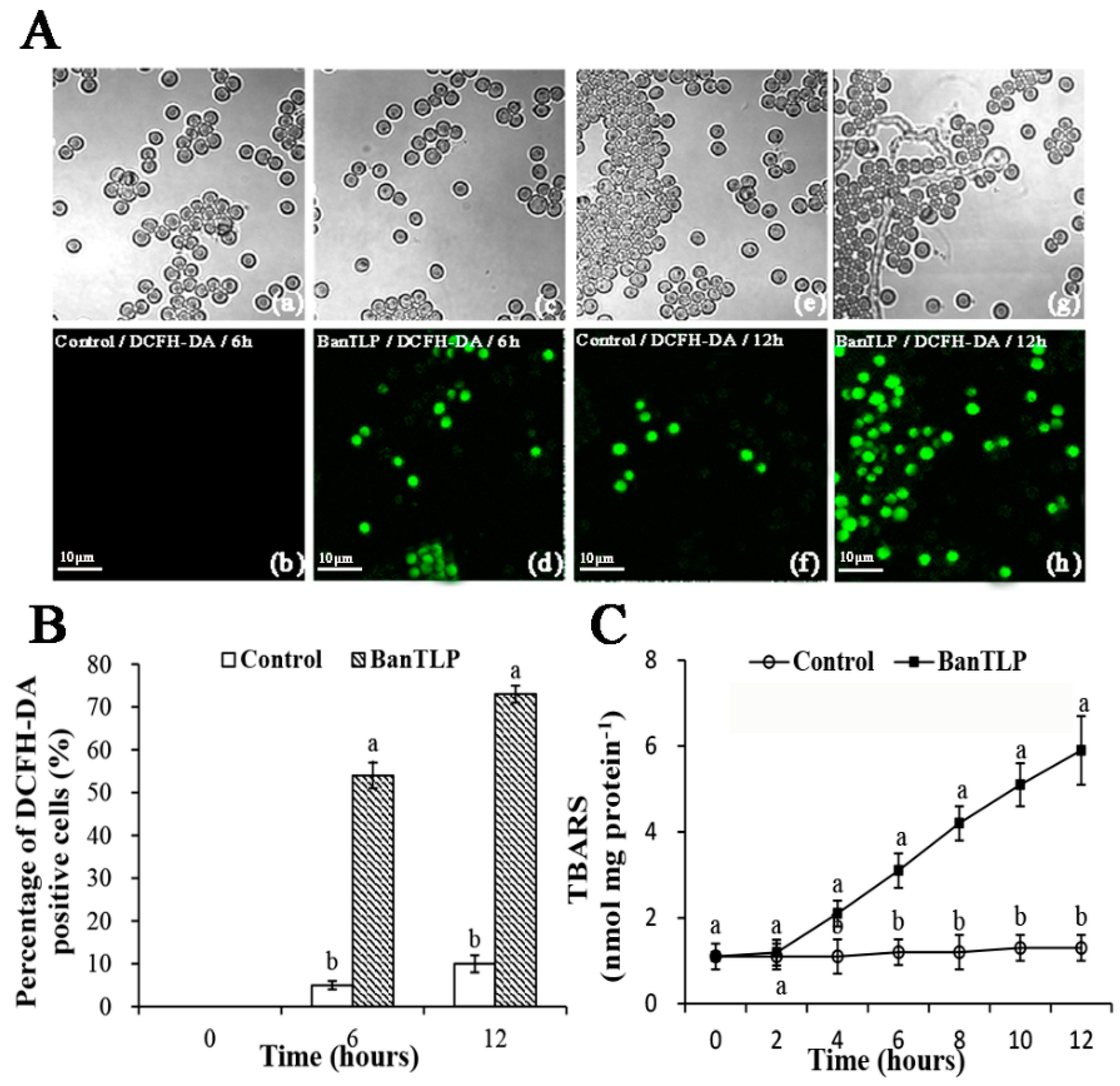
2.4. Oxidative Stress in P. expansum Induced by BanTLP
2.5. Effect of BanTLP on Cell Wall Integrity in P. expansum
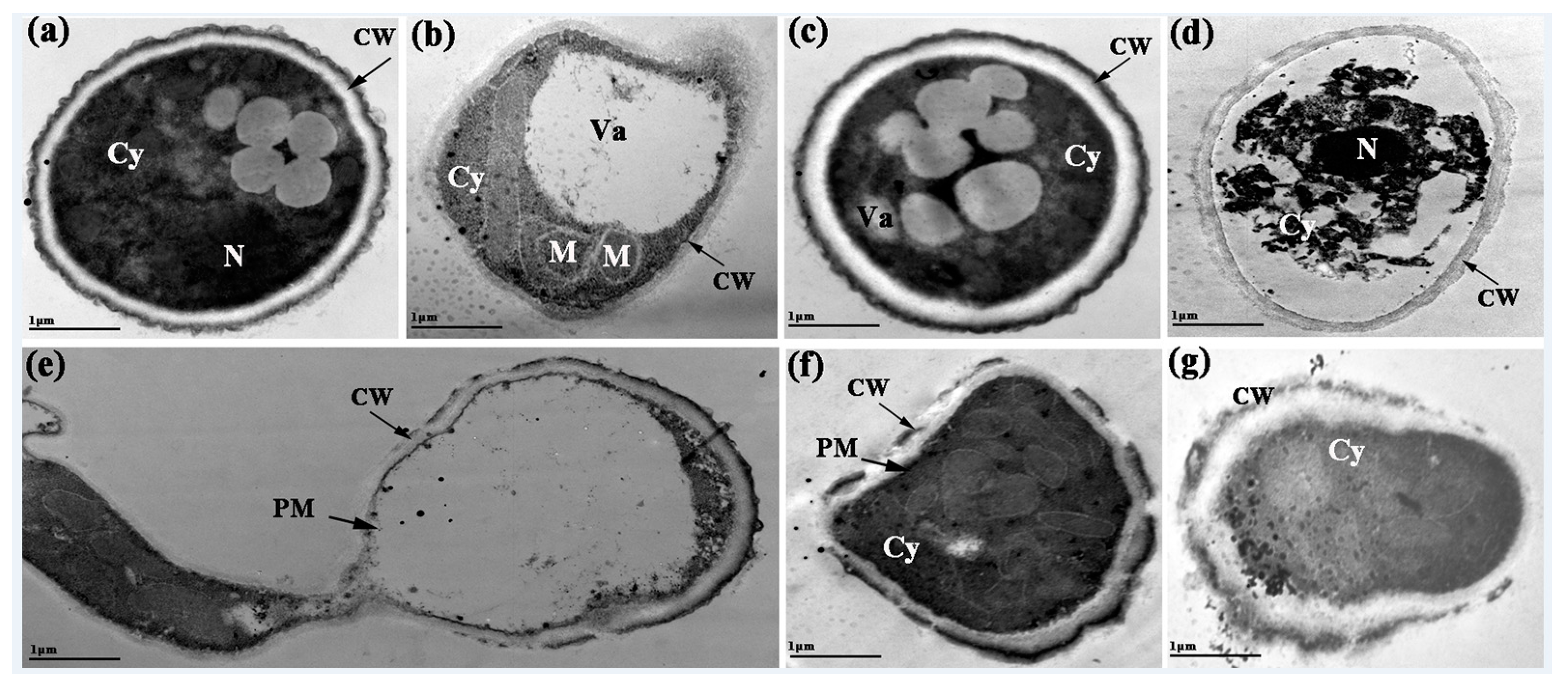
2.6. Morphological and Ultrastructural Changes of P. expansum Conidia
2.6.1. Scanning Electron Microscope (SEM)
2.6.2. Transmission Electron Microscope (TEM)
3. Materials and Methods
3.1. Fruit and Fungal Materials
3.2. Purification and Identification of BanTLP from Banana
3.3. Effect of BanTLP on Spore Germination of P. expansum
3.4. Effect of BanTLP on the Plasma Membrane of P. expansum
3.4.1. PI Influx
3.4.2. Potassium Release Assay
3.4.3. Fluorescence Intensities Assay Related to the Membrane Disturbance
3.4.4. ROS Determination
3.4.5. Determination of TBARS
3.5. Effect of BanTLP on Cell Wall Integrity of P. expansum
3.6. Morphological and Ultrastructural Changes of P. expansum Conidia
3.6.1. SEM
3.6.2. TEM
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Tsamo, C.V.P.; Herent, M.F.; Tomekpe, K.; Emaga, T.H.; Quetin-Leclercq, J.; Rogez, H.; Larondelle, Y.; Andre, C. Phenolic profiling in the pulp and peel of nine plantain cultivars (Musa sp.). Food Chem. 2015, 167, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Barre, A.; Peumans, W.J.; Menu-Bouaouiche, L.; Damme, E.J.; May, G.D.; Herrera, A.F.; Leuven, F.V.; Rougé, P. Purification and structural analysis of an abundant thaumatin-like protein from ripe banana fruit. Planta 2000, 211, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Sturrock, R.; Ekramoddoullah, A.K.M. The superfamily of thaumatin-like proteins: Its origin, evolution, and expression towards biological function. Plant Cell Rep. 2010, 29, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Menu, L. A molecular basis for the endo-β1, 3-glucanase activity of the thaumatin-like proteins from edible fruits. Biochimie 2003, 85, 123–131. [Google Scholar] [CrossRef]
- Grenier, J.; Potvin, C.; Asselin, A. Some Fungi Express β-1,3-Glucanases Similar to Thaumatin-like Proteins. Mycologia 2000, 92, 841–848. [Google Scholar] [CrossRef]
- Tian, B.; Harrison, R.; Morton, J.; Jaspers, M.; Hodge, S.; Grose, C.; Trought, M. Extraction of Pathogenesis-Related Proteins and Phenolics in Sauvignon Blanc as Affected by Grape Harvesting and Processing Conditions. Molecules 2017, 22, 1164. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, N.; Saleem, M. Biochemical characterization of fruit-specific pathogenesis-related antifungal protein from basrai banana. Microbiol. Res. 2014, 169, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Palacín, A.; Tordesillas, L.; Gamboa, P.; Sanchez-Monge, R.; Cuesta-Herranz, J.; Sanz, M.L.; Barber, D.; Salcedo, G.; Díaz-Perales, A. Characterization of peach thaumatin-like proteins and their identification as major peach allergens. Clin. Exp. Allergy 2010, 40, 1422–1430. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ng, T.B. Isolation of an antifungal thaumatin-like protein from kiwi fruits. Phytochemistry 2002, 61, 1–6. [Google Scholar] [CrossRef]
- Koiwa, H.; Kato, H.; Nakatsu, T.; Oda, J.; Yamada, Y.; Sato, F. Crystal structure of tobacco PR-5d protein at 1.8 A resolution reveals a conserved acidic cleft structure in antifungal thaumatin-like proteins. J. Mol. Biol. 1999, 286, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cheng, F.; Sun, Y.; Ma, H.; Yang, X. Structure–Function Relationship of a Novel PR-5 Protein with Antimicrobial Activity from Soy Hulls. J. Agric. Food Chem. 2016, 64, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, L.; Viã As, I.; Torres, R.; Usall, J.; Jauset, A.M.; Teixidã, N. Infection capacities in the orange-pathogen relationship: Compatible (Penicillium digitatum) and incompatible (Penicillium expansum) interactions. Food Microbiol. 2012, 29, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, H.P.; Madsen, L.S.; Petersen, J.; Andersen, J.T.; Hansen, A.M.; Beck, H.C. Oat (Avena sativa) seed extract as an antifungal food preservative through the catalytic activity of a highly abundant class I chitinase. Appl. Biochem. Biotechnol. 2010, 160, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Chakrabarti, C. Crystal structure analysis of NP24-I: A thaumatin-like protein. Planta 2008, 228, 883. [Google Scholar] [CrossRef] [PubMed]
- Fierens, E.; Gebruers, K.; Voet, A.R.; De Maeyer, M.; Courtin, C.M.; Delcour, J.A. Biochemical and structural characterization of TLXI, the Triticum aestivum L. thaumatin-like xylanase inhibitor. J. Enzyme Inhib. Med. Chem. 2009, 24, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lai, T.; Qin, G.; Tian, S. Ambient pH Stress Inhibits Spore Germination of Penicillium expansum by Impairing Protein Synthesis and Folding: A Proteomic-Based Study. J. Proteome Res. 2010, 9, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Ho, V.; Wong, J.T. A thaumatin-like antifungal protein from the emperor banana. Peptides 2007, 28, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, H.; Qin, G.; Meng, X. Chitosan disrupts Penicillium expansum and controls postharvest blue mold of jujube fruit. Food Control 2014, 41, 56–62. [Google Scholar] [CrossRef]
- Yu, S.P. Regulation and critical role of potassium homeostasis in apoptosis. Prog. Neurobiol. 2003, 70, 363–386. [Google Scholar] [CrossRef]
- Repáková, J.; Holopainen, J.M.; Morrow, M.R.; Mcdonald, M.C.; Čapková, P.; Vattulainen, I. Influence of DPH on the Structure and Dynamics of a DPPC Bilayer. Biophys. J. 2005, 88, 3398–3410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Wen, X.; Xie, Y.; Guo, Z.; Zhao, R.; Yu, P.; Gong, D.; Deng, S.; Zeng, Z. Antifungal activity and mechanism of monocaprin against food spoilage fungi. Food Control 2017, 84, 561–568. [Google Scholar] [CrossRef]
- Zheng, S.; Jing, G.; Xiao, W.; Ouyang, Q.; Lei, J.; Tao, N. Citral exerts its antifungal activity against Penicillium digitatum by affecting the mitochondrial morphology and function. Food Chem. 2015, 178, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Duprécrochet, S.; Erard, M.; Nüβe, O. ROS production in phagocytes: Why, when, and where? J. Leukoc. Biol. 2013, 94, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, M.L.; Coca, M.A.; Jin, J.; Yamauchi, T.; Ito, Y.; Kadowaki, T.; Kim, K.K.; Pardo, J.M.; Damsz, B.; Hasegawa, P.M. Osmotin is a homolog of mammalian adiponectin and controls apoptosis in yeast through a homolog of mammalian adiponectin receptor. Mol. Cell 2005, 17, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.M.E.; Bleackley, M.R.; Wiltshire, J.L.; Anderson, M.A.; Traven, A.; van der Weerden, N.L. Identification and Mechanism of Action of the Plant Defensin NaD1 as a New Member of the Antifungal Drug Arsenal against Candida albicans. Antimicrob. Agents Chemother. 2013, 57, 3667–3675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puig, M.; Moragrega, C.; Ruz, L.; Calderã, C.E.; Cazorla, F.M.; Montesinos, E.; Llorente, I. Interaction of antifungal peptide BP15 with Stemphylium vesicarium, the causal agent of brown spot of pear. Fungal Biol. 2016, 120, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Osmond, R.I.W.; Hrmova, M.; Fontaine, F.; Imberty, A.; Fincher, G.B. Binding interactions between barley thaumatin-like proteins and (1,3)-β-d-glucans. FEBS J. 2001, 268, 4190–4199. [Google Scholar] [CrossRef]
- Xi, Y.; Jiao, W.; Cao, J.; Jiang, W. Effects of chlorogenic acid on capacity of free radicals scavenging and proteomic changes in postharvest fruit of nectarine. PLoS ONE 2017, 12, e0182494. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yang, X.; Kang, M.C.; Li, Y.; Kim, E.A.; Kang, S.M.; Jeon, Y.J. Asperflavin, an Anti-Inflammatory Compound Produced by a Marine-Derived Fungus, Eurotium amstelodami. Molecules 2017, 22, 1823. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Nesterenko, M.V.; Tilley, M.; Upton, S.J. A simple modification of Blum’s silver stain method allows for 30 minutes detection of proteins in polyacrylamide gels. J. Biochem. Biophys. Meth. 1994, 28, 239–242. [Google Scholar] [CrossRef]
- Sharma, N.; Tripathi, A. Effects of Citrus sinensis (L.) Osbeck epicarp essential oil on growth and morphogenesis of Aspergillus niger (L.) Van Tieghem. Microbiol. Res. 2008, 163, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha Neto, A.C.; Maraschin, M.; Di Piero, R.M. Antifungal activity of salicylic acid against Penicillium expansum and its possible mechanisms of action. Int. J. Food Microbiol. 2015, 215, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Theis, T.; Marx, F.; Salvenmoser, W.; Stahl, U.; Meyer, V. New insights into the target site and mode of action of the antifungal protein of Aspergillus giganteus. Res. Microbiol. 2005, 156, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Li, B.Q.; Tian, S.P. Effects of trehalose on stress tolerance and biocontrol efficacy of Cryptococcus laurentii. J. Appl. Microbiol. 2006, 100, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Tao, N.; Ouyang, Q.; Jia, L. Citral inhibits mycelial growth of Penicillium italicum by a membrane damage mechanism. Food Control 2014, 41, 116–121. [Google Scholar] [CrossRef]
- Yun, J.; Lee, H.; Ko, H.J.; Woo, E.R.; Lee, D.G. Fungicidal effect of isoquercitrin via inducing membrane disturbance. Biochim. Biophys. Acta 2015, 1848, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Cerioni, L.; Volentini, S.I.; Prado, F.E.; Rapisarda, V.A.; Rodríguez-Montelongo, L. Cellular damage induced by a sequential oxidative treatment on Penicillium digitatum. J. Appl. Microbiol. 2010, 109, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Deng, L.; Yao, S.; Zeng, K. Control of green and blue mold and sour rot in citrus fruits by the cationic antimicrobial peptide PAF56. Postharvest Biol. Technol. 2018, 136, 132–138. [Google Scholar] [CrossRef]
- Li, J.; Bao, X.; Xu, Y.; Zhang, M.; Cai, Q.; Li, L.; Wang, Y. Hypobaric storage reduced core browning of Yali pear fruits. Sci. Hortic. 2017, 225, 547–552. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Total Protein from 1.5 kg Flesh (mg) |
|---|---|
| Crude extract | 1038 |
| Unbound fraction from DEAE–Sephadex A50 | 648 |
| Major peak from Sephadex G50 chromatography | 261 |
| After reversed-phase chromatography | 58 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, W.; Li, X.; Zhao, H.; Cao, J.; Jiang, W. Antifungal Activity of an Abundant Thaumatin-Like Protein from Banana against Penicillium expansum, and Its Possible Mechanisms of Action. Molecules 2018, 23, 1442. https://doi.org/10.3390/molecules23061442
Jiao W, Li X, Zhao H, Cao J, Jiang W. Antifungal Activity of an Abundant Thaumatin-Like Protein from Banana against Penicillium expansum, and Its Possible Mechanisms of Action. Molecules. 2018; 23(6):1442. https://doi.org/10.3390/molecules23061442
Chicago/Turabian StyleJiao, Wenxiao, Xiangxin Li, Handong Zhao, Jiankang Cao, and Weibo Jiang. 2018. "Antifungal Activity of an Abundant Thaumatin-Like Protein from Banana against Penicillium expansum, and Its Possible Mechanisms of Action" Molecules 23, no. 6: 1442. https://doi.org/10.3390/molecules23061442