
The Use of Juçara (Euterpe edulis Mart.) Supplementation for Suppression of NF-κB Pathway in the Hypothalamus after High-Fat Diet in Wistar Rats

 , and
, and
Abstract
:1. Introduction
2. Results
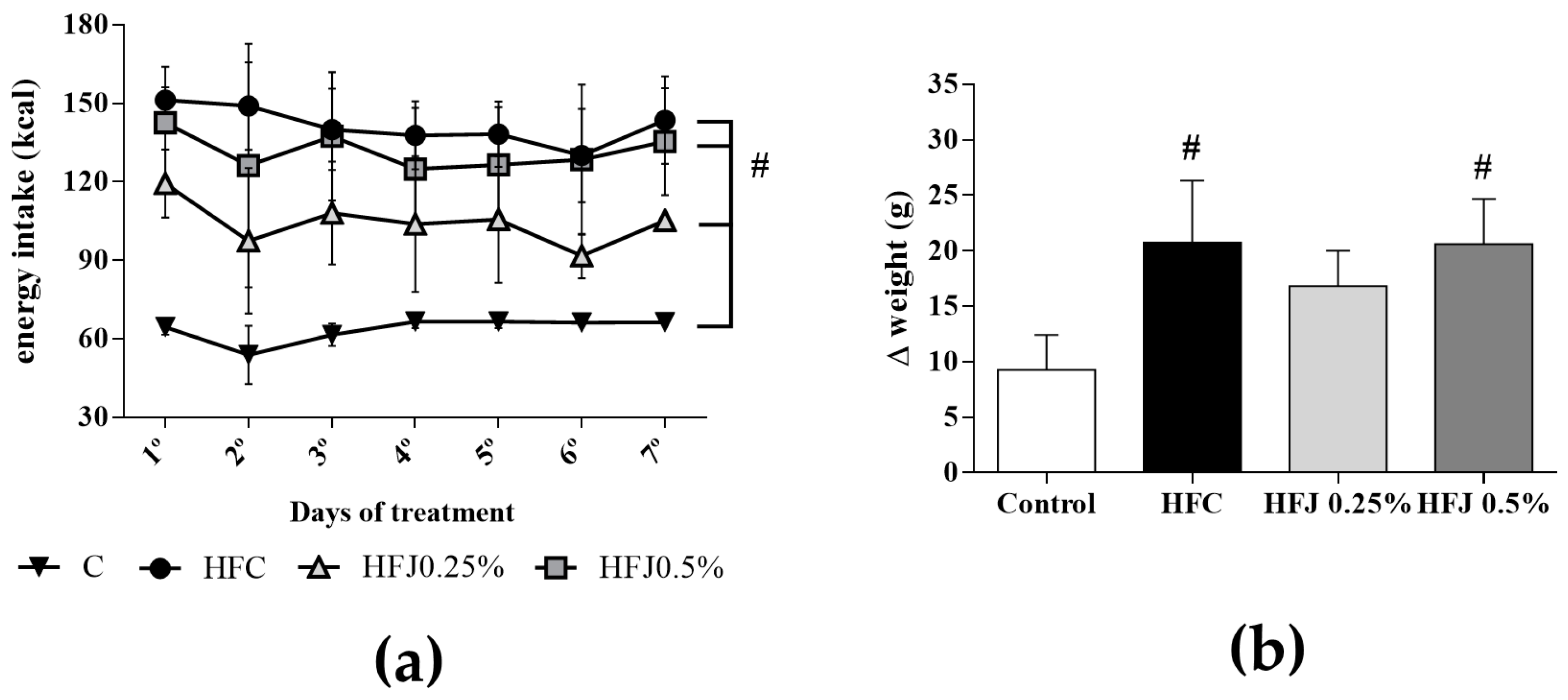
2.1. Energy Intake, Body and Tissue Weights
2.2. Lipoproteins, Triacylglycerol, Free Fatty Acids and Adiponectin
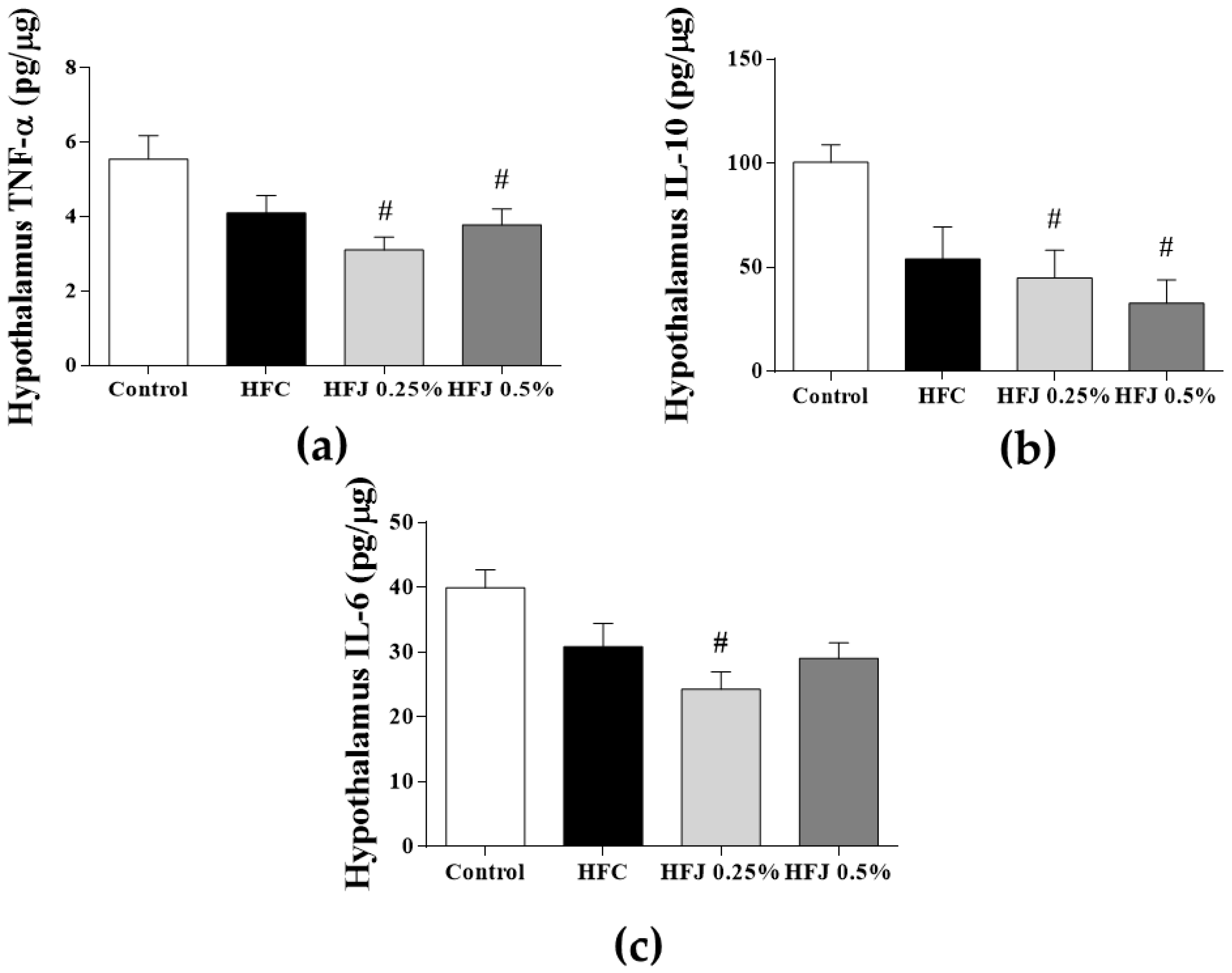
2.3. Hypothalamic Cytokine Concentration
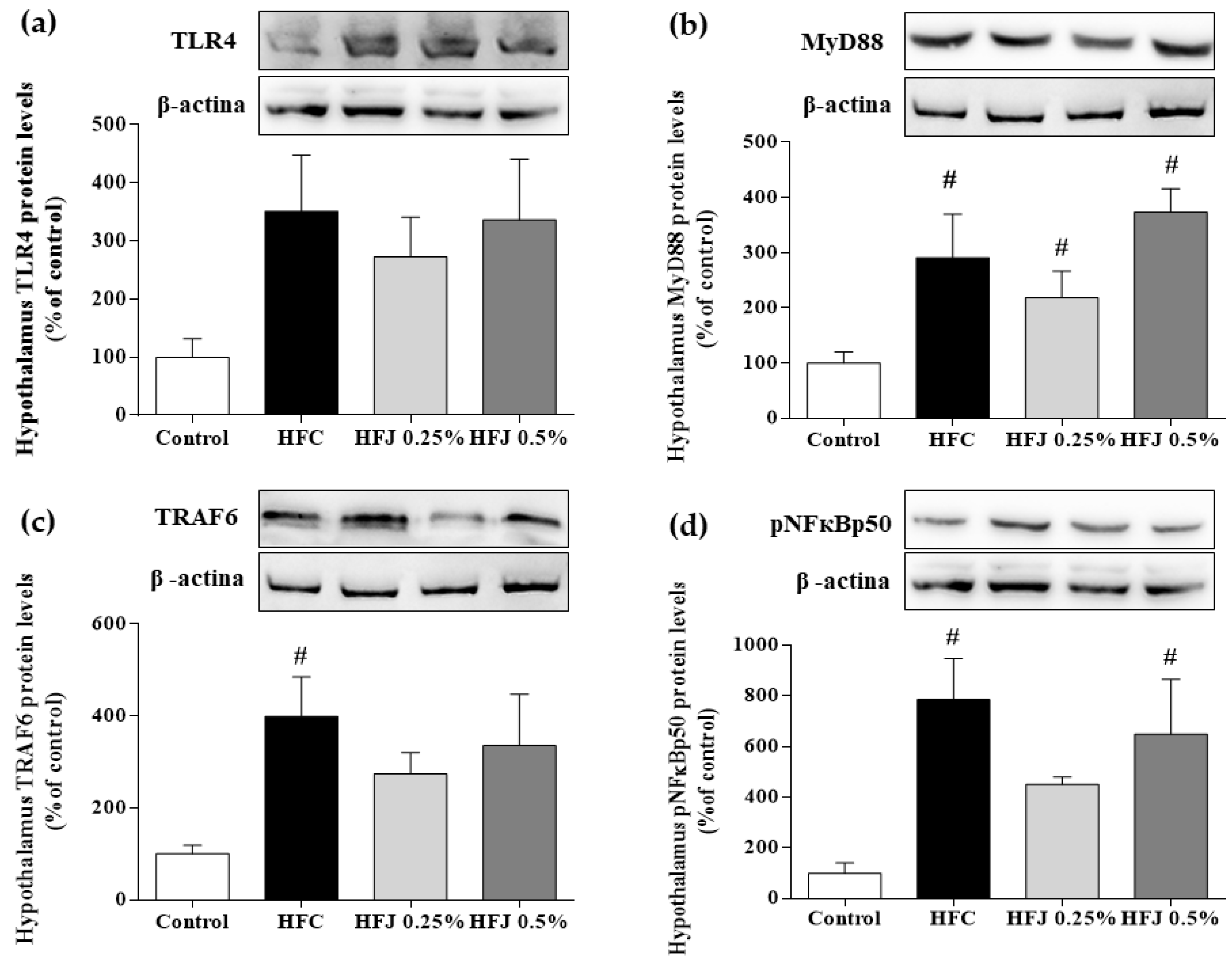
2.4. Hypothalamic NF-κB Pathway Protein Expression
3. Discussion
4. Materials and Methods
4.1. Freeze-Dried Juçara Pulp Powder
4.2. Animal Treatment
4.3. Serum Parameters
4.4. Tissue Cytokine Concentrations
4.5. Western Blot Analyses
4.6. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Myles, I.A. Fast food fever: Reviewing the impacts of the Western diet on immunity. Nutr. J. 2014, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Zobel, E.H.; Hansen, T.W.; Rossing, P.; von Scholten, B.J. Global changes in food supply and the obesity epidemic. Curr. Obes. Rep. 2016, 5, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Swinburn, B.; Caterson, I.; Seidell, J.; James, W. Diet, nutrition and the prevention of excess weight gain and obesity. Public Health Nutr. 2007, 7, 123–146. [Google Scholar] [CrossRef]
- Turner, N.; Kowalski, G.M.; Leslie, S.J.; Risis, S.; Yang, C.; Lee-Young, R.S.; Babb, J.R.; Meikle, P.J.; Lancaster, G.I.; Henstridge, D.C.; et al. Distinct patterns of tissue-specific lipid accumulation during the induction of insulin resistance in mice by high-fat feeding. Diabetologia 2013, 56, 1638–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogero, M.M.; Calder, P.C. Obesity, inflammation, toll-like receptor 4 and fatty acids. Nutrients 2018, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Namavar, M.R.; Raminfard, S.; Jahromi, Z.V.; Azari, H. Effects of high-fat diet on the numerical density and number of neuronal cells and the volume of the mouse hypothalamus: A stereological study. Anat. Cell Biol. 2012, 45, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Thaler, J.P.; Schwartz, M.W. Minireview: Inflammation and obesity pathogenesis: The hypothalamus heats up. Endocrinology 2010, 151, 4109–4115. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jiao, Y.; Xie, M. Paeoniflorin ameliorates atherosclerosis by suppressing TLR4-mediated NF-κB activation. Inflammation 2017, 40, 2042–2051. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black currant anthocyanins attenuate weight gain and improve glucose metabolism in diet-induced obese mice with intact, but not disrupted, gut microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef] [PubMed]
- Schreckinger, M.E.; Lotton, J.; Lila, M.A.; de Mejia, E.G. Berries from South America: A comprehensive review on chemistry, health potential, and commercialization. J. Med. Food 2010, 13, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Campelo Borges, G.; Vieira, F.G.K.; Copetti, C.; Gonzaga, L.V.; Zambiazi, R.C.; Mancini Filho, J.; Fett, R. Chemical characterization, bioactive compounds, and antioxidant capacity of jussara (Euterpe edulis) fruit from the Atlantic Forest in southern Brazil. Food Res. Int. 2011, 44, 2128–2133. [Google Scholar] [CrossRef]
- Schulz, M.; da Silva Campelo Borges, G.; Gonzaga, L.V.; Oliveira Costa, A.C.; Fett, R. Juçara fruit (Euterpe edulis Mart.): Sustainable exploitation of a source of bioactive compounds. Food Res. Int. 2016, 89, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Felzenszwalb, I.; da Costa Marques, M.R.; Mazzei, J.L.; Aiub, C.A. Toxicological evaluation of Euterpe edulis: A potential superfruit to be considered. Food Chem. Toxicol. 2013, 58, 536–544. [Google Scholar] [CrossRef] [PubMed]
- da Silva, N.A.; Rodrigues, E.; Mercadante, A.Z.; de Rosso, V.V. Phenolic compounds and carotenoids from four fruits native from the brazilian atlantic forest. J. Agric. Food Chem. 2014, 62, 5072–5084. [Google Scholar] [CrossRef] [PubMed]
- Morais, C.A.; Oyama, L.M.; de Moura Conrado, R.; de Rosso, V.V.; do Nascimento, C.O.; Pisani, L.P. Polyphenols-rich fruit in maternal diet modulates inflammatory markers and the gut microbiota and improves colonic expression of ZO-1 in offspring. Food Res. Int. 2015, 77, 186–193. [Google Scholar] [CrossRef]
- Das, N.; Sikder, K.; Bhattacharjee, S.; Majumdar, S.B.; Ghosh, S.; Majumdar, S.; Dey, S. Quercetin alleviates inflammation after short-term treatment in high-fat-fed mice. Food Funct. 2013, 4, 889. [Google Scholar] [CrossRef] [PubMed]
- Most, J.; Goossens, G.H.; Jocken, J.W.E.; Blaak, E.E. Short-term supplementation with a specific combination of dietary polyphenols increases energy expenditure and alters substrate metabolism in overweight subjects. Int. J. Obes. 2014, 38, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Dornellas, A.P.S.; Watanabe, R.L.H.; Pimentel, G.D.; Boldarine, V.T.; Nascimento, C.M.O.; Oyama, L.M.; Ghebremeskel, K.; Wang, Y.; Bueno, A.A.; Ribeiro, E.B. Deleterious effects of lard-enriched diet on tissues fatty acids composition and hypothalamic insulin actions. Prostaglandins Leukot. Essent. Fat. Acids 2015, 102–103, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, M.S.F.; Wueest, S.; Item, F.; Schoenle, E.J.; Konrad, D. Adipose tissue inflammation contributes to short-term high-fat diet-induced hepatic insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E388–E395. [Google Scholar] [CrossRef] [PubMed]
- Oyama, L.M.; Silva, F.P.; Carnier, J.; De Miranda, D.A.; Santamarina, A.B.; Ribeiro, E.B.; Oller Do Nascimento, C.M.; De Rosso, V.V. Jucąra pulp supplementation improves glucose tolerance in mice. Diabetol. Metab. Syndr. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Seymour, E.M.; Lewis, S.K.; Urcuyo-Llanes, D.E.; Tanone, I.I.; Kirakosyan, A.; Kaufman, P.B.; Bolling, S.F. Regular tart cherry intake alters abdominal adiposity, adipose gene transcription, and inflammation in obesity-prone rats fed a high fat diet. J. Med. Food 2009, 12, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Razquin, C.; Martinez, J.A.; Martinez-Gonzalez, M.A.; Mitjavila, M.T.; Estruch, R.; Marti, A. A 3 years follow-up of a Mediterranean diet rich in virgin olive oil is associated with high plasma antioxidant capacity and reduced body weight gain. Eur. J. Clin. Nutr. 2009, 63, 1387–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarreal-Molina, M.T.; Antuna-Puente, B. Adiponectin: Anti-inflammatory and cardioprotective effects. Biochimie 2012, 94, 2143–2149. [Google Scholar] [CrossRef] [PubMed]
- Moraes, J.C.; Coope, A.; Morari, J.; Cintra, D.E.; Roman, E.A.; Pauli, J.R.; Romanatto, T.; Carvalheira, J.B.; Oliveira, A.L.R.; Saad, M.J.; et al. High-fat diet induces apoptosis of hypothalamic neurons. PLoS ONE 2009, 4, e5045. [Google Scholar] [CrossRef] [PubMed]
- Menard, C.; Bastianetto, S.; Quirion, R. Neuroprotective effects of resveratrol and epigallocatechin gallate polyphenols are mediated by the activation of protein kinase C gamma. Front. Cell Neurosci. 2013, 7, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Cintra, D.E.; Ropelle, E.R.; Moraes, J.C.; Pauli, J.R.; Morari, J.; de Souza, C.T.; Grimaldi, R.; Stahl, M.; Carvalheira, J.B.; Saad, M.J.; et al. Unsaturated fatty acids revert diet-induced hypothalamic inflammation in obesity. PLoS ONE 2012, 7, e30571. [Google Scholar] [CrossRef] [PubMed]
- Thaler, J.P.; Yi, C.-X.; Schur, E.A.; Guyenet, S.J.; Hwang, B.H.; Dietrich, M.O.; Zhao, X.; Sarruf, D.A.; Izgur, V.; Maravilla, K.R.; et al. Obesity is associated with hypothalamic injury in rodents and humans. J. Clin. Investig. 2012, 122, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milanski, M.; Degasperi, G.; Coope, A.; Morari, J.; Denis, R.; Cintra, D.E.; Tsukumo, D.M.L.; Anhe, G.; Amaral, M.E.; Takahashi, H.K.; et al. Saturated fatty acids produce an inflammatory response predominantly through the activation of Tlr4 signaling in hypothalamus: Implications for the pathogenesis of obesity. J. Neurosci. 2009, 29, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Viggiano, E.; Mollica, M.P.; Lionetti, L.; Cavaliere, G.; Trinchese, G.; De Filippo, C.; Chieffi, S.; Gaita, M.; Barletta, A.; De Luca, B.; et al. Effects of an high-fat diet enriched in lard or in fish oil on the hypothalamic amp-activated protein kinase and inflammatory mediators. Front. Cell. Neurosci. 2016, 10, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, E.P.; Moraes, J.C.; Cintra, D.E.; Velloso, L.A. Mechanisms in endocrinology: Hypothalamic inflammation and nutrition. Eur. J. Endocrinol. 2016, 175, R97–R105. [Google Scholar] [CrossRef] [PubMed]
- Dragano, N.R.V.; Marques, A.Y.C.; Cintra, D.E.C.; Solon, C.; Morari, J.; Leite-Legatti, A.V.; Velloso, L.A.; Marósticar, M.R., Jr. Freeze-dried jaboticaba peel powder improves insulin sensitivity in high-fat-fed mice. Br. J. Nutr. 2013, 110, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, N.J.; Norris, G.H.; Ryan, J.; Porter, C.M.; Jiang, C.; Blesso, C.N. Black elderberry extract attenuates inflammation and metabolic dysfunction in diet-induced obese mice. Br. J. Nutr. 2015, 114, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Buettner, R.; Parhofer, K.G.; Woenckhaus, M.; Wrede, C.E.; Kunz-Schughart, L.A.; Schölmerich, J.; Bollheimer, L.C. Defining high-fat-diet rat models: Metabolic and molecular effects of different fat types. J. Mol. Endocrinol. 2006, 36, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Blachnio-Zabielska, A.; Baranowski, M.; Zabielski, P.; Gorski, J. Effect of high fat diet enriched with unsaturated and diet rich in saturated fatty acids on sphingolipid metabolism in rat skeletal muscle. J. Cell. Physiol. 2010, 225, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Argentato, P.P.; Morais, C.A.; Santamarina, A.B.; de Cassia César, H.; Estadella, D.; de Rosso, V.V.; Pisani, L.P. Jussara (Euterpe edulis Mart.) supplementation during pregnancy and lactation modulates UCP-1 and inflammation biomarkers induced by trans-fatty acids in the brown adipose tissue of offspring. Clin. Nutr. Exp. 2017, 12, 50–65. [Google Scholar] [CrossRef]
- O’Neill, H.M.; Holloway, G.P.; Steinberg, G.R. AMPK regulation of fatty acid metabolism and mitochondrial biogenesis: Implications for obesity. Mol. Cell. Endocrinol. 2013, 366, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Ren, J.; Huang, Z.; Wang, T.; Xiang, K.; Cheng, L.; Tang, L. The role of exogenous hydrogen sulfide in free fatty acids induced inflammation in macrophages. Cell. Physiol. Biochem. 2017, 42, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, G.; Vaccaro, O.; Costabile, G.; Rivellese, A.A. How well can we control dyslipidemias through lifestyle modifications? Curr. Cardiol. Rep. 2016, 18, 66. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Anderson, R.A. An extract of chokeberry attenuates weight gain and modulates insulin, adipogenic and inflammatory signalling pathways in epididymal adipose tissue of rats fed a fructose-rich diet. Br. J. Nutr. 2012, 108, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Mykkänen, O.T.; Huotari, A.; Herzig, K.H.; Dunlop, T.W.; Mykkänen, H.; Kirjavainen, P.V. Wild blueberries (vaccinium myrtillus) alleviate inflammation and hypertension associated with developing obesity in mice fed with a high-fat diet. PLoS ONE 2014, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.; Carmo, L.; Silva, G.; Silveira-diniz, M.; Casemiro, R.; Spoto, M. Physical, Chemical, and Lipid Composition of Juçara (Euterpe edulis Mart.) Pulp. Braz. J. Food Nutr. 2013, 24, 7–13. [Google Scholar]
- Karlsen, A.; Retterstøl, L.; Laake, P.; Paur, I.; Bøhn, S.K.; Sandvik, L.; Blomhoff, R. Anthocyanins inhibit nuclear factor-kappaB activation in monocytes and reduce plasma concentrations of pro-inflammatory mediators in healthy adults. J. Nutr. 2007, 137, 1951–1954. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xia, M.; Ma, J.; Hao, Y.; Liu, J.; Mou, H.; Cao, L.; Ling, W. Anthocyanin supplementation improves serum LDL-and HDL-cholesterol concentrations associated with the inhibition of cholesteryl ester transfer protein in dyslipidemic subjects. Am. J. Clin. Nutr. 2009, 90, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Center for Drug Evaluation and Research. Estimating the maximum safe starting dose in initial clinical trials for therapeutics in adult healthy volunteers. In Guidance for Industry; US Department of Health and Human Services: Washington, DC, USA, 2005; pp. 1–27. [Google Scholar] [CrossRef]
- Graf, D.; Seifert, S.; Jaudszus, A.; Bub, A.; Watzl, B. Anthocyanin-rich juice lowers serum cholesterol, leptin, and resistin and improves plasma fatty acid composition in fischer rats. PLoS ONE 2013, 8, e66690. [Google Scholar] [CrossRef] [PubMed]
- Brasil, C. Diretriz Brasileira Para o Cuidado e a Utilização de Animais Para Fins Científicos e Didáticos; Ministério da Ciência Tecnologia e Inovação: Brasília, Brazil, 2013; pp. 1–50. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Almeida Morais, C.; Oyama, L.M.; de Oliveira, J.L.; Carvalho Garcia, M.; de Rosso, V.V.; Sousa Mendes Amigo, L.; do Nascimento, C.M.; Pisani, L.P. Jussara (Euterpe edulis Mart.) Supplementation during Pregnancy and Lactation Modulates the Gene and Protein Expression of Inflammation Biomarkers Induced by trans-Fatty Acids in the Colon of Offspring. Med. Inflamm. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mennitti, L.V.; Oyama, L.M.; de Oliveira, J.L.; Hachul, A.C.; Santamarina, A.B.; de Santana, A.A.; Okuda, M.H.; Ribeiro, E.B.; do Nascimento, C.M.; Pisani, L.P.; et al. Oligofructose supplementation during pregnancy and lactation impairs offspring development and alters the intestinal properties of 21-d-old pups. Lipids Health Dis. 2014, 13, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Tissue | (g/100 g of Body Mass) | |||||||
|---|---|---|---|---|---|---|---|---|
| Control (n = 6) | HFC (n = 7) | HFJ 0.25% (n = 7) | HFJ 0.5% (n = 7) | |||||
| Mean | S.E.M. | Mean | S.E.M. | Mean | S.E.M. | Mean | S.E.M. | |
| Hypothalamus | 0.03 | 0.003 | 0.02 #,* | 0.001 | 0.03 | 0.002 | 0.03 | 0.001 |
| Liver | 2.92 | 0.09 | 2.81 | 0.05 | 2.87 | 0.07 | 2.80 | 0.03 |
| RET | 0.93 | 0.13 | 1.68 # | 0.14 | 1.27 | 0.14 | 1.39 | 0.15 |
| EPI | 1.03 | 0.085 | 1.37 | 0.112 | 1.16 | 0.12 | 1.06 | 0.14 |
| MES | 0.98 | 0.16 | 1.03 | 0.10 | 0.96 | 0.03 | 1.09 | 0.09 |
| ΣWAT | 2.94 | 0.32 | 4.07 # | 0.21 | 3.39 | 0.26 | 3.54 | 0.30 |
| Parameter | Experimental Groups | |||||||
|---|---|---|---|---|---|---|---|---|
| Control (n = 6) | HFC (n = 7) | HFJ 0.25% (n = 7) | HFJ 0.5% (n = 7) | |||||
| Mean | S.E.M | Mean | S.E.M. | Mean | S.E.M. | Mean | S.E.M. | |
| triacylglycerol (mg/dL) | 104.84 | 2.91 | 117.96 | 7.15 | 109.84 | 4.86 | 120.55 *,# | 6.70 |
| total cholesterol (mg/dL) | 108.13 | 6.24 | 136.40 # | 10.44 | 118.04 | 6.02 | 133.41 | 12.38 |
| HDL-Cholesterol (mg/dL) | 54.72 | 3.44 | 46.21 | 4.05 | 48.86 | 3.96 | 47.11 | 4.30 |
| LDL-Cholesterol (mg/dL) | 41.30 | 9.49 | 68.32 #,* | 5.15 | 50.79 | 9.38 | 58.42 | 9.95 |
| Free fatty acids (mM/mL) | 1.87 | 1.65 | 2.05 * | 2.74 | 1.39 | 5.40 | 1.97 | 2.08 |
| Adiponectin (ng/mL) | 1.77 | 0.029 | 1.92 | 0.33 | 2.69 | 0.48 # | 1.95 | 0.32 |
| Juçara Pulp | Concentration in 100 g of Fresh Matter | References | |
|---|---|---|---|
| Mean | S.E.M | ||
| Moisture (%) | 88.7 | 3.8 | [14] |
| Carbohydrates (g) | 28.3 | 3.5 | [14] |
| Proteins (g) | 6.0 | 0.3 | [14] |
| Lipids (g) | 29.2 | 0.9 | [14] |
| Palmitic acid (%) | 34.43 | 3.42 | [41] |
| Stearic acid (%) | 3.01 | 0.30 | [41] |
| SAT (%) | 37.44 | [41] | |
| Palmitoleic acid (%) | 2.61 | 0.26 | [41] |
| Oleic acid (%) | 35.96 | 3.08 | [41] |
| MUFA (%) | 38.57 | [41] | |
| Linoleic acid (%) | 19.18 | 1.89 | [41] |
| Linolenic acid (%) | 0.91 | 0.20 | [41] |
| PUFA (%) | 20.08 | [41] | |
| Fiber (g) | 28.3 | 0.3 | [14] |
| Ashes (g) | 8.8 | 0.8 | [14] |
| Energetic values (kcal) | 400.0 | 23.9 | [14] |
| Cyanidin 3-rutinoside (mg) | 191.0 | 6.5 | [15] |
| Cyanidin 3-glucoside (mg) | 71.4 | 2.1 | [15] |
| Total anthocyanins (mg) | 262.4 | 8.6 | [15] |
| Apigenin deoxyhexosyl-hexoside (mg) | 25.4 | 1.5 | [14] |
| Luteolin deoxyhexosyl-hexoside (mg) | 37.6 | 1.9 | [14] |
| Dihydrokaempferol-hexoside (mg) | 66.4 | 2.6 | [14] |
| Total phenolic compounds (mg) | 415.1 | 22.3 | [14] |
| Ingredients | Diet (g/100 g) | |||
|---|---|---|---|---|
| Control | HF Control | HF Juçara 0.25% | HF Juçara 0.5% | |
| Standard chow | 100 | 50 | 50 | 50 |
| Sucrose | - | 10 | 10 | 10 |
| Casein | - | 20 | 20 | 20 |
| Soybean Oil | - | 2 | 2 | 2 |
| Lard | - | 18 | 18 | 18 |
| Butyl hydroquinone | - | 0.004 | 0.004 | 0.004 |
| Juçara pulp powder | - | - | 0.25 | 0.5 |
| Protein | 22.4 | 23.6 | 23.6 | 23.6 |
| Total fat | 4.8 | 22 | 22 | 22 |
| Carbohydrates | 39.1 | 26.8 | 26.8 | 26.8 |
| Alimentary fiber | 11.4 | 15.1 | 15.1 | 15.1 |
| Mineral residues | 11.9 | 9 | 9 | 9 |
| Energy (Kcal/100 g) | 270 | 410 | 420 | 430 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamarina, A.B.; Jamar, G.; Mennitti, L.V.; De Rosso, V.V.; Cesar, H.C.; Oyama, L.M.; Pisani, L.P. The Use of Juçara (Euterpe edulis Mart.) Supplementation for Suppression of NF-κB Pathway in the Hypothalamus after High-Fat Diet in Wistar Rats. Molecules 2018, 23, 1814. https://doi.org/10.3390/molecules23071814
Santamarina AB, Jamar G, Mennitti LV, De Rosso VV, Cesar HC, Oyama LM, Pisani LP. The Use of Juçara (Euterpe edulis Mart.) Supplementation for Suppression of NF-κB Pathway in the Hypothalamus after High-Fat Diet in Wistar Rats. Molecules. 2018; 23(7):1814. https://doi.org/10.3390/molecules23071814
Chicago/Turabian StyleSantamarina, Aline Boveto, Giovana Jamar, Laís Vales Mennitti, Veridiana Vera De Rosso, Helena Cassia Cesar, Lila Missae Oyama, and Luciana Pellegrini Pisani. 2018. "The Use of Juçara (Euterpe edulis Mart.) Supplementation for Suppression of NF-κB Pathway in the Hypothalamus after High-Fat Diet in Wistar Rats" Molecules 23, no. 7: 1814. https://doi.org/10.3390/molecules23071814