Determination of the Phenolic Profile and Antioxidant Properties of Salvia viridis L. Shoots: A Comparison of Aqueous and Hydroethanolic Extracts


Abstract
:
1. Introduction
2. Results and Discussion
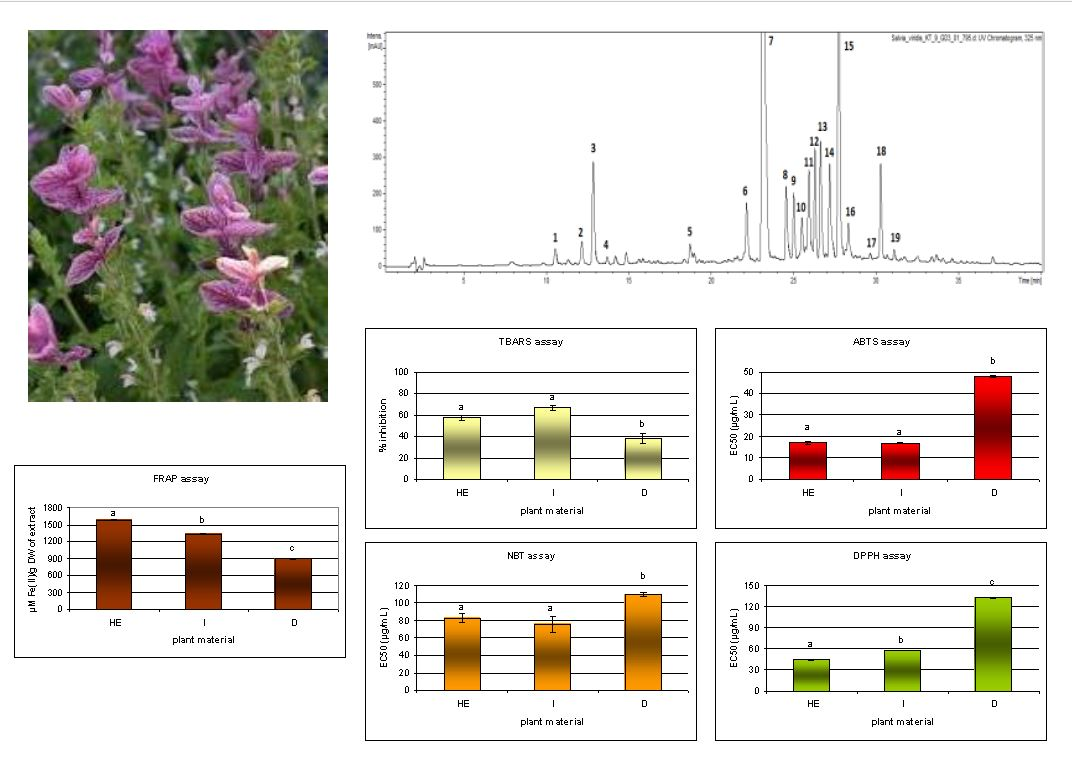
2.1. Identification and Characterization of Bioactive Compounds
2.1.1. Phenolic Acid Derivatives
2.1.2. Flavonoids
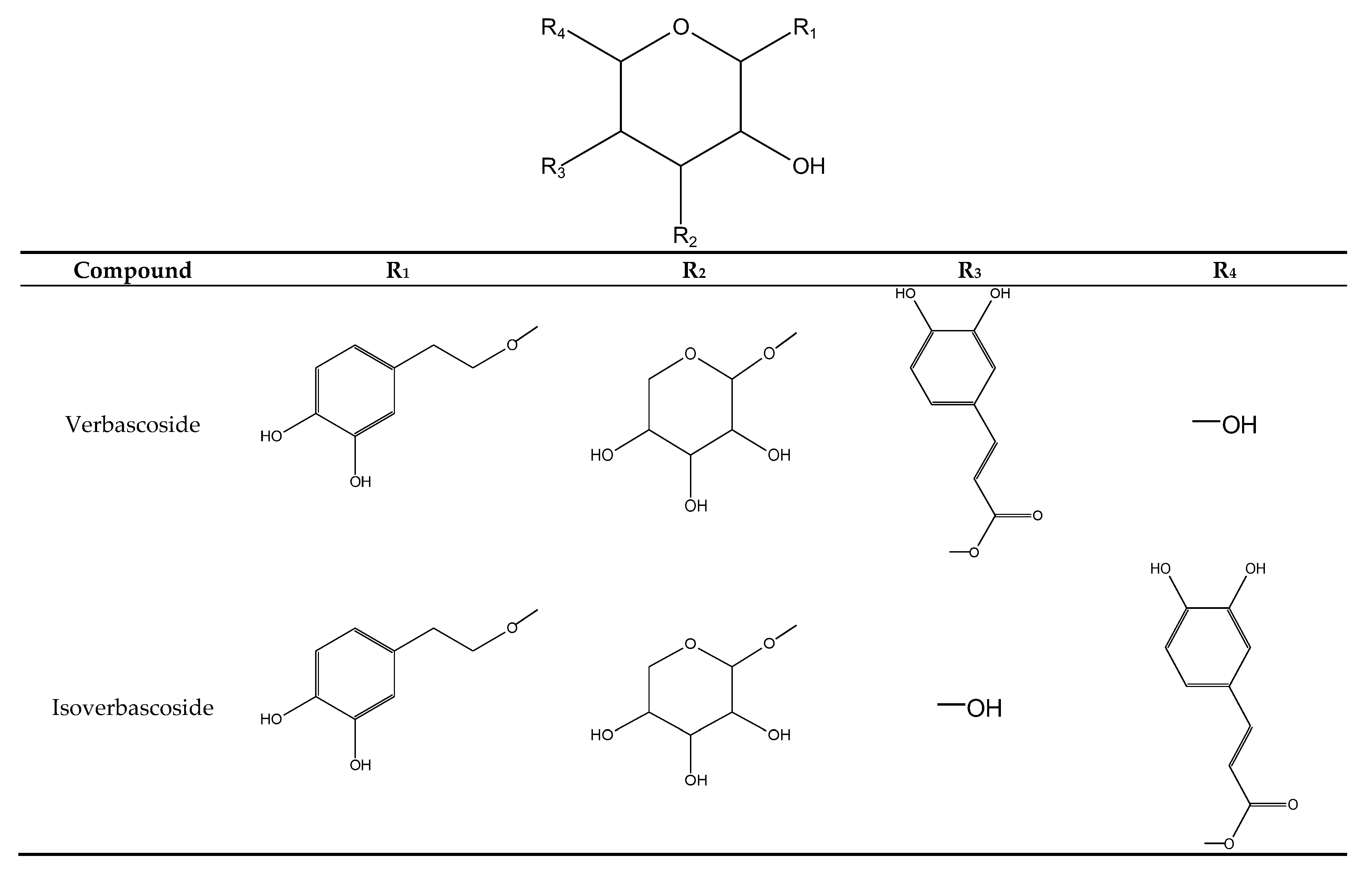
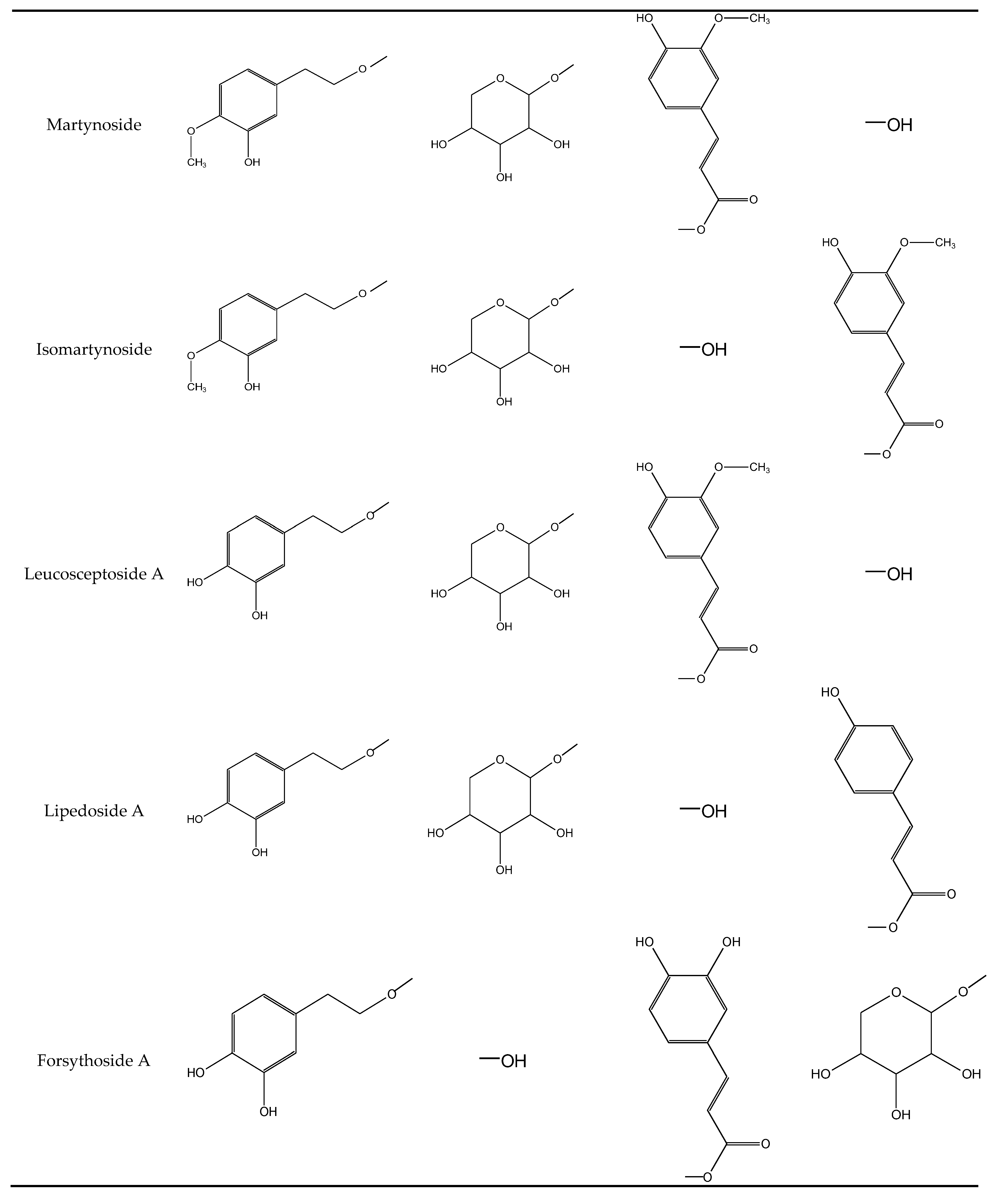
2.1.3. Phenylethanoids
2.2. Quantitative Analysis
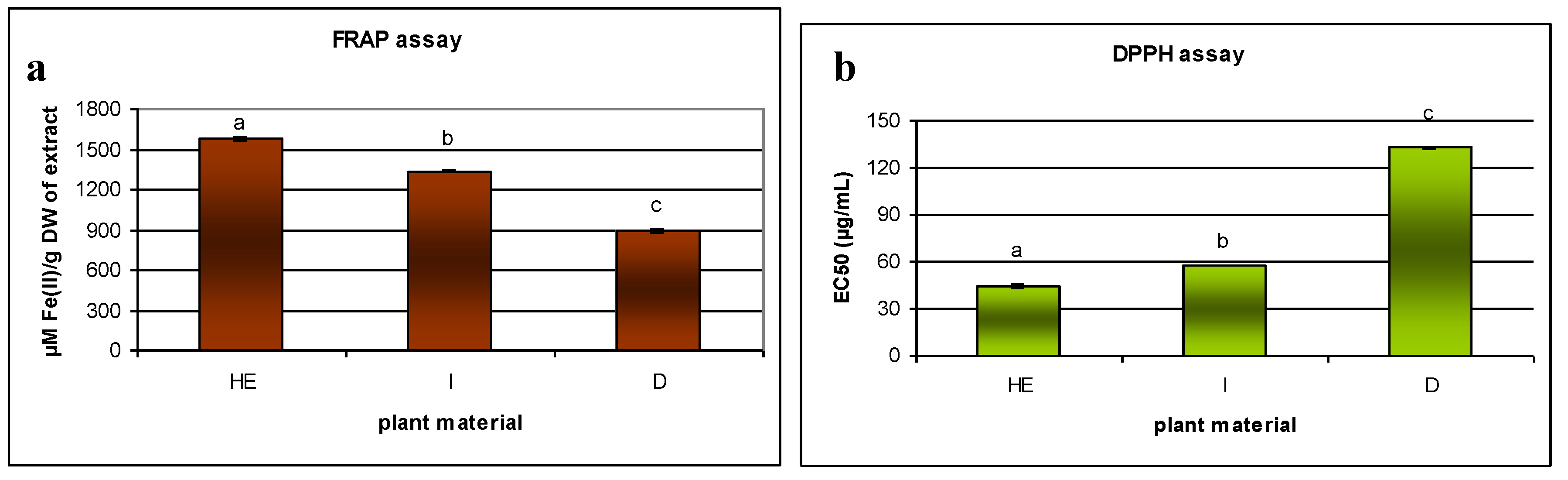
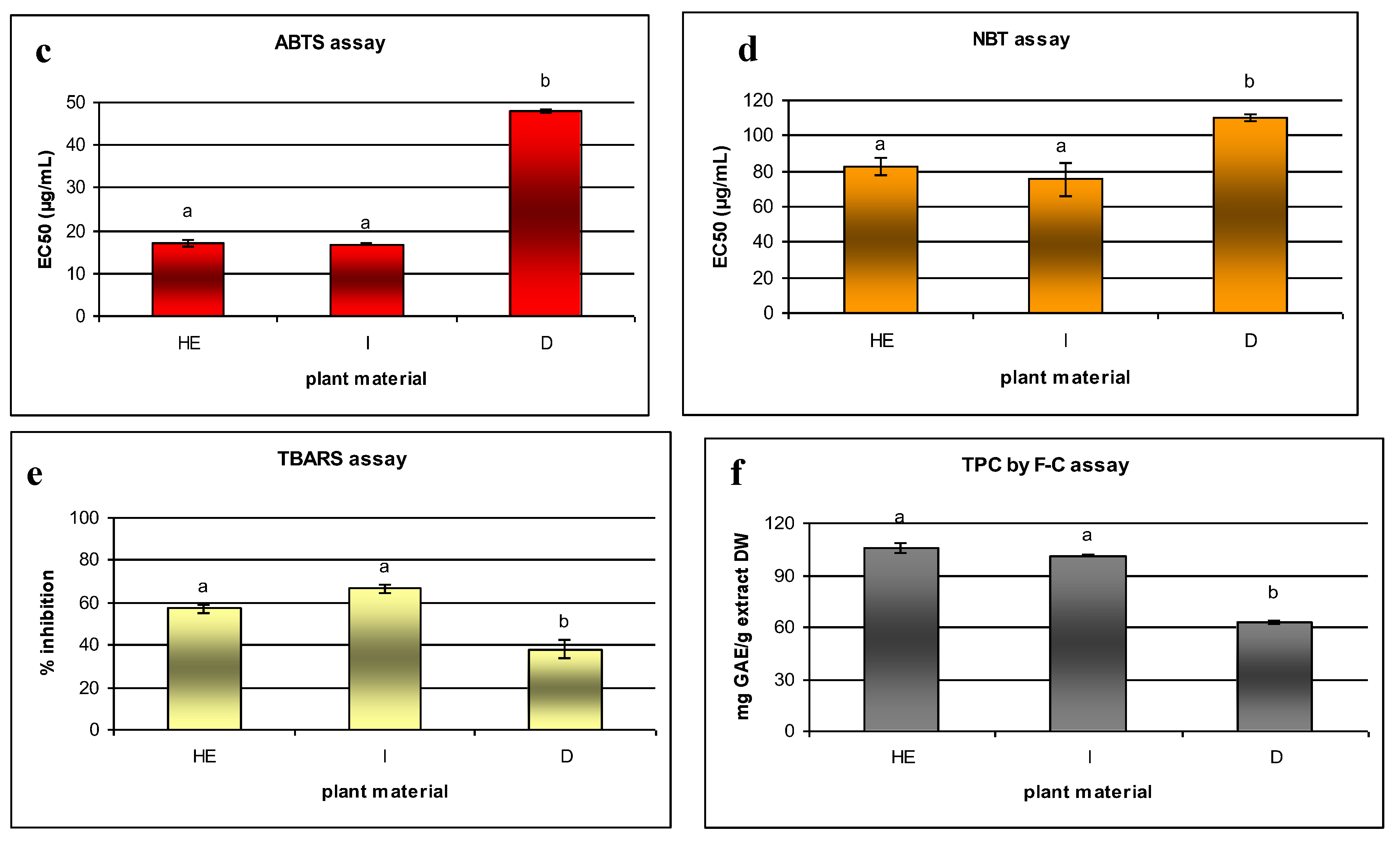
2.3. Antioxidant Properties
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material
3.3. Preparation of Extracts
- -
- Aqueous infusion: Lyophilized and powdered plant material (100 mg) was infused into 30 mL of boiled solvent for 15 min. This process was replicated twice more with 10 mL of the same solvent for 15 min. After filtration, the extracts were combined and evaporated to dryness under reduced pressure.
- -
- Decoction was prepared by boiling of the lyophilized and powdered plant material (100 mg) with the water for 15 min. The extraction process was then replicated twice more with 10 mL of the same solvent for 15 min. After filtration, the extracts were combined and evaporated to dryness under reduced pressure.All obtained extracts were stored as dry extracts at 4 °C until analysis.
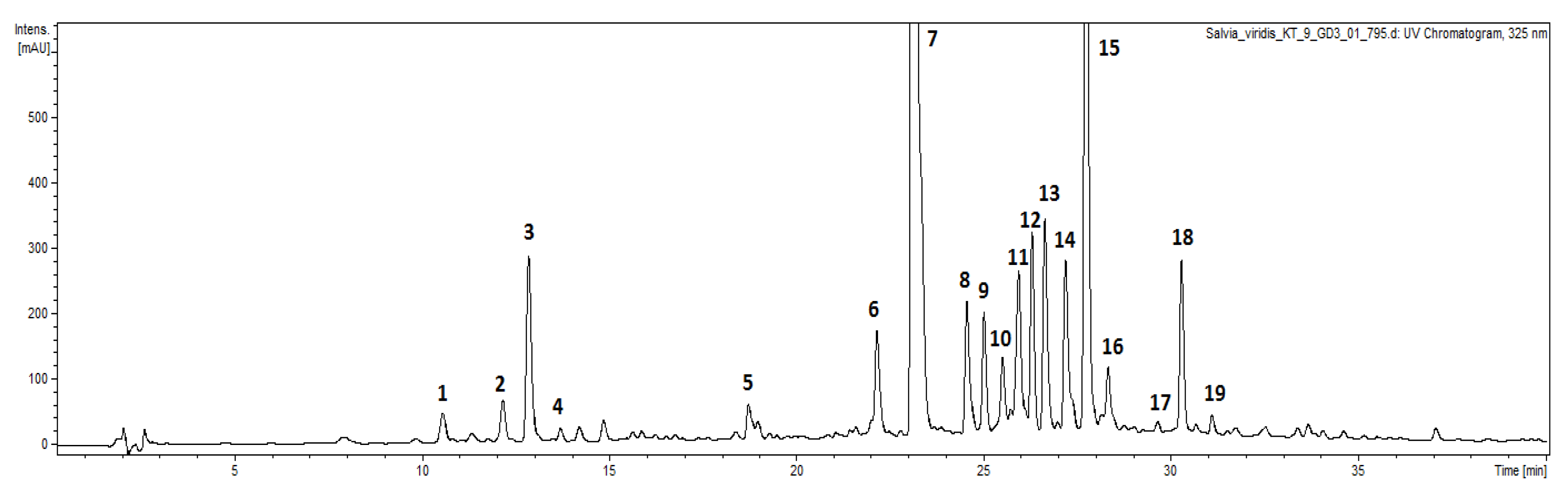
3.4. Qualitative UPLC-DAD-ESI-MS Analysis
3.5. Qualitative Analysis
3.5.1. Total Phenolic Content (TPC)
3.5.2. UPLC Analysis
3.6. Antioxidant Activity
3.6.1. Ferric Reducing Antioxidant Power (FRAP) Assay
3.6.2. DPPH Assay
3.6.3. ABTS Assay
3.6.4. Scavenging of Superoxide Anion Radical (O2•−)—NBT Assay
3.6.5. TBARS Assay
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dweck, A.C. The folklore and cosmetic use of various Salvia species. In Sage Genus Salvia; Taylor & Francis Group: London, UK, 2000; Volume 14, pp. 1–25. [Google Scholar]
- Ghorbani, A. Studies on pharmaceutical ethnobotany in the region of Turkmen Sahra, north of Iran: (Part 1): General results. J. Ethnopharmacol. 2005, 102, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Mosaddegh, M.; Naghibi, F.; Moazzeni, H.; Pirani, A.; Esmaeili, S. Ethnobotanical survey of herbal remedies traditionally used in Kohghiluyeh va Boyer Ahmad province of Iran. J. Ethnopharmacol. 2012, 141, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Ulubelen, A.; Brieskorn, C.H. Micromeric acid from Salvia horminum. Phytochemistry 1975, 14, 1450. [Google Scholar] [CrossRef]
- Ulubelen, A.; Brieskorn, C.H.; Oezdemir, N. Triterpenoids of Salvia horminum, constitution of a new diol. Phytochemistry 1977, 16, 790–791. [Google Scholar] [CrossRef]
- Kokkalou, E.; Kapetanidis, I. Flavonoids and phenolic acids in Salvia horminum L. (Lamiaceae). Pharm. Acta Helv. 1988, 63, 90. [Google Scholar] [PubMed]
- Rungsimakan, S.; Rowan, M.G. Terpenoids, flavonoids and caffeic acid derivatives from Salvia viridis L. cvar. Blue Jeans. Phytochemistry 2014, 108, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Cvetkovikj, I.; Stefkov, G.; Acevska, J.; Stanoeva, J.P.; Karapandzova, M.; Stefova, M.; Dimitrovska, A.; Kulevanova, S. Polyphenolic characterization and chromatographic methods for fast assessment of culinary Salvia species from South East Europe. J. Chromatogr. A 2013, 1282, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Stanojević, L.; Stanković, M.; Nikolić, V.; Nikolić, L.; Ristić, D.; Čanadanovic-Brunet, J.; Tumbas, V. Antioxidant activity and total phenolic and flavonoid contents of Hieracium pilosella L. extracts. Sensors 2009, 9, 5702–5714. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Lee, S.; Elam, M.L.; Johnson, S.A.; Kang, J.; Arjmandi, B.H. Study to find the best extraction solvent for use with guava leaves (Psidium guajava L.) for high antioxidant efficacy. Food Sci. Nutr. 2014, 2, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, Q.; Wang, X.; Yang, J.; Wang, Q. Qualitative analysis and simultaneous quantification of phenolic compounds in the aerial parts of Salvia miltiorrhiza by HPLC-DAD and ESI/MSn. Phytochem. Anal. 2011, 22, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MS n identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Kikuzaki, H.; Zhu, N.; Sang, S.; Nakatani, N.; Ho, C.T. Isolation and structural elucidation of two new glycosides from sage (Salvia officinalis L.). J. Agric. Food Chem. 2000, 48, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.; Chen, J.M.; Peng, Y.; Wu, Q.; Xiao, P.G. Investigation of Danshen and related medicinal plants in China. J. Ethnopharmacol. 2008, 120, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Milevskaya, V.V.; Temerdashev, Z.A.; Butylskaya, T.S.; Kiseleva, N.V. Determination of phenolic compounds in medicinal plants from the Lamiaceae family. J. Anal. Chem. 2017, 72, 342–348. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Kuźma, Ł.; Skała, E.; Kiss, A.K. Hairy root cultures of Salvia viridis L. for production of polyphenolic compounds. Ind. Crops Prod. 2018, 117, 235–244. [Google Scholar] [CrossRef]
- Mitreski, I.; Stanoeva, J.P.; Stefova, M.; Stefkov, G.; Kulevanova, S. Polyphenols in representative Teucrium species in the flora of R. Macedonia: LC/DAD/ESI-MS(n) profile and content. Nat. Prod. Commun. 2014, 9, 175–180. [Google Scholar] [PubMed]
- Pereira, O.R.; Peres, A.M.; Silva, A.M.; Domingues, M.R.; Cardoso, S.M. Simultaneous characterization and quantification of phenolic compounds in Thymus x citriodorus using a validated HPLC–UV and ESI–MS combined method. Food Res. Int. 2013, 54, 1773–1780. [Google Scholar] [CrossRef]
- Beninger, C.W.; Hall, J.C. Allelopathic activity of luteolin 7-O-β-glucuronide isolated from Chrysanthemum morifolium L. Biochem. Syst. Ecol. 2005, 33, 103–111. [Google Scholar] [CrossRef]
- Lech, K.; Witkoś, K.; Jarosz, M. HPLC–UV–ESI MS/MS identification of the color constituents of sawwort (Serratula tinctoria L.). Anal. Bioanal. Chem. 2014, 406, 3703–3708. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.F.; Saleh, N.A.; Gabr, S.; Abu-Eyta, A.M.; El-Said, H. Flavone glycosides of Salvia triloba. Phytochemistry 1983, 22, 2057–2060. [Google Scholar] [CrossRef]
- Bisio, A.; Romussi, G.; Ciarallo, G.; De Tommasi, N. Flavonoide und triterpenoide aus Salvia blepharophylla Brandegee ex Epling. Pharmazie 1997, 52, 330–331. [Google Scholar]
- Ibrahim, R.M.; El-Halawany, A.M.; Saleh, D.O.; El Naggar, E.M.B.; El-Shabrawy, A.E.R.O.; El-Hawary, S.S. HPLC-DAD-MS/MS profiling of phenolics from Securigera securidaca flowers and its anti-hyperglycemic and anti-hyperlipidemic activities. Rev. Bras. Farmacogn. 2015, 25, 134–141. [Google Scholar] [CrossRef]
- Li, L.; Tsao, R.; Liu, Z.; Liu, S.; Yang, R.; Young, J.C.; Fu, Z. Isolation and purification of acteoside and isoacteoside from Plantago psyllium L. by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1063, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Ye, M.; Guo, H.; Yang, M.; Wang, B.R.; Guo, D.A. Analysis of multiple constituents in a Chinese herbal preparation Shuang-Huang-Lian oral liquid by HPLC-DAD-ESI-MSn. J. Pharm. Biomed. 2007, 44, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Göger, F.; Köse, Y.B.; Göger, G.; Demirci, F. Phytochemical characterization of phenolics by LC-MS/MS and biological evaluation of Ajuga orientalis from Turkey. Bangladesh J. Pharmacol. 2015, 10, 639–644. [Google Scholar] [CrossRef]
- Guo, H.; Liu, A.H.; Ye, M.; Yang, M.; Guo, D.A. Characterization of phenolic compounds in the fruits of Forsythia suspensa by high-performance liquid chromatography coupled with electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Kırmızıbekmez, H.; Montoro, P.; Piacente, S.; Pizza, C.; Dönmez, A.; Çalış, İ. Identification by HPLC-PAD-MS and quantification by HPLC-PAD of phenylethanoid glycosides of five Phlomis species. Phytochem. Anal. 2005, 16, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hady, H.; El-Sayed, M.M.; Abdel-Hady, A.A.; Hashash, M.M.; Abdel-Hady, A.M.; Aboushousha, T.; Abdel-Hameed, E.S.; Abdel-Lateef, E.E.; Morsi, E.A. Nephroprotective Activity of methanolic extract of Lantana camara and squash (Cucurbita pepo) on cisplatin-induced nephrotoxicity in rats and identification of certain chemical constituents of Lantana camara by HPLC-ESI-MS. Pharmacogn. J. 2018, 10, 136–147. [Google Scholar] [CrossRef]
- Wu, S.J.; Chan, Y.Y. Five new iridoids from roots of Salvia digitaloides. Molecules 2014, 19, 155521–155534. [Google Scholar] [CrossRef] [PubMed]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Nazemiyeh, H.; Rahman, M.M.; Gibbons, S.; Nahar, L.; Delazar, A.; Ghahramani, M.A.; Sarker, S.D. Assessment of the antibacterial activity of phenylethanoid glycosides from Phlomis lanceolata against multiple-drug-resistant strains of Staphylococcus aureus. J. Nat. Med. 2008, 62, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhao, Y.; Norman, V.L.; Starks, C.M.; Rice, S.M.; Goering, M.G.; O’Neil-Johnson, M.; Eldridge, G.R.; Hu, J.F. Antibiofilm phenylethanoid glycosides from Penstemon centranthifolius. Phytother. Res. 2010, 24, 778–781. [Google Scholar] [PubMed]
- Lee, K.Y.; Jeong, E.J.; Lee, H.S.; Kim, Y.C. Acteoside of Callicarpa dichotoma attenuates scopolamine-induced memory impairments. Biol. Pharm. Bull. 2006, 29, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.M.; Gao, L.; Huo, S.X.; Liu, X.M.; Yan, M. The mechanism of memory enhancement of acteoside (verbascoside) in the senescent mouse model induced by a combination of d-gal and AlCl3. Phytother. Res. 2015, 29, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Kurisu, M.; Miyamae, Y.; Murakami, K.; Han, J.; Isoda, H.; Irie, K.; Shigemori, H. Inhibition of amyloid β aggregation by acteoside, a phenylethanoid glycoside. Biosci. Biotechnol. Biochem. 2013, 77, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Jia, Z.; Deng, Z.; Wei, Y.; Zheng, R.; Yu, L. In vitro modulation of telomerase activity, telomere length and cell cycle in MKN45 cells by verbascoside. Planta Med. 2002, 68, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Yang, B. Phenylethanoid glycosides: Research advances in their phytochemistry, pharmacological activity and pharmacokinetics. Molecules 2016, 21, 991. [Google Scholar] [CrossRef] [PubMed]
- Tóth, E.; Tóth, G.; Máthé, I.; Blunden, G. Martynoside, forsythoside B, ladanein and 7α-acetoxyroyleanone from Ballota nigra L. Biochem. Syst. Ecol. 2007, 35, 894–897. [Google Scholar] [CrossRef]
- Çaliş, İ.; Hosny, M.; Khalifa, T.; Rüedi, P. Phenylpropanoid glycosides from Marrubium alysson. Phytochemistry 1992, 31, 3624–3626. [Google Scholar] [CrossRef]
- Charami, M.T.; Lazari, D.; Karioti, A.; Skaltsa, H.; Hadjipavlou-Litina, D.; Souleles, C. Antioxidant and antiinflammatory activities of Sideritis perfoliata subsp. perfoliata (Lamiaceae). Phytother. Res. 2008, 22, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, J.; Rédei, D.; Máthé, I.; Blunden, G. Phenylpropanoid glycosides and diterpenoids from Salvia officinalis. Biochem. Syst. Ecol. 2003, 31, 427–429. [Google Scholar] [CrossRef]
- Kasimu, R.; Tanaka, K.; Tezuka, Y.; Gong, Z.N.; Li, J.X.; Basnet, P.; Kadota, S. Comparative study of seventeen Salvia plants: Aldose reductase inhibitory activity of water and MeOH extracts and liquid chromatography-mass spectrometry (LC-MS) analysis of water extracts. Chem. Pharm. Bull. 1998, 46, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Simmonds, M.S. Rosmarinic acid. Phytochemistry 2003, 62, 121–125. [Google Scholar] [CrossRef]
- Moon, D.O.; Kim, M.O.; Lee, J.D.; Choi, Y.H.; Kim, G.Y. Rosmarinic acid sensitizes cell death through suppression of TNF-α-induced NF-κB activation and ROS generation in human leukemia U937 cells. Cancer Lett. 2010, 288, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Tewtrakul, S.; Miyashiro, H.; Nakamura, N.; Hattori, M.; Kawahata, T.; Otake, T.; Rattanasuwon, P. HIV-1 integrase inhibitory substances from Coleus parvifolius. Phytother. Res. 2003, 17, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Ci, X.; He, J.; Jiang, L.; Wei, M.; Cao, Q.; Guan, M.; Xie, X.; Deng, X.; He, J. Effects of a natural prolyl oligopeptidase inhibitor, rosmarinic acid, on lipopolysaccharide-induced acute lung injury in mice. Molecules 2012, 17, 3586–3598. [Google Scholar] [CrossRef] [PubMed]
- Kocak, M.S.; Sarikurkcu, C.; Cengiz, M.; Kocak, S.; Uren, M.C.; Tepe, B. Salvia cadmica: Phenolic composition and biological activity. Ind. Crop. Prod. 2016, 85, 204–212. [Google Scholar] [CrossRef]
- Li, H.; Pordesimo, L.; Weiss, J. High intensity ultrasound assisted extraction of oil from soy beans. Food Res. Int. 2004, 37, 731–738. [Google Scholar] [CrossRef]
- Goltz, C.; Ávila, S.; Barbieri, J.B.; Igarashi-Mafra, L.; Mafra, M.R. Ultrasound-assisted extraction of phenolic compounds from Macela (Achyrolcine satureioides) extracts. Ind. Crop. Prod. 2018, 115, 227–234. [Google Scholar] [CrossRef]
- Han, L.; Boakye-Yiadom, M.; Liu, E.; Zhang, Y.; Li, W.; Song, X.; Fu, F.; Gao, X. Structural characterization and identification of phenylethanoid glycosides from Cistanches deserticola YC Ma by UHPLC/ESI–QTOF–MS/MS. Phytochem. Anal. 2012, 23, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Saimaru, H.; Orihara, Y. Biosynthesis of acteoside in cultured cells of Olea europaea. J. Nat. Med. 2010, 64, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, A.; Pati, S.; Minervini, F.; D’Antuono, I.; Linsalata, V.; Lattanzio, V. Verbascoside, isoverbascoside, and their derivatives recovered from olive mill wastewater as possible food antioxidants. J. Agric. Food Chem. 2012, 60, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Tatli, I.I.; Akdemir, Z.S.; Yesilada, E.; Küpeli, E. Anti-inflammatory and antinociceptive potential of major phenolics from Verbascum salviifolium Boiss. Z. Naturforsch. C 2008, 63, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Nishibe, S.; Sasahara, M.; Ying, J.; Lu, Y.C.; Tanaka, T. Phenylethanoids glycosides from Plantago depressa. Phytochemistry 1993, 32, 975–977. [Google Scholar] [CrossRef]
- Wang, P.; Kang, J.; Zheng, R.; Yang, Z.; Lu, J.; Gao, J.; Jia, Z. Scavenging effects of phenylpropanoid glycosides from Pedicularis on superoxide anion and hydroxyl radical by the spin trapping method(95)02255-4. Biochem. Pharmacol. 1996, 51, 687–691. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Žugić, A.; Đorđević, S.; Arsić, I.; Marković, G.; Živković, J.; Jovanović, S.; Tadić, V. Antioxidant activity and phenolic compounds in 10 selected herbs from Vrujci Spa, Serbia. Ind. Crops Prod. 2014, 52, 519–527. [Google Scholar] [CrossRef]
- Weremczuk-Jeżyna, I.; Grzegorczyk-Karolak, I.; Frydrych, B.; Królicka, A.; Wysokińska, H. Hairy roots of Dracocephalum moldavica: Rosmarinic acid content and antioxidant potential. Acta Physiol. Plant. 2013, 35, 2095–2103. [Google Scholar] [CrossRef]
- Erdemoglu, N.; Turan, N.N.; Cakõcõ, I.; Sener, B.; Aydõn, A. Antioxidant activities of some Lamiaceae plant extracts. Phytother. Res. 2006, 20, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Matkowski, A.; Zielińska, S.; Oszmiański, J.; Lamer-Zarawska, E. Antioxidant activity of extracts from leaves and roots of Salvia miltiorrhiza Bunge, S. przewalskii Maxim., and S. verticillata L. Bioresour. Technol. 2008, 99, 7892–7896. [Google Scholar] [CrossRef] [PubMed]
- Grzegorczyk, I.; Matkowski, A.; Wysokińska, H. Antioxidant activity of extracts from in vitro cultures of Salvia officinalis L. Food Chem. 2007, 104, 536–541. [Google Scholar] [CrossRef]
- Kamatou, G.P.; Viljoen, A.M.; Steenkamp, P. Antioxidant, antiinflammatory activities and HPLC analysis of South African Salvia species. Food Chem. 2010, 119, 684–688. [Google Scholar] [CrossRef]
- Bejeli, M.; Rowshan, V.; Zakerin, A. Comparison of total phenolic content and antioxidant activity of five Salvia species by FRAP and DPPH assay. Int. J. Pharm. Pharm. Sci. 2012, 4, 572–575. [Google Scholar]
- Grąbkowska, R.; Matkowski, A.; Grzegorczyk-Karolak, I.; Wysokińska, H. Callus cultures of Harpagophytum procumbens (Burch.) DC. ex Meisn.; production of secondary metabolites and antioxidant activity. S. Afr. J. Bot. 2016, 103, 41–48. [Google Scholar] [CrossRef]
- Piątczak, E.; Grzegorczyk-Karolak, I.; Wysokińska, H. Micropropagation of Rehmannia glutinosa Libosch.: Production of phenolics and flavonoids and evaluation of antioxidant activity. Acta Physiol. Plant. 2014, 36, 1693–1702. [Google Scholar] [CrossRef]
- Ruffino Maria, M.; Alves, R.E.; de Brito, E.S.; Pérez-Jiménez, J.; Saura-Calixto, F.; Mancini-Filho, J. Bioactive compounds and antioxidant capacities of 18 non-traditional tropical fruits from Brazil. Food Chem. 2010, 121, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Arts, M.J.; Haenen, G.R.; Voss, H.P.; Bast, A. Antioxidant capacity of reaction products limits the applicability of the Trolox Equivalent Antioxidant Capacity (TEAC) assay. Food Chem. Toxicol. 2004, 42, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Chaves López, C.; Mazzarrino, G.; Rodríguez, A.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda Martos, M. Assessment of antioxidant and antibacterial potential of borojo fruit (Borojo apatinoi Cuatre-casas) from the rain forests of South America. Ind. Crops Prod. 2015, 63, 79–86. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Kuźma, Ł.; Wysokińska, H. Study on the chemical composition and antioxidant activity of extracts from shoot culture and regenerated plants of Scutellaria altissima L. Acta Physiol. Plant. 2015, 37, 1736. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Kuźma, Ł.; Wysokińska, H. The effect of cytokinins on shoot proliferation, secondary metabolite production and antioxidant potential in shoot cultures of Scutellaria alpina. Plant Cell Tissue Organ Cult. 2015, 122, 699–708. [Google Scholar] [CrossRef]
- Nishikimi, M.; Rao, N.A.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Marchelak, A.; Owczarek, A.; Matczak, M.; Pawlak, A.; Kolodziejczyk-Czepas, J.; Nowak, P.; Olszewska, M.A. Bioactivity potential of Prunus spinosa L. flower extracts: Phytochemical profiling, cellular safety, pro-inflammatory enzymes inhibition and protective effects against oxidative stress in vitro. Front. Pharmacol. 2017, 8, 680. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Retention Time [min] | UV [nm] | [M − H]− | Fragmentation Ions | |
|---|---|---|---|---|---|
| 1 | 6-O-caffeoylglucose (I) | 10.6 | 324 | 341 | 323, 281, 251, 221, 179 |
| 2 | 6-O-caffeoylglucose (II) | 12.3 | 325 | 341 | 323, 281, 251, 221, 179 |
| 3 | 5-O-caffeoylquinic acid (chlorogenic acid) a | 12.8 | 325 | 353 | 191 |
| 4 | 4-O-caffeoylquinic acid (cryptochlorogenic acid) | 13.7 | 325 | 353 | 191, 173, 135 |
| 5 | caffeoyl-hexoside derivative | 18.7 | 325 | 537 | 519, 341, 281, 179 |
| 6 | luteolin-O-rutinoside | 22.1 | 253, 269, 344 | 593 | 285 |
| 7 | verbascoside a | 23.1 | 330 | 623 | 461, 315, 135 |
| 8 | forsythoside A | 24.5 | 326 | 623 | 461, 477, 315, 135 |
| 9 | isoverbascoside a | 25.0 | 326 | 623 | 461, 315, 135 |
| 10 | lipedoside A | 25.8 | 316 | 607 | 461, 443, 315, 297, 135 |
| 11 | dicaffeoylquinic acid | 26.0 | 328 | 515 | 353, 191, 179 |
| 12 | leucosceptoside A | 26.3 | 328 | 637 | 461, 315, 135 |
| 13 | apigenin-O-hexuronide | 26.7 | 267, 332 | 445 | 269, 149 |
| 14 | methylluteolin-O-hexuronide (chrysoeriol hexuronide) | 27.1 | 269, 343 | 475 | 299, 285, 175 |
| 15 | rosmarinic acid a | 27.7 | 327 | 359 | 197, 179, 161 |
| 16 | unidentified phenylethanoid | 28.4 | 328 | 803 | 641, 461, 443, 315 |
| 17 | luteolin-O-dihexoside | 29.7 | 255, 360 | 609 | 447, 429, 285 |
| 18 | martynoside | 30.3 | 328 | 651 | 505, 475, 457, 329 |
| 19 | isomartynoside | 31.1 | 328 | 651 | 505, 475, 457, 329, 193 |
| Compound | Hydroethanolic Extract | Infusion | Decoction |
|---|---|---|---|
| Phenylethanoids | |||
| verbascoside (A) | 9.60 ± 0.170a | 9.10 ± 0.433a | 0.870 ± 0.109b |
| forsythoside A (A) | 0.493 ± 0.005a | 0.251 ± 0.011b | trace |
| isoverbascoside (B) | 0.474 ± 0.007b | 0.657 ± 0.069a | 0.388 ± 0.059b |
| lipedoside A (A) | 0.218 ± 0.005a | 0.234 ± 0.001a | 0.0883 ± 0.005b |
| leucosceptoside A (A) | 0.524 ± 0.011a | 0.512 ± 0.030a | 0.0136 ± 0.0002b |
| unidentified phenylethanoid (A) | 0.158 ± 0.006b | 0.197 ± 0.007a | 0.0433 ± 0.008c |
| martynoside (A) | 0.332 ± 0.009b | 0.358 ± 0.002a | 0.0445 ± 0.005c |
| isomartynoside (A) | 0.044 ± 0.002a | 0.035 ± 0.005a | trace |
| Total phenylethanoids (TP) | 11.843 ± 0.215a | 11.344 ± 0.558a | 1.448 ± 0.167b |
| Polyphenolic acids | |||
| 6-O-caffeoylglucose (I) (C) | 0.033 ± 0.001a | 0.040 ± 0.0004a | 0.011 ± 0.002b |
| 6-O-caffeoylglucose (II) (C) | 0.066 ± 0.001a | 0.047 ± 0.0002b | 0.016 ± 0.002c |
| 5-O-caffeoylquinic acid (D) | 0.354 ± 0.026a | 0.313 ± 0.006a | 0.054 ± 0.01b |
| 4-O-caffeoylquinic acid (D) | 0.019 ± 0.0009c | 0.026 ± 0.0008b | 0.083 ± 0.003a |
| caffeoyl-hexoside derivative (C) | 0.039 ± 0.0007b | 0.088 ± 0.001a | 0.066 ± 0.017ab |
| dicaffeoylquinic acid (C) | 0.124 ± 0.003a | 0.054 ± 0.001b | 0.051 ± 0.005b |
| rosmarinic acid (E) | 1.267 ± 0.058a | 1.283 ± 0.050a | 0.525 ± 0.145b |
| Total polyphenolic acids (TPA) | 1.902 ± 0.091a | 1.81 ± 0.07a | 0.806 ± 0.18b |
| Flavonoids | |||
| luteolin-O-rutinoside (F) | 1.662 ± 0.029a | 1.630 ± 0.015a | 1.236 ± 0.111b |
| apigenin-O-hexuronide (F) | 4.580 ± 0.111b | 5.287 ± 0.016a | 0.988 ± 0.033c |
| methylluteolin-O-hexuronide (F) | 2.661 ± 0.050b | 5.163 ± 0.655a | 2.406 ± 0.199b |
| luteolin-O-dihexoside (F) | 0.143 ± 0.003a | 0.0659 ± 0.006b | 0.015 ± 0.002c |
| Total flavonoids (TF) | 9.046 ± 0.193b | 12.146 ± 0.692a | 4.299 ± 0.345c |
| Total phenolic compound | 22.791 ± 0.499a | 25.341 ± 1.320a | 6.553 ± 0.696b |
| Assay | Antioxidant Activity Method | ||||
|---|---|---|---|---|---|
| FRAP | DPPH | ABTS | NBT | TBARS | |
| TP | 0.952 | −0.997 | −0.999 | −0.970 | 0.930 |
| TPA | 0.932 | −0.900 | −0.999 | −0.983 | 0.951 |
| TF | 0.731 | −0.866 | −0.922 | −0.981 | 0.998 |
| TPC | 0.886 | −0.968 | −0.992 | −0.997 | 0.979 |
| TPC (by Folin-Ciocalteu) | 0.968 | −0.999 | −0.995 | −0.954 | 0.908 |
| FRAP | - | −0.974 | −0.938 | −0.849 | 0.774 |
| DPPH | −0.974 | - | 0.992 | 0.947 | −0.897 |
| ABTS | −0.938 | 0.992 | - | 0.980 | −0.946 |
| NBT | −0.849 | 0.947 | 0.980 | - | −0.992 |
| TBARS | 0.774 | −0.897 | −0.946 | −0.992 | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzegorczyk-Karolak, I.; Kiss, A.K. Determination of the Phenolic Profile and Antioxidant Properties of Salvia viridis L. Shoots: A Comparison of Aqueous and Hydroethanolic Extracts. Molecules 2018, 23, 1468. https://doi.org/10.3390/molecules23061468
Grzegorczyk-Karolak I, Kiss AK. Determination of the Phenolic Profile and Antioxidant Properties of Salvia viridis L. Shoots: A Comparison of Aqueous and Hydroethanolic Extracts. Molecules. 2018; 23(6):1468. https://doi.org/10.3390/molecules23061468
Chicago/Turabian StyleGrzegorczyk-Karolak, Izabela, and Anna K. Kiss. 2018. "Determination of the Phenolic Profile and Antioxidant Properties of Salvia viridis L. Shoots: A Comparison of Aqueous and Hydroethanolic Extracts" Molecules 23, no. 6: 1468. https://doi.org/10.3390/molecules23061468