An Ethnopharmacological, Phytochemical and Pharmacological Review on Lignans from Mexican Bursera spp.



Department of Pharmaceutical Sciences, University of Perugia, via del Liceo, 1-06123 Perugia, Italy
*
Author to whom correspondence should be addressed.
Molecules 2018, 23(8), 1976; https://doi.org/10.3390/molecules23081976
Submission received: 5 July 2018
/
Revised: 1 August 2018
/
Accepted: 5 August 2018
/
Published: 8 August 2018
(This article belongs to the Special Issue Lignans)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The genus Bursera belongs to the family Burseraceae and has been used in traditional Mexican medicine for treating various pathophysiological disorders. The most representative phytochemicals isolated from this genus are terpenoids and lignans. Lignans are phenolic metabolites known for their antioxidant, apoptotic, anti-cancer, anti-inflammatory, anti-bacterial, anti-viral, anti-fungal, and anti-protozoal properties. Though the genus includes more than 100 species, we have attempted to summarize the biological activities of the 34 lignans isolated from selected Mexican Bursera plants.
1. Introduction
The genus Bursera Jacq. ex L. (family Burseraceae, order Sapindales), named after the Danish botanist Joachim Burser (1583–1639), is a monophyletic genus [1] that includes about 105 species of small trees and shrubs distributed from Southern U.S. to Peru and the Caribbean, particularly in Mexico (ca. 92 species) [2]. These plants are characterized by the production of resins that are exuded from the trunk and leaves and provide a chemical defense against specialized herbivores [3]. Once dried, the resin obtained by Bursera spp. is called “copal”, a term which is also used to describe a large group of resins characterized by hardness and a relative high melting point, which also is found in other plants [4]. The loss of the essential oils and the oxidation and polymerization processes transform copal into amber.
The phytochemistry of this genus is characterized by the presence of volatile metabolites such as simple hydrocarbons and terpenoids as well as phenolics [5,6,7,8]. Among the compounds present in the volatile fraction, heptane, α- and β-pinene, β-phellandrene, and limonene are among the most frequent [5], whereas β-caryophyllene and germacrene D are the most common sesquiterpenes in the genus Bursera [8]. Cembrane and verticillane diterpenoids are often present [9,10,11]. Pentacyclic triterpenoids are largely present in the resin of several species, and the study of triterpenoidic composition of resins is important to define the botanical origin of archaeological samples of copal [12]. In Bursera microphylla resin, malabaricane triterpenoids were also found [11]. Leaves and branches of some Bursera also contain flavonoids [13,14,15] and luteolin 3′-O-rhamnoside is very common [13].
Lignans are naturally occurring plant phenolics, biosynthetically derived from phenylpropanoids, that are important components in foods and medicines; their chemical and biological properties have been reviewed [16]. The aim of this review is to summarize literature findings on the botanical characterization, distribution, ethnopharmacology, and biological activities of Mexican Bursera that produce lignans. Different Bursera species have been sorted according to, and synonyms are those reported in, the Plant List Database [17]. Unless otherwise specified, common names are those reported by Lemos and Rivera [18]. The phytochemistry was analyzed by data reported in the SciFinder database. Images of the species reported in this paper can be found on the Enciclovida web site [19].
2. Genus Bursera
Members of this genus are typically small- to medium-sized trees or shrubs, mostly dioecious, succulent, and with resin canals in vascularized tissues. Their leaves are deciduous, imparipinnate, or sometimes unifoliolate or trifoliolate (occasionally bipinnate). Their flowers are small, almost always unisexual, three- to six-merous. Their fruit is a dehiscent two- to three-valve drupe with a fleshy to coriaceous skin, and with a pyrene (the stone or pit that contains the seed), cartilaginous to bony, enveloped totally or partially by an arillate structure.
The taxonomy of the genus Bursera is based on morphological characteristics of fruit, bark, and leaves, as well as molecular data. Currently, there are two recognized subgenera: one subgenus named Bursera (previously called section Bursera) that includes species commonly known with the general vernacular name of “cuajiotes”, and the other is called Elaphrium (previously called section Bullockia) that comprises species with the general common name of “copales” [20,21,22]. The most conspicuous difference between the subgenera is the bark: in subgenus Bursera, it tends to be colorful and exfoliating, whereas in Bursera subgenus Elaphrium, it is likely to be complete (not exfoliating), and grey or reddish grey. However, although bark helps in species identification because it is easy to see, whether the bark is complete or exfoliating is not an absolute difference between the two groups. Setting the bark aside, the most reliable distinction between the two subgenera is the number of locules in the ovary (three in subg. Bursera vs. two in Elaphrium), and the number of valves in the fruit (three in subg. Bursera vs. two in Elaphrium) [20,21]. Another distinguishing trait is the presence of well-developed cataphylls (small bract-like leaves that appear before “true leaves” and are short-lived) in subgenus Elaphrium and absent or very inconspicuous in subgenus Bursera [23]. Toledo further divided section Bursera into three groups that can be distinguished by the color of the exfoliating bark: mulatos, red cuajiotes, and yellow cuajiotes [24]. Furthermore, the section Bullockia was divided into two groups: pseudoaril-covered fruits group and partially covered fruits group [24]. In 1980, Gillet changed the name of section Bullockia into Elaphrium due to the fact that some characteristics of this section resemble those of Elaphrium tomentosum Jacq. [25]. Phylogeny studies by Becerra and Venable allowed the recognition of four different groups in section Bursera: the simaruba group (massive trees, trilobate cotyledons, red exfoliating bark, poor producing resin (mulatos)), the microphylla group (medium-sized trees or shrubs, multilobate cotyledons, yellow to red exfoliating bark, highly resinous), the fagaroides group (medium-sized trees or shrubs, multilobate cotyledons, highly resinous), and the fragilis group (medium-sized trees, multilobate cotyledons, red exfoliating bark, highly resinous (cuajiotes)) [22]; and two groups in Elaphrium: copallifera (seed completely or at least two-thirds covered by pseudoaril) and glabrifolia (seed partially covered or at least less than two-thirds by pseudoaril) [5]. Bursera’s flowers are small and inconspicuous, with few species-specific characteristics, occurring during bursts during the dry season. Thus, in their natural habitats, it is often easier to recognize them by their bark and leaf characteristics as well as the locations where they grow. The Bursera genus is closely related to the other two resin producing Burseraceae: Boswellia and Commiphora, and they differ mostly in their geographic distribution. Boswellia and Commiphora are present in desert parts of tropical Africa, Arabia, Pakistan, and India, whereas Bursera is distributed from the Southern U.S. to Peru and the Caribbean, and particularly in Mexico. The Bursera section shares some similarities with Boswellia, whereas Elaphrium is similar to Commiphora [26].
2.1. Traditional Uses, Phytochemistry, and Biological Activities
Most of the Bursera species that produce lignans are widely used by the Mexican native population. Although different Bursera species are used for different health issues, they are traditionally attributed medicinal properties including providing relief from pain, inflammation, rheumatism, and can help treat illnesses such as colds, skin tumors, polyps, and venereal diseases [27,28,29,30]. The following are reported lignan-producing Bursera plants from Mexico listed according to the subgenus they belong to and sorted alphabetically. Traditional uses, when found, and biological properties of the isolated compounds are described for each species.
2.2. Subgenus Elaphrium
2.2.1. Bursera citronella McVaugh and Rzed.
B. citronella (synonyms: none reported) is also known as xochicopal (Náhuatl name) or lináloe, and as almárciga in Spanish [31]. It is a 10 m tree with grey trunk bark and unifoliate or trifoliolate leaves, distributed in Western Mexico (Michoacan, Colima, Jalisco, and Guerrero). It belongs to the subsection glabrifolia [5]. The resin is mostly used as incense. It has been reported that B. citronella is used as antitussive in several regions in Mexico [32].
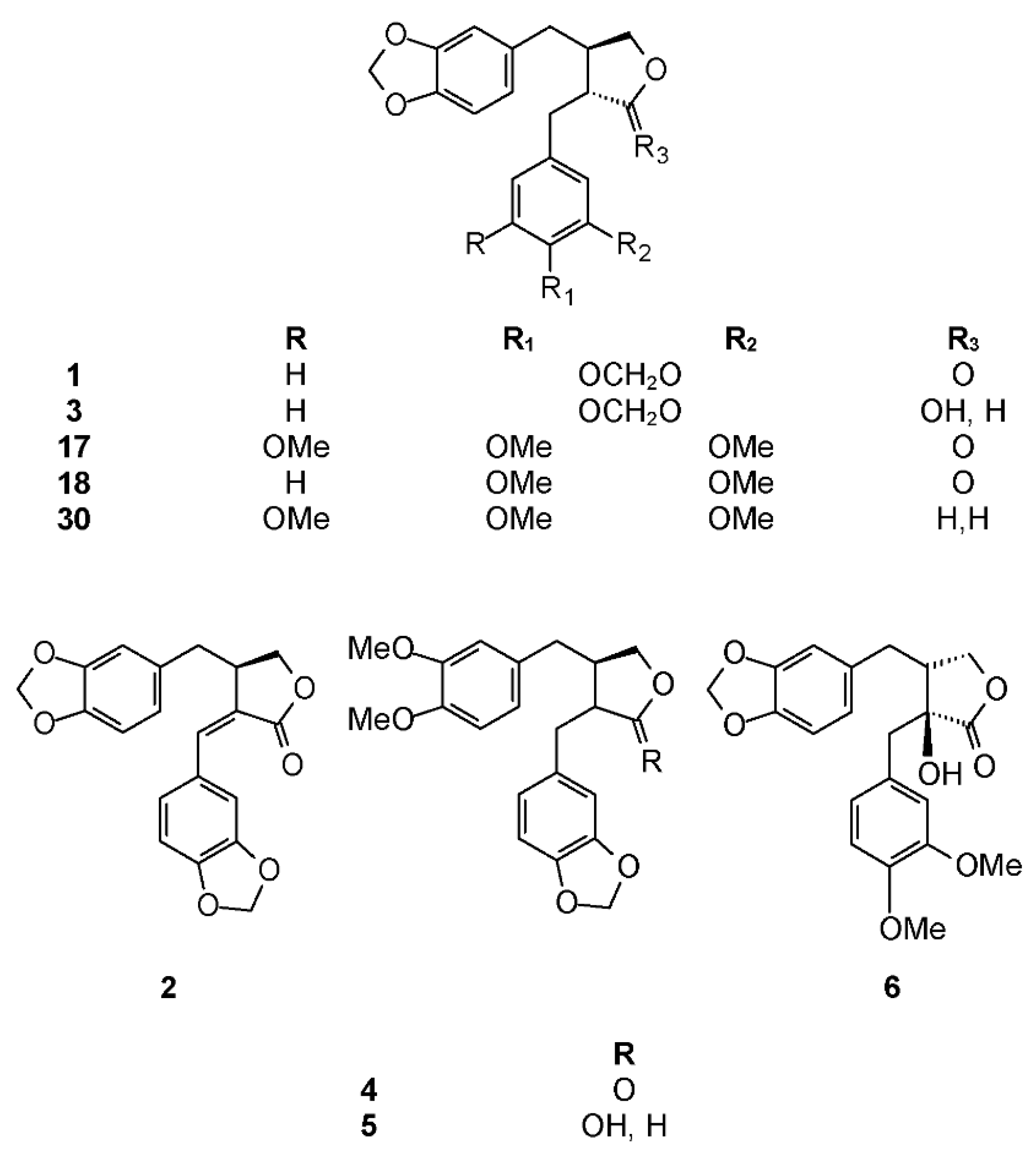
The phytochemistry of B. citronella has been studied by Koulman who isolated two lignans: hinokinin (1) and savinin (2) (Figure 1) [33]. Biological activities of hinokinin have been recently reviewed [34]. Cytotoxicity of hinokinin (1) has been tested by several authors [35,36,37,38]. Hinokinin (1) has been shown to have anti-inflammatory [39,40,41,42], immunosuppressive [43,44], antibacterial [45], and antiviral [46] properties. Hinokinin (1) was tested for several other biological activities, such as antispasmodic [47], neurite outgrowth-promoting in PC12 cells [48], antileukemic [49], antiproliferative [50], and neuroprotective activities [51]. This compound showed an interesting activity against Trypanosoma cruzi, [52,53,54,55,56], but with a low parasite selectivity [57]. Hinokinin has been chosen as a trypanosomicidal marker in P. cubeba [58]. In order to ascertain the safety of this compound toward mammalian cells, several studies have been performed [59,60,61]. The authors found that hinokinin did not increase DNA damage, demonstrating the absence of mutagenic and genotoxic activities. On the other hand, the results on the antimutagenic potential of this compound showed a strong inhibitory effect against some direct and indirect-acting mutagens.
Savinin (2), also called hibalactone [62], was isolated for the first time from Juniperus spp. [63]. It has been tested for different biological activities, such as cytotoxicity against several tumor cell lines [36,64,65,66]. Savinin (2) was shown to have an anti-inflammatory activity in several assays [40,44,67,68,69], but also interesting antinociceptive, anxiolytic, and antioxidant activities [70,71].
2.2.2. Bursera cuneata (Schltdl.) Engl.
B. cuneata (synonym: Elaphrium cuneatum Schltdl. L.) is a tree that grows up to 10 m in height with no-peeling grey-reddish bark. It has imparipinnate leaves of coriaceous texture with 3 to 13 leaflets, 6.5 cm long and 2.3 cm wide, and margin roughly serrated. Their flowers are clustered in inflorescences up to 8 cm long. Its flowers are white and its fruits are up to 1.2 cm long with a black pit, almost completely covered by a yellow or orange pseudoaril. It is native to Mexican oak-tropical deciduous forest transition zones from Jalisco to Oaxaca and is often known as copal or copalillo [72]. Although B. cuneata is characterized by seeds covered by pseudoaril, Becerra and Venable did not classify it into the copallifera group [22]. No medical uses have been reported for this species, but it is largely used as incense during sacred ceremonies and to prepare handcrafted objects.
Koulman isolated three lignans from this species: hinokinin (1), savinin (2), and cubebin (3) (Figure 1) [33]. Cubebin was first isolated by Chatterjee in 1968 from Piper cubeba [73] and then from several Aristolochia spp. Biological activities of cubebin have been recently reviewed by Cunha et al. [16]. In particular, trypanocidal activity of this compound against free amastigote forms of Trypanosoma cruzi has been studied by de Souza et al. [52], and Bastos et al. showed it is inactive against trypomastigote forms [74]. Notably, cubebin (3) is usually the starting material for the semi-synthetic preparation of hinokinin (1) and other lignans [52]. Recently, cubebin was proven to induce vasorelaxation via nitric oxide activation without prostacyclin involvement [75]. Because of its therapeutic potential, the effects of cubebin on mutagenicity and genotoxicity has been deeply studied by several research groups [76]. The authors found that cubebin (3) was cytotoxic at high doses (280 μM), but at lower concentrations, no cytotoxic, mutagenic, or proliferative effects were observed for this compound. The mutagenicity of cubebin (3), alone or in combination with doxorubicin (DXR), using standard (ST) and high bioactivation (HB) crosses of the wing Somatic Mutation And Recombination Test (SMART) in Drosophila melanogaster was also studied [77]. Even in this case, the effect of cubebin was dose-dependent. At lower doses (<1 mM), it reduces DXR toxicity, whereas at higher doses (>2.0 mM), it is cytotoxic. The biological activities of cubebin have been recently reviewed [78].
2.2.3. Bursera excelsa (Kunth) Engl.
B. excelsa (synonyms: Bullockia sphaerocarpa and Elaphrium excelsum) is commonly known as tecomajaca, or copal santo, pom (in the Maya language) [79] and tecomahaca in the Náhuatl language [29]. It belongs to the copallifera group. These are trees up to 8 m tall with grey non-peeling bark. Their leaves are 11 to 23 cm long and 6 to 10.5 cm wide, with winged raquis and 9 to 15 leaflets, hairy and margin conspicuously toothed. Their flowers are small, yellow, and densely hairy. Traditionally, it is used to treat tumors and muscle spasms [29]. It is widely distributed across Mexico from the state of Sinaloa to Chiapas. The phytochemistry of B. excelsa has been extensively studied [8,12,13]. Regarding lignan composition, Koulman isolated three compounds from this species: 3,4-dimethoxy-3′,4′-methylenedioxylignano-9,9′-lactone (4), 3,4-dimethoxy-3′,4′-methylenedioxylignano-9,9′-epoxylignan-9′-ol (DME) (5), and guayadequiol (6) (Figure 1) [33]. Compound 4 was named iso-bursehernin “…since the only difference between this compound and bursehernin is the placement of the aromatic groups in relation to the lactone ring” and compound 5 was named DME [33]. Iso-bursehernin (also called kusunokinin) was isolated from several plants, such as Cinnamomun camphora [80], Virola spp. [81,82], and from different species of Haplophyllum [83,84,85]. Compounds 4 and 5 were identified as active glioma inhibitors in a bioassay-guided isolation process from Piper nigrum fruits [86] and 4 selectively docked to Leishmania mexicana pyruvate kinase in a study to find potential antiprotozoal polyphenolic plant extracts [87]. Kusunokinin isolated from P. nigrum showed potent cytotoxic activity on breast cancer cells (MCF-7 and MDA-MB-468) with IC50 values of 1.18 and 1.62 μg/mL, respectively, but demonstrated lower cytotoxicity on normal breast cell lines (IC50 higher than 11 μg/mL). Cell cycle studies showed that this compound induced cell apoptosis and drove cells toward the G2/M phase. Moreover, it decreased topoisomerase II and Bcl-2. The authors observed an increasing in p53, p21, bax, cytochrome c, and caspase-8, -7, and -3 activities, except caspase-9, suggesting that kusunokinin has potent anticancer activity through the extrinsic pathway and G2/M phase arrest [88].
Lignans that contain a methylenedioxy group show high antifeedant or deterrent activity against insects. Polar substituents on the aromatic rings, such as hydroxyl or glycosyl groups, reduce this activity. Guayadequiol (6) was isolated for the first time from Bupleurum salicifolium [89]. No biological data for this compound have been reported. The hexane extract of B. excelsa was shown to possess in vivo anti-inflammatory activity [29].
2.2.4. Bursera graveolens (Kunth) Triana and Planch.
B. graveolens (synonyms: Amyris caranifera, A. graveolens, B. andersonii, B. pilosa, B. tatamaco, Elaphrium graveolens, E. pilosum, Terebinthus graveolens, and T. pilosa) is called copal and mizquixochicopalli in Náhuatl language [90]. Its Spanish common name is palo santo and it is native to the tropical dry forests from the Yucatan Peninsula of Mexico, south to Peru, and the Galapagos Islands of Ecuador. These are trees and sometimes shrubs up to 15 m tall, highly fragrant, with grey bark. Their leaves are imparipinnate, sometimes bipinnate, up to 30 cm long and 18 cm wide, with 7 to 11 leaflets. The leaflets are 3 to 9 cm long and 1 to 4 cm wide, of acuminate apex, and margin roughly serrated. Their small flowers are yellowish, white, or green and their fruits glabrous and are up to 1.0 cm long. Their seeds are black and about two-thirds covered by an orange-red pseudoaril. Traditionally, the alcoholic extract of the bark is used for rheumatism, and the bark infusion, as a digestive and for respiratory problems. In recent years, the resin and oils have been extracted from the wood by the perfume industry. From the active methanol extract of stems, Nakanishi et al. isolated a new aryltetralin lignan, burseranin (7), and picropolygamain (8) (Figure 2) along with known triterpenes, lupeol and epi-lupeol [91]. The two isolated lignans 7 and 8 showed important cytotoxic activity against the human HT1080 fibrosarcoma cell line. Both compounds exhibited potent inhibitory effects in comparison with adriamycin as a positive control (5.5 and 1.9 μg/mL vs. 0.1 μg/mL). Picropolygamain (8) was isolated for the first time in 1985 from Commiphora incisa resin [92] and later from Bursera simaruba [93]. This compound was shown to be active against LNCaP (androgen-sensitive human prostate adenocarcinoma) cell line (ED50 1.1 μg/mL) during tests aimed at developing an in vivo Hollow Fiber Assay [94].
2.2.5. Bursera penicillata (Sessé and Moç. ex DC.) Engl.
B. penicillata (synonyms: Amyris penicillata, Bursera mexicana, Elaphrium delpechianum, E. mexicanum, E. penicillatum, Terebinthus delpechiana, and T. mexicana) belongs to the section glabrifolia. Its common names are coyoluche, torote incienso, and torote copal [95]. These are trees up to 12 m tall of grey or reddish grey bark, and are very fragrant, even sometimes from a distance. Their leaves are imparipinnate, 12 to 38 cm long, rachis-winged, and with 3 to 15 leaflets. The leaf blades are finely pubescent on both surfaces and the margins strongly toothed. Their flowers are small, white, and arranged in few to many inflorescences up to 14 cm long. Fruits are 1 to 1.3 cm long, 0.8 to 1.1 cm wide with a black pit, and partially covered by a red, orange, or pale pseudoaril. Endemic to Northwest Mexico, this species prospers in tropical deciduous forests and sporadically thornscrub and transition areas to oak woodland, from Southeastern Sonora and Southwest Chihuahua to Michoacan. According to Gentry, the leaves are used to treat the common cold and the gum is used for toothaches. It is also used as incense [96]. Koulman reported the presence of savinin (2) in this species [33].
2.2.6. Bursera submoniliformis Engl.
B. submoniliformis (synonyms: Bursera subsessiliformis Engl. and Elaphrium submoniliforme (Engl.) Marchand ex Engl.) is commonly known as copal chino. These trees are up to 12 m tall with grey to reddish gray bark. Their leaves are imparipinnate, up to 20 cm long and 7 cm wide, with 9 to 17 leaflets. The leaflets are velvety, 1.3 to 5 cm long, 0.5 to 2 cm wide, and have toothed margins. They have small white flowers that are arranged in inflorescences. Fruits are 7.5 to 12 mm long with a black pit almost or completely covered by a yellow or orange pseudoaril. Endemic to Mexico, this species inhabits tropical deciduous forests at altitudes of 500 to 1600 m of the Balsas and Papaloapan river basins in the states of Mexico, Michoacan, Guerrero, Puebla, Morelos, and Oaxaca. It belongs to the subsection copallifera. The gum resin is used to alleviate pain associated with flatulence and tooth-ache [97]. The only reference about the phytochemistry of B. submoniliformis is by Koulman, who reported the presence of savinin (2) in this species [33].
2.3. Section Bursera
2.3.1. Bursera aptera Ramirez
B. aptera (synonyms: Elaphrium apterum and Terebinthus aptera) belongs to the section fagaroides. Its common names are cuajiote verde [98,99] and cuajiote blanco (Náhuatl names). The species is distributed in Guerrero, Morelos, Oaxaca, and Puebla regions in Mexico [2]. These are shrubs or trees up to 10 m high with green trunks and bark that exfoliates in yellow or beige papyrus-like sheets. The leaves are glabrous, 2.5–7.5 cm long comprising 4 to 9 pairs of leaflets up to 15 mm long and 6 mm wide. The flowers are reddish, yellow, or white and the fruits are small, up to 7 mm long, greyish red when mature, and with a pit completely covered by a yellow or white papery pseudoaril.
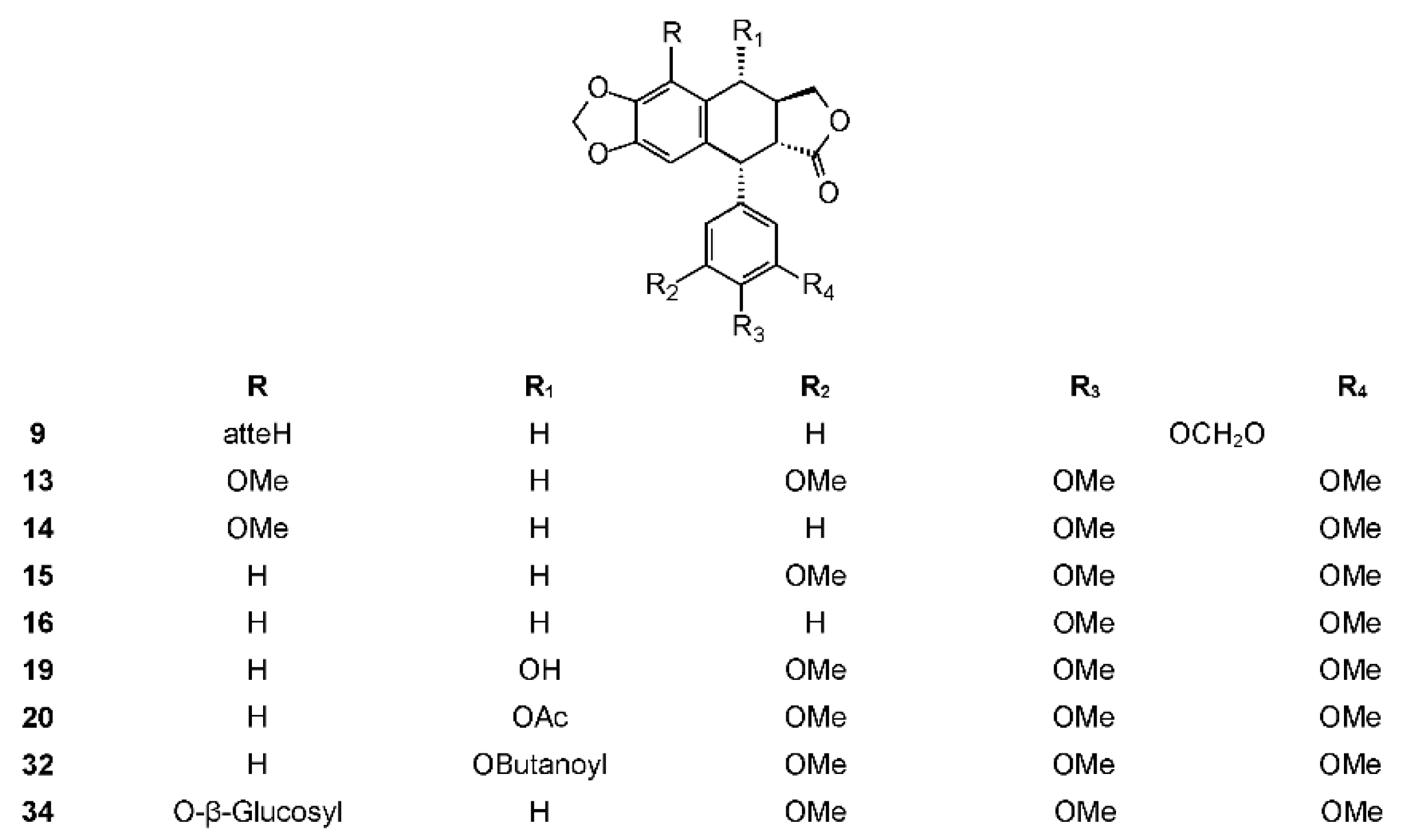
Nieto-Yañez et al. evaluated the anti-leishmanial activity of a B. aptera methanolic extract. The extract showed strong activity against Leishmania mexicana both in the in vitro and in vivo tests. The gas chromatography-mass spectrometry (GC-MS) phytochemical analysis of the extract showed the presence of 11 compounds. Most of these compounds were fatty acids and fatty acid esters, but they revealed the presence of podophyllotoxin (19) (Figure 3) [100].
2.3.2. Bursera arida (Rose) Standl.
B. arida (synonyms: Elaphrium aridum Rose and Terebinthus arida Rose) is endemic in the states of Oaxaca and Puebla [2]. It is commonly known as zapotillo [99]. These are small trees, often shrubs, of reddish-brown exfoliating bark with leaves up to 2.5 cm long and 1 cm wide, comprising 3 to 11 leaflets. Their flowers are very small, reddish, and solitary. Their fruits are solitary or in pairs, over short and pilose peduncles 1 to 2 mm long, with a seed completely covered by a pale yellow pseudoaril [101]. It belongs to the microphylla subsection. Traditionally, the plant latex is topically used for healing wounds and skin eruptions [102] in the Tehuacán-Cuicatlán valley. Bursera arida has different medicinal uses such as a disinfectant, cough suppressant, and antidepressant [103].
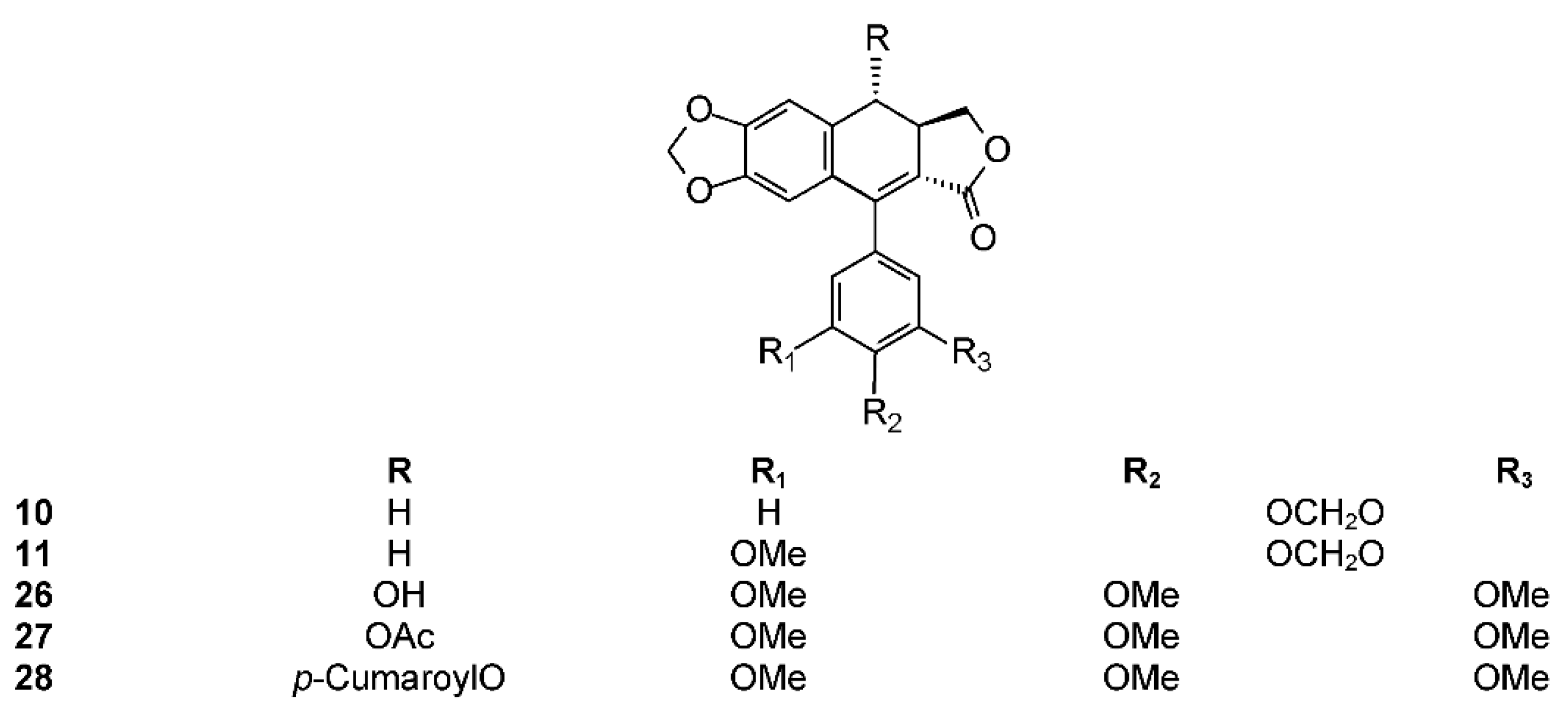
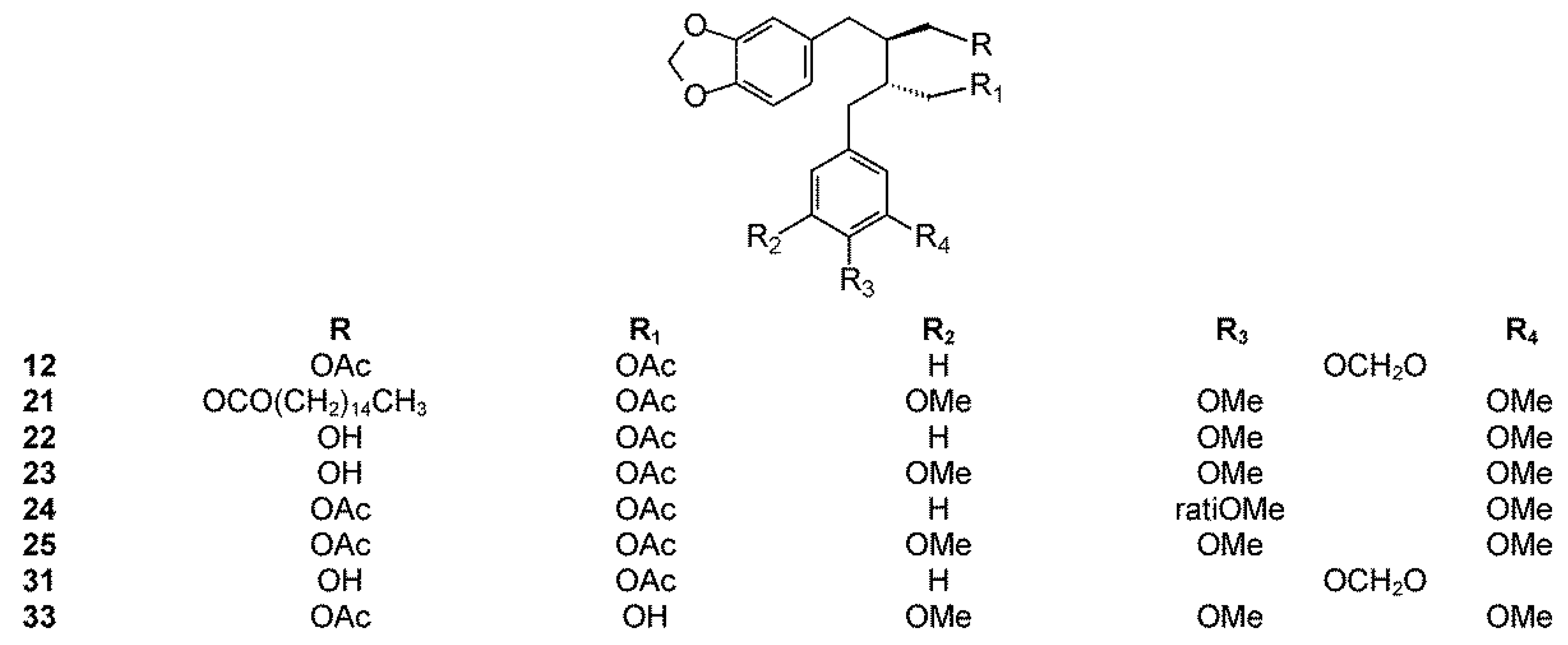
The phytochemistry of B. arida was studied by Ionescu, who prepared a chloroform extract of the stems, leaves, twigs, and bark, and found naringenin, β-sitosterol, betulonic acid, and four lignans: (+)-3-hydroxymethyl-5-methoxy-6,7-methylenedioxy-1-(3′,4′-methylenedioxybenzene)-l,2,3,4-tetrahydronaphthalene-2-carboxylic acid lactone (9) (Figure 3), (+)-3-hydroxymethyl-6,7-methylenedioxy-1-(3′,4′-methylenedioxybenzene)-3,4-dihydronaphthalene-2-carboxylic acid lactone (10) (Figure 4), (+)-3-hydroxymethy1-6,7-methylenedioxy-1-(5′-methoxy-3′,4′-methylenedioxybenzene)-3,4-dihydronaphthalene-2-carboxylic acid lactone (11) (Figure 4), and 2,3-bis-(3,4-methylenedioxybenzyl)butane-l,4-diol diacetate (12) (Figure 5) [104]. Compound 12 has the structure of ariensin.
2.3.3. Bursera ariensis (Kunth) McVaugh and Rzed.
B. ariensis (synonyms B. panosa Engl., B. sessiflora Engl., E. ariensis Kunth, and E. brachypodium Rose). These are trees and sometimes shrubs between 2 and 8 m tall with greenish-gray trunks and bark that exfoliates in papery sheets that are yellowish or beige, sometimes with orange tones, with whitish resin that darkens upon contact with air. Their leaves are hairy especially when young, 5 to 22 cm long, and 2 to 7 cm wide, with a winged rachis and 5 to 9 leaflet pairs. Their flowers can be solitary but often develop in conglomerates at the end of branches of reddish-yellow color. Their fruits are 6 to 8 mm long, growing in thick conglomerates, with a pit completely covered by a yellow or orange pseudoaril. It is distributed in Mexico (Chiapas, Guerrero, Jalisco, and Oaxaca regions) where it is commonly known as guande and cuajiote blanco (Náhuatl name) [101] and used to treat colds and inflammation. It belongs to the fagaroides group and is poorly studied. From the acetonic extract of the bark, Hernandez isolated a new lignan named ariensin (12) [105]. Ariensin was shown to be active against the RAW246.7 murine cell line (IC50 9.8 μM) [11].
2.3.4. Bursera fagaroides (H.B.K.) Engl.
B. fagaroides, or “fragrant bursera” (B. obovata Turcz., B. schaffneri S. Wats) [2] is a dioecious shrub or tree, occasionally hermaphrodite, 0.5 to 10 m high, and highly resinous. The trunk is green with a bark that exfoliates in yellowish-gray papery sheets. The leaves are most often compound, with 5 to 13 leaflets, although occasionally they are unifoliolate or trifoliolate. They have whitish-green or yellow flowers, with a few of them arranged in small inflorescences or solitary. Male flowers are most often 5-merous (sometimes 3- and 4-). Female flowers are 3-merous. Fruits are typically 0.5 to 0.8 cm long with short peduncles no more than 2 mm long that terminate in a sharp point. When the pits mature, they are covered by a yellow or red pseudoaril. It is commonly known as cuajiote amarillo (in Morelos) or pima bajo [106]. This species belongs to the fagaroides group. Three subspecific variants of this species were recognized by McVaugh and Rzedowski (1965). B. fagaroides var. fagaroides, commonly known as xixote and jiote [72], are most often shrubs with leaves with serrated margins and are distributed in Northern, Central, and Western Mexico. B. fagaroides var. purpusii (common name aceitillo) [107] are most often trees with leaves of entire margin (not toothed) distributed throughout Southeastern Mexico. B. fagaroides var. elongata, common Mayo name to’oro sahuali [95], is from Northwestern Mexico, and has been classified by recent molecular studies as a separate taxon not belonging within B. fagaoides [108].
Bursera fagaroides is used to alleviate inflammation, skin tumors, and warts, and is perhaps the most studied Bursera species in terms of its chemistry and biological effects. The first report about B. fagaroides phytochemistry dates back to 1969, when Bianchi et al. isolated β-peltatin A-methyl ether (13) and the new 5′-demethoxy-β-peltatin A-methyl ether (14) (Figure 3) that showed activity against the Walker carcinoma 256 (WA16) tumor system from the chloroform extract of this plant. The ethanol extract from the dried exudate of Bursera fagaroides showed significant cytotoxic activity against the HT-29 (human colon adenocarcinoma) cell line. From this extract, Velazquez-Jimenez et al. isolated two aryltetraline lignans: (−)-deoxypodophyllotoxin (15) and (−)-morelensin (16) (Figure 3), and two dibenzylbutirolactone lignans: (−)-yatein (17) and (−)-5′-desmethoxy yatein (18) or bursehernin] (Figure 1) [109]. The authors determined the absolute configuration of these compounds by comparing the vibrational circular dichroism spectra of known podophyllotoxin and deoxypodophyllotoxin with those obtained by density functional theory calculations. Morelensin (16) was shown to be cytotoxic and antiproliferative against several cancer cell lines [35,110]. Yatein (17) was shown to be cytotoxic and antiproliferative to several cancer cell lines [111,112,113,114]. For example, it was tested against HL-60 (human promyelocytic leukemia cells), SMMC-7721 (human hepatoma), A-549 (human lung adenocarcinoma), MCF-7 (human breast cancer), and SW480 (human colon adenocarcinoma) cell lines using the MTT (3-(4,5-dimethylthiazol-2)2,5-difeniltetrazolium bromide) method as previously reported [115], with cisplatin as the positive control [116]. Yatein (17) showed significant cytotoxic activity against all tested cell lines being superior to cisplatin. Chen et al. tested yatein [117] against DLD-1 (human colorectal carcinoma), CCRF-CEM (human lymphoblastic leukemia), and IMR-32 (human neuroblastoma) cell lines. The study showed that yatein possesses similar cytotoxic activity to doxorubicin (positive control) against DLD-1 and CCRF-CEM cell lines [117]. Other studies showed that it is able to suppress herpes simplex virus type 1 (HSV-1) replication in HeLa cells in a plaque reduction assay. Doussot et al. studied the lignan profile and the antiproliferative activity of ethanol extracts from plants belonging to different species of Linum, Callitris, and Juniperus. They compared the activity of deoxypodophyllotoxin (15), podophyllotoxin (19), and yatein (17) against six human cancer cell lines: A549, U373 (glioblastoma), T98G (glioma), Hs683 (oligodendro-glioma), MCF7 (breast cancer), and SKMEL-28 (melanoma). The most active compound was deoxypodophyllotoxin (IC50 < 0.01 μM against all cell lines, not tested against U373), followed by podophyllotoxin (IC50 = 0.03 μM against all cell lines except U373 (IC50 > 100 μM). Yatein (17) showed an antiproliferative activity, but to a lesser extent (IC50 = 30.9, not tested, 26.5, 29.8, 31.9, and 39.6 μM, respectively) [118]. The inhibitory effect of yatein (17) on HSV-1 replication was concentration-dependent with an IC50 value of 30.6 ± 5.5 μM [119]. Furthermore, yatein (17) was demonstrated to be a potent CYP3A4 inhibitor and this study is of particular importance as 17 and other methylenedioxyphenol compounds were found to induce herb-drug interactions in clinical situations [120]. Yatein (17) showed other important biological activities, such as anti-platelet aggregation [121], and was shown to have moderate inhibitory activity against cytochrome P450 [122]. The bioactivity-guided separation of the hydroalcoholic extract of B. fagaroides var. fagaroides by Rojas-Sepúlveda et al. led to, besides the already isolated lignans, podophyllotoxin (19), burseranin (7), and acetyl podophyllotoxin (20) (Figure 3). All the isolated compounds were found to be active against tumor cell lines tested, especially 5′-demethoxy-β-peltatin A-methyl ether (14), which exhibited greater activity than camptothecin and podophyllotoxin against PC-3 (ED50 = 1.0 × 10−5 μg/mL) and KB (ED50 = 1.0 × 10−5 μg/mL) cell lines [123]. Furthermore, the cytotoxic and antitumor activity of the ethanol extract (70%) of B. fagaroides bark against L5178Y lymphoma cell line was tested. The antitumor activity was studied on BALB/c mice (2 × 104 cells L5178Y i.p). Treated animals (at 50 mg/kg/day over 15 days) showed a significant increase in survival compared with those treated with the placebo or without treatment [124].
Podophyllotoxin and deoxypodophyllotoxin are secondary metabolites of many plants [125]. The biological activities and the importance of podophyllotoxin (19), as the lead compound in the development of new anticancer agents, are well known [126,127,128]. The problem connected with its use is the scarce amount isolated from natural sources. For this reason, biotechnological production of this lignan has been studied [129]. Deoxypodophyllotoxin (15) is a promising anticancer agent [130,131].
Other diarylbutane lignans were isolated by Morales-Serna et al. from the chloroform extract of B. fagaroides resin, named by the author as 9′-acetyl-9-pentadecanoyl-dihydroclusin (21) (correctly, a hexadecanoyl derivative), 2,3-demethoxy-secoisolintetralin monoacetate (22), and dihydroclusin monoacetate (23), together with two known lignans: 2,3-demethoxy-secoisolintetralin diacetate (24) and dihydroclusin diacetate (25) [132] (Figure 5). Recently, Antúnez Mojica et al. isolated three new aryldihydronaphtalene-type lignans from the dichloromethane stem bark extract of B. fagaroides var. fagaroides: 7′,8′-dehydropodophyllotoxin (26), 7′,8′-dehydroacetylpodophyllotoxin (27), and 7′,8′-dehydro-trans-p-cumaroyl podophyllotoxin (28) (Figure 4), along with six known lignans: podophyllotoxin (19), acetylpodophyllotoxin (20), 5′-demethoxy-β-peltatin A methylether (14), acetylpicropodophyllotoxin (29) (Figure 2), burseranin (7), and hinokinin (1) [133]. The cytotoxic activity of the new isolated compounds 26–28 against the cancer cell lines KB, PC-3, MCF-7, and HF-6 was evaluated, which showed that all of them displayed good activity against KB, PC-3, and HF-6, but were not active against the MCF-7 cell line. When compared with podophyllotoxin (19) (ED50 = 2.10 × 10−4 μM), compounds 26 and 28 were most active against the PC-3 cell line displaying similar toxicity (ED50 = 2.4 × 10−5 and 2.42 × 10−5 μM, respectively), whereas compound 27 was less active (ED50 = 0.06 μM). Lignans 26–28 showed moderate activity against KB and HF-6 cell lines when compared to 19. The cytotoxic activity of the other isolated compounds was already proven [123]. Acetylpicropodophyllotoxin (29) was previously isolated from Hernandia ovigera and it is a potent, selective inhibitor of type I insulin-like growth factor receptor (IGF-IR) [134].
Ornithine decarboxylases (ODC) are enzymes that catalyze the decarboxylation of ornithine to produce putrescine in the biosynthesis of polyamines. Polyamine metabolism is closely related with the progression of growth, proliferation, and cell regeneration. The in vitro effect of an ethanolic extract from the stem bark of Bursera fagaroides on ODC activity, and on the growth of Entamoeba histolytica, was studied by Rosas-Arreguín et al. using metronidazole and G418 as positive controls [135]. The authors found growth inhibition, with IC50 values in the order of 0.05 mg/mL. The ODC activity was inhibited by 12% at 4.0 mg/mL.
Gutiérrez-Gutiérrez et al., considering the use in Mexican traditional medicine of B. fagaroides as an antidiarrheic, investigated the in vitro anti-giardial activities of four podophyllotoxin-type lignans from Bursera fagaroides var. fagaroides: 5′-demethoxy-β-peltatin A methylether (14), acetylpodophyllotoxin (20), burseranin (7), and podophyllotoxin (19) [136]. They found that all lignans affected Giardia adhesion, but only compounds 14, 19, and 20 caused growth inhibition.
2.3.5. Bursera microphylla A. Gray
B. microphylla, or “elephant tree” (Elaphrium microphyllum (A. Gray) Rose, Terebinthus microphylla (A. Gray) Rose), is commonly known as xoop (Seri name) and torote blanco [95]. It is typically a small tree that grows up to 10 m tall, with a thickened trunk and thickened lower branches, with light gray to white peeling bark, with younger branches having a reddish color. The leaves are 3 to 8 cm long and have 7 to 35 small, linear, and glabrous leaflets. The leaflets are up to 1.5 cm long. The flowers are yellowish white or greenish and inconspicuous. The fruits are brownish red at maturity with a black pit completely covered by a yellow-orange pseudoaril [26]. It belongs to the microphylla group. The Seri Indians from Sonora, Mexico use the bark, leaves, flowers, and fruits to treat a variety of maladies such as inflammation, diarrhea, and venereal diseases. The first lignan isolated from Bursera microphylla was burseran (30) (Figure 1) [137]. Cole et al. showed that burseran has cytotoxic activity against human epidermoid carcinoma of the nasopharynx (9KB cell line) in a Cancer Chemotherapy National Service Center (CCNSC) test (ED50 < 10 μg/mL) acting as a spindle poison. Tomioka et al. synthetized cis (H8′ α) and trans burseran and tested them in a cilia regeneration test in Tetrahymena, that is a useful model for studying the antitubulinic activity of spindle poisons [138]. Both trans and cis burseran have shown inhibitory activity, but the antitumor activity was higher for trans burseran [139]. An analysis of the chemical composition of the methanol extract of B. microphylla resin from the Sonora Desert (Mexico), revealed the presence of several known lignans: ariensin (12), burseranin (7), dihydroclusin diacetate (25), picropolygamain (8), desmethoxy-yatein (18), hemiariensin (31) (Figure 5), and dihydroclusin 9′-acetate (23); and two new ones: podophyllotoxin butanoate (32) and dihydroclusin 9-acetate (33) (Figure 3 and Figure 5) [11,140]; in addition to burseran (30). Compound 32 was already known as a synthetic derivative of podophyllotoxin [141] but it was new as a natural product. Burseran (30) and dihydroclusin diacetate (25) were tested against human cancer cell lines: A549 (lung cancer), HeLa (cervix cancer), and PC-3 (prostate cancer), and on murine cell lines M12.C3.F6 (B cell lymphoma) and RAW264.7 (macrophages transformed by virus Abelson leukemia); which were found to be more active against murine cell lines in micromolar range (IC50 13.8, 36.3, and 2.5 μM, respectively). The anti-proliferative activities of dihydroclusin 9-acetate (31), dihydroclusin 9′-acetate (23), burseranin (7), picropolygamain (8), and hemiariensin (31) were evaluated on the human cancer cell lines A549, LS 180, and HeLa, and on the human non-cancer cell line ARPE-19. None of the evaluated compounds had statistically significant anti-proliferative effects with respect to dimethyl sulfoxide (DMSO) control on LS 180, A549, and ARPE cell lines. However, burseranin (7) and picropolygamain (8) had an interesting anti-proliferative activity on the gynecological cancer cell line, HeLa, with IC50 values of 21.72 ± 1.03 and 9.31 ± 1.01 µM, respectively [140].
2.3.6. Bursera morelensis Ramírez
B. morelensis (synonym: Elaphrium morelense (Ramírez) Rose) is widely distributed in Mexico [101], where it is commonly known as coabinillo [99]. It is a tree up to 13 m tall with red bark that exfoliates in thin sheets. Its leaves are 5 to 11 cm long and 1.5 to 4.5 cm wide, with 15 to 51 linear leaflets. The flowers are yellow, pink, greenish, or white. The fruits are 0.5 to 1 cm long, with a pit completely covered by a pale yellow pseudoaril. It belongs to the microphylla group. The only two papers describing the phytochemistry of B. morelensis address the composition and the anti-inflammatory activity of the essential oil [142] and the isolation of deoxypodophyllotoxin (15) and morelensin (16) from the resin [110]. Morelensin (16), although highly active against the KB (epidermoid carcinoma) cancer cells, demonstrated only marginal activity against the porcine stable (PS) kidney cell line [110].
2.3.7. Bursera roseana Rzed., Calderón & Medina
B. roseana (synonyms: Bursera acuminata (Rose) Engl. and Terebinthus acuminata Rose) is a 12–20 m high tree with bark peeling in reddish-orange stripes. It is imparipinnate with 3 to 7 (sometimes 9) leaflets with a hairy underside but a bright and glabrous upper side. The leaflets are typically of oval shape, 4.5 to 15 cm long and 2 to 6 cm wide, ending in a long point. The flowers are white or greenish and the is fruit is glabrous, 0.9 to 1.2 cm long, with a pit completely covered by a pale pseudoaril [143].
This species grows in moist canyons in the transition zone between highland pine-oak forest and lowland tropical subdeciduous forest. It is common in Nayarit, Zacatecas, Aguascalientes, Jalisco, Colima, Michoacán, Estado de México, and Guerrero [143]. It belongs to the simaruba group. The only phytochemical study on this species was reported by Koulman, who found 5′-desmethoxy yatein [bursehernin or (−)-trans-methylpluviatolide [144] (+)-trans-methylpluviatolide, which has been called dextrobursehernin [145] (18)], morelensin (16), deoxypodophyllotoxin (15), and β-peltatin-A methylether (13). Bursehernin (18) was studied by Ito et al. for its inhibitory effects on Epstein-Barr virus early antigen activation induced by 12-O-tetradecanoylphorbol 13-acetate in Raji cells [146]. The data reported demonstrated that bursehernin (18) was slightly weaker than β-carotene, which is commonly used in cancer prevention studies [147] so it might be a valuable antitumor promoter. Bursehernin (18) is able to inhibit the growth of Neisseria gonorrhoeae [148]. The trypanocidal activity of racemic mixtures of cis- and trans-bursehernin was evaluated in vitro against trypomastigote forms of two strains of Trypanosoma cruzi, and results showed that the racemic cis-stereoisomer was inactive, whereas the racemic trans-stereoisomer displayed trypanocidal activity, with an IC50 ~89.3 μM. These results were different from those obtained for pure (−)-trans-methylpluviatolide by Bastos et al., but the difference could be ascribed to the use of the racemic mixture [74].
2.3.8. Bursera schlechtendalii Engl.
B. schlechtendalii (synonyms: Bursera jonesii Rose, Elaphrium jonesii (Rose) Rose, Terebinthus jonesii (Rose) Rose, and Terebinthus schlechtendalii (Engl.) Rose) is a small tree or shrub, 4–6 m high, with a strong turpentine smell, and known as sak chakaj. It has a glossy greyish pink bark that peels off in thin papery sheets; the branches are thick and stout. The leaves are simple (unifoliolate), often less than 6 cm long and 2.5 cm wide. The flowers are small, usually solitary, with yellow or reddish petals, and the fruits are 4 to 8 mm long with a pit completely covered by a yellow or red pseudoaril. It is used to treat the flu. It is found at altitudes of 200–400 m on dry rocky hillsides or in thickets in Southern Mexico and Guatemala. It belongs to the fagaroides group. In 1972, McDoniel et al. isolated from stems and leaves the chloroform extract of Mexican B. schlechtendalii Engl. yatein (17) and bursehernin (18).
2.3.9. Bursera simaruba (L.) Sarg.
B. simaruba or “gumbo-limbo” (Bursera simaruba var. yucatanensis Lundell) is commonly known as yala-guito [99]. It is a 6–15 m high tree with a peeling reddish bark that reveals a smooth grey underbark. The leaves are compound, bright, and mostly glabrous when mature. Leaves have 3 to 13 leaflets, 4 to 9 cm long and 1.8 to 3.5 cm wide. The small flowers have pink, pale yellow-green, or white petals and are arranged in inflorescences. The fruits are glabrous, red to brownish, 1 to 1.5 cm long, ending in a point, with a pit completely covered by a red pseudoaril. This is perhaps the most widespread species of Bursera, occurring from Southern Florida and the Caribbean, along both coasts of Mexico, to South America. Taken orally or as curative baths, the leaves and bark are attributed a variety of medicinal properties. It belongs to the simaruba group. In 1992, Peraza-Sánchez and Peña-Rodriguez isolated picropolygamain (8), which showed activity in the brine shrimp assay (LC50 = 52.2 ppm). Further in vitro evaluation against three human tumor cell lines (A-549, lung), MCF-7 (breast), and HT-29 (colon) showed that 8 has cytotoxic activity comparable to that of Adriamycin [93]. Noguera et al. isolated the anti-inflammatory β-peltatin A-methyl ether (13) from the leaf hexane extract of B. simaruba. It inhibited the carrageenan-induced rat paw edema, in a dose- and time-dependent manner (three hours = 9.55%, five hours = 34.37%, and seven hours = 35.6%) [149]. Maldini et al. studied the methanolic extract of Bursera simaruba bark and isolated 11 compounds, including lignans yatein (17), β-peltatin-O-β-d-glucopyranoside (34) (Figure 3), hinokinin (1), and bursehernin (18) [149].
3. Conclusions
Lignans are phenolic secondary metabolites characterized by a large variety of biological activities. Among these, the cytotoxic and anti-proliferative ones are perhaps the most common and most studied. Literature analysis of Mexican plants producing lignans Burseara spp., revealed that the most common lignan types are dibenzyl butyrolactones (nine compounds), picro aryltetraline derivatives (three compounds), aryltetraline derivatives (nine compounds), 7′,8′-dehydro-aryltetraline derivatives (five compounds), and dibenzylbutane diols (eight compounds). Notably, all the examined Elaphrium subgenus species produce only dibenzyl butyrolactones and picro aryltetraline derivatives, whereas Bursera subgenus produce all lignan types. The most common compound, up to now, appears to be bursehernin (18), which is present in five species belonging to the Bursera section. Hinokinin (1) and savinin (2) are also widespread (four species), both in the Elaphrium and Bursera sections. Compounds 3, 8, 12, 13, 15, 16, 17, and 33 were isolated from three species.
Author Contributions
M.C.M. and J.X.B. contributed to the conception of the review; M.C. contributed to the constructive discussions; all authors have read and approved the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Becerra, J.X.; Noge, K.; Olivier, S.; Venable, D.L. The monophyly of Bursera and its impact for divergence times of Burseraceae. Taxon 2012, 61, 333–343. [Google Scholar]
- Rzedowski, J.; Medina Lemos, R.; Calderón de Rzedowski, G. Inventario del conocimiento taxonómico, así como de la diversidad y del endemismo regionales de las especies mexicanas de Bursera (Burseraceae). Acta Bot. Mex. 2005, 70, 75–111. [Google Scholar] [CrossRef]
- Becerra, J.X.; Venable, D.L.; Evans, P.H.; Bowers, W.S. Interactions between chemical and mechanical defenses in the plant genus Bursera and their implications for herbivores. Am. Zool. 2001, 41, 865–876. [Google Scholar]
- Gigliarelli, G.; Becerra, J.X.; Curini, M.; Marcotullio, M.C. Chemical composition and biological activities of fragrant Mexican copal (Bursera spp.). Molecules 2015, 20, 22383–22394. [Google Scholar] [CrossRef] [PubMed]
- Becerra, J.X. Evolution of Mexican Bursera (Burseraceae) inferred from ITS, ETS, and 5S nuclear ribosomal DNA sequences. Mol. Phylogenet. Evol. 2003, 26, 300–309. [Google Scholar] [CrossRef]
- Becerra, J.X.; Noge, K.; Venable, D.L. Macroevolutionary chemical escalation in an ancient plant-herbivore arms race. Proc. Natl. Acad. Sci. USA 2009, 106, 18062–18066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, P.H.; Becerra, J.X. Non-terpenoid essential oils from Bursera chemapodicta. Flavour Fragr. J. 2006, 21, 616–618. [Google Scholar] [CrossRef]
- Noge, K.; Becerra, J.X. Germacrene D, a Common Sesquiterpene in the Genus Bursera (Burseraceae). Molecules 2009, 14, 5289–5297. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Hernandez, J.D.; Garcia-Gutierrez, H.A.; Roman-Marin, L.U.; Torres-Blanco, Y.I.; Cerda-Garcia-Rojas, C.M.; Joseph-Nathan, P. Absolute configuration of cembrane diterpenoids from Bursera multijuga. Nat. Prod. Commun. 2014, 9, 1249–1252. [Google Scholar] [PubMed]
- Hernández-Hernández, J.D.; Román-Marín, L.U.; Cerda-García-Rojas, C.M.; Joseph-Nathan, P. Verticillane Derivatives from Bursera suntui and Bursera kerberi. J. Nat. Prod. 2005, 68, 1598–1602. [Google Scholar] [CrossRef] [PubMed]
- Messina, F.; Curini, M.; Di Sano, C.; Zadra, C.; Gigliarelli, G.; Rascon-Valenzuela, L.A.; Robles Zepeda, R.E.; Marcotullio, M.C. Diterpenoids and triterpenoids from the resin of Bursera microphylla and their cytotoxic activity. J. Nat. Prod. 2015, 78, 1184–1188. [Google Scholar] [CrossRef] [PubMed]
- Lucero-Gómez, P.; Mathe, C.; Vieillescazes, C.; Bucio, L.; Belio, I.; Vega, R. Analysis of Mexican reference standards for Bursera spp. resins by Gas Chromatography–Mass Spectrometry and application to archaeological objects. J. Archaeol. Sci. 2014, 41, 679–690. [Google Scholar] [CrossRef]
- Hernandez, J.D.; Garcia, L.; Hernandez, A.; Alvarez, R.; Roman, L.U. Luteolin and myricetin glycosides of Burseraceae. Rev. Soc. Quim. Mex. 2002, 46, 295–300. [Google Scholar]
- Nakanishi, T.; Inatomi, Y.; Arai, S.; Yamada, T.; Fukatsu, H.; Murata, H.; Inada, A.; Matsuura, N.; Ubukata, M.; Murata, J.; et al. New luteolin 3′-O-acylated rhamnosides from leaves of Bursera graveolens. Heterocycles 2003, 60, 2077–2083. [Google Scholar]
- Souza, M.P.; Machado, M.I.L.; Braz-Filho, R. Six flavonoids from Bursera leptophloeos. Phytochemistry 1989, 28, 2467–2470. [Google Scholar] [CrossRef]
- Cunha, W.R.; Andrade e Silva, M.L.; Sola Veneziani, R.C.; Ambrósio, S.R.; Kenupp Bastos, J. Lignans: Chemical and biological properties. In Phytochemicals-A Global Perspective of Their Role in Nutrition and Health; Rao, V., Ed.; InTech: Vienna, Austria, 2012. [Google Scholar] [CrossRef]
- The Plant List A Working List of All Plant Species. Available online: http://www.theplantlist.org/1.1/browse/A/Burseraceae/Bursera/ (accessed on 12 February 2015).
- Medina Lemos, R.; Ramos Rivera, P. El Género Bursera en México. Parte II; Version 1.3; Comisión Nacional Para el Conocimiento y uso de la Biodiversidad: Mexico City, Mexico, 2017. [Google Scholar]
- CONABIO Enciclovida CONABIO. Available online: http://www.enciclovida.mx/busquedas/resultados?utf8=%E2%9C%93&busqueda=basica&id=&nombre=bursera&button= (accessed on 29 July 2018).
- Bullock, L.L. Notes on the Mexican species of the genus Bursera. Bull. Misc. Inf. Kew 1936, 347–387. [Google Scholar]
- McVaugh, R.; Rzedowski, J. Synopsis of the genus Bursera L. in western Mexico, with notes on the material of Bursera collected by Sessè & Mocino. Kew Bull. 1965, 18, 317–382. [Google Scholar]
- Becerra, J.X.; Venable, D.L. Nuclear ribosomal DNA phylogeny and its implications for evolutionary trends in Mexican Bursera (Burseraceae). Am. J. Bot. 1999, 86, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Rzedowki, J. Notas sobre el genero Bursera (Burseraceae) en el Estado de Guerrero, (Mexico). Am. Esc Nac Cienc. Biol. Mex. 1968, 7, 17–36. [Google Scholar]
- Toledo, C.A. El Genero Bursera en el Estado Guerrero; Universidad Nacional Autónoma de México: Mexico City, Mexico, 1982. [Google Scholar]
- Gillet, J. Commiphora (Burseraceae) in South America and its relationship to Bursera. Kew Bull. 1980, 34, 569–587. [Google Scholar] [CrossRef]
- Rzedowki, J.; Kruse, H. Algunas tendencias evolutivas en Bursera (Burseraceae). Taxon 1979, 28, 103–116. [Google Scholar] [CrossRef]
- Hernández, F. Historia de las Plantas de Nueva España; Imprenta Universitaria: Mexico City, México, 1942. [Google Scholar]
- Tripplett, K.J. The Ethnobotany of Plant Resins in the Maya Cultural Region of Southern Mexico and Central America; University of Texas: Austin, TX, USA, 1999. [Google Scholar]
- Acevedo, M.; Nuñez, P.; Gónzalez-Maya, L.; CardosoTaketa, A.; Villarreal, M.L. Cytotoxic and anti-inflammatory activities of Bursera species from Mexico. J. Clin. Toxicol. 2015, 5, 1–8. [Google Scholar]
- Alonso-Castro, A.J.; Villarreal, M.L.; Salazar-Olivo, L.A.; Gomez-Sanchez, M.; Dominguez, F.; Garcia-Carranca, A. Mexican medicinal plants used for cancer treatment: Pharmacological, phytochemical and ethnobotanical studies. J. Ethnopharmacol. 2011, 133, 945–972. [Google Scholar] [CrossRef] [PubMed]
- CONABIO Biodiversidad Mexicana. Available online: http://www.biodiversidad.gob.mx/usos/copales/almarciga.html (accessed on 8 May 2018).
- Waizel, J.; Waizel-Hayat, S. Antitussive plants used in Mexican traditional medicine. Pharmacogn. Rev. 2009, 3, 29–43. [Google Scholar]
- Koulman, A. Podophyllotoxin: A Study of the Biosynthesis, Evolution, Function and Use of Podophyllotoxin and Related Lignans; University of Groningen: Groningen, The Netherlands, 2003. [Google Scholar]
- Marcotullio, M.C.; Pelosi, A.; Curini, M. Hinokinin, an emerging bioactive lignan. Molecules 2014, 19, 14862–14878. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, R.; Nagao, T.; Okabe, H.; Nakano, Y.; Matsunaga, H.; Katano, M.; Mori, M. Antiproliferative constituents in Umbelliferae plants. IV. Constituents in the fruits of Anthriscus sylvestris Hoffm. Chem. Pharm. Bull. 1998, 46, 875–878. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T.; Wang, D.S.Y.; Wu, C.L.; Shiah, S.G.; Kuo, Y.H.; Chang, C.J. Cytotoxicity of extractives from Taiwania cryptomerioides heartwood. Phytochemistry 2000, 55, 227–232. [Google Scholar] [CrossRef]
- Chen, J.-J.; Huang, H.-Y.; Duh, C.-Y.; Chen, I.-S. Cytotoxic constituents from the stem bark of Zanthoxylum pistaciiflorum. J. Chin. Chem. Soc. 2004, 51, 659–663. [Google Scholar] [CrossRef]
- Lin, R.-W.; Tsai, I.-L.; Duh, C.-Y.; Lee, K.-H.; Chen, I.-S. New lignans and cytotoxic constituents from Wikstroemia lanceolata. Planta Med. 2004, 70, 234–238. [Google Scholar] [PubMed]
- Chen, J.-J.; Chung, C.-Y.; Hwang, T.-L.; Chen, J.-F. Amides and benzenoids from Zanthoxylum ailanthoides with inhibitory activity on superoxide generation and elastase release by neutrophils. J. Nat. Prod. 2009, 72, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-Y.; Seo, K.-H.; Jeong, R.-H.; Lee, S.-M.; Kim, G.-S.; Noh, H.-J.; Kim, S.-Y.; Kim, G.-W.; Kim, J.-Y.; Baek, N.-I. Anti-inflammatory lignans from the fruits of Acanthopanax sessiliflorus. Molecules 2013, 18, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.; de Souza, G.H.B.; da Silva, A.A.; de Souza, V.A.; Pereira, A.C.; Royo, V.D.A.; e Silva, M.L.A.; Donate, P.M.; de Matos Araujo, A.L.S.; Carvalho, J.C.T.; et al. Synthesis and biological activity evaluation of lignan lactones derived from (−)-cubebin. Bioorg. Med. Chem. Lett. 2005, 15, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.S.C.; Lucarini, R.; Volpe, A.C.; de Andrade, C.Q.J.; Souza, A.M.P.; Pauletti, P.M.; Januário, A.H.; Símaro, G.V.; Bastos, J.K.; Cunha, W.R.; et al. In vivo and in silico anti-inflammatory mechanism of action of the semisynthetic (−)-cubebin derivatives (−)-hinokinin and (−)-O-benzylcubebin. Bioorg. Med. Chem. Lett. 2017, 27, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Ramos, F.; Takaishi, Y.; Kawazoe, K.; Osorio, C.; Duque, C.; Acuna, R.; Fujimoto, Y.; Sato, M.; Okamoto, M.; Oshikawa, T.; et al. Immunosuppressive diacetylenes, ceramides and cerebrosides from Hydrocotyle leucocephala. Phytochemistry 2006, 67, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yao, Z.; Zhang, Y.W.; Zhang, X.X.; Takaishi, Y.; Duan, H.Q. Immunosuppressive sesquiterpenes from Buddleja daviddi. Planta Med. 2010, 76, 1882–1887. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.L.A.; Coimbra, H.S.; Pereira, A.C.; Almeida, V.A.; Lima, T.C.; Costa, E.S.; Vinholis, A.H.C.; Royo, V.A.; Silva, R.; Filho, A.A.S.; et al. Evaluation of Piper cubeba extract, (−)-cubebin and its semi-synthetic derivatives against oral pathogens. Phytoth. Res. 2007, 21, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.-L.; Huang, Y.-L.; Ou, J.-C.; Chen, C.-C.; Hsu, F.-L.; Chang, C. Screening of 25 compounds isolated from Phyllanthus species for anti-human Hepatitis B virus in vitro. Phytother. Res. 2003, 17, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Shimokawa, S.; Mochizuki, M.; Kumamoto, T.; Nakanishi, W.; Watanabe, T.; Ishikawa, T.; Matsumoto, K.; Tashima, K.; Horie, S.; et al. Chemical constituents of Aristolochia constricta: Antispasmodic effects of its constituents in guinea-pig ileum and isolation of a diterpeno-lignan hybrid. J. Nat. Prod. 2008, 71, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Kuroyanagi, M.; Ikeda, R.; Gao, H.Y.; Muto, N.; Otaki, K.; Sano, T.; Kawahara, N.; Nakane, T. Neurite outgrowth-promoting active constituents of the Japanese cypress (Chamaecyparis obtusa). Chem. Pharm. Bull. 2008, 56, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Adfa, M.; Rahmad, R.; Ninomiya, M.; Yudha, S.; Tanaka, K.; Koketsu, M. Antileukemic activity of lignans and phenylpropanoids of Cinnamomum parthenoxylon. Bioorg. Med. Chem. Lett. 2016, 26, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Cunha, N.L.; Teixeira, G.M.; Martins, T.S.D.; Souza, A.R.; Oliveira, P.F.; Símaro, G.V.N.; Rezende, K.C.S.; Gonçalves, N.L.D.S.; Souza, D.G.; Tavares, D.C.; et al. (−)-Hinokinin induces G2/M arrest and contributes to the antiproliferative effects of doxorubicin in breast cancer cells. Planta Med. 2016, 82, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Koo, K.A.; Ma, C.J.; Sung, S.H.; Kim, Y.C. Neuroprotective lignans from Biota orientalis leaves. Nat. Prod. Sci. 2008, 14, 167–170. [Google Scholar]
- De Souza, V.A.; da Silva, R.; Pereira, A.C.; Royo Vde, A.; Saraiva, J.; Montanheiro, M.; de Souza, G.H.; da Silva Filho, A.A.; Grando, M.D.; Donate, P.M.; et al. Trypanocidal activity of (−)-cubebin derivatives against free amastigote forms of Trypanosoma cruzi. Bioorg. Med. Chem. Lett. 2005, 15, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Esperandim, V.R.; da Silva Ferreira, D.; Saraiva, J.; Silva, M.L.A.; Costa, E.S.; Pereira, A.C.; Bastos, J.K.; de Albuquerque, S. Reduction of parasitism tissue by treatment of mice chronically infected with Trypanosoma cruzi with lignano lactones. Parasitol. Res. 2010, 107, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Esperandim, V.R.; da Silva Ferreira, D.; Rezende, K.C.; Cunha, W.R.; Saraiva, J.; Bastos, J.K.; e Silva, M.L.; de Albuquerque, S. Evaluation of the in vivo therapeutic properties of (−)-cubebin and (−)-hinokinin against Trypanosoma cruzi. Experim. Parasitol. 2013, 133, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, J.; Vega, C.; Rolon, M.; da Silva, R.; Andrade, E.S.M.L.; Donate, P.M.; Bastos, J.K.; Gomez-Barrio, A.; de, A.S. In vitro and in vivo activity of lignan lactones derivatives against Trypanosoma cruzi. Parasitol. Res. 2007, 100, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, J.; Lira, A.A.M.; Esperandim, V.R.; da Silva Ferreira, D.; Ferraudo, A.S.; Bastos, J.K.; e Silva, M.L.A.; de Gaitani, C.M.; de Albuquerque, S.; Marchetti, J.M. (−)-Hinokinin-loaded poly(D,L-lactide-co-glycolide) microparticles for Chagas disease. Parasitol. Res. 2010, 106, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Sartorelli, P.; Carvalho, C.S.; Reimao, J.Q.; Lorenzi, H.; Tempone, A.G. Antitrypanosomal activity of a diterpene and lignans isolated from Aristolochia cymbifera. Planta Med. 2010, 76, 1454–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haribabu, K.; Ajitha, M.; Mallavadhani, U.V. Quantitative estimation of (−)-hinokinin, a trypanosomicidal marker in Piper cubeba, and some of its commercial formulations using HPLC-PDA. J. Pharm. Anal. 2015, 5, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Andrade e Silva, M.L.; Cicarelli, R.M.; Pauletti, P.M.; Luz, P.P.; Rezende, K.C.; Januario, A.H.; da Silva, R.; Pereira, A.C.; Bastos, J.K.; de Albuquerque, S.; et al. Trypanosoma cruzi: Evaluation of (−)-cubebin derivatives activity in the messenger RNAs processing. Parasitol. Res. 2011, 109, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Resende, F.A.; Tomazella, I.M.; Barbosa, L.C.; Ponce, M.; Furtado, R.A.; Pereira, A.C.; Bastos, J.K.; Andrade, E.S.M.L.; Tavares, D.C. Effect of the dibenzylbutyrolactone lignan (−)-hinokinin on doxorubicin and methyl methanesulfonate clastogenicity in V79 Chinese hamster lung fibroblasts. Mutat. Res. 2010, 700, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Resende, F.A.; Barbosa, L.C.; Tavares, D.C.; de Camargo, M.S.; de Souza Rezende, K.C.; ML, E.S.; Varanda, E.A. Mutagenicity and antimutagenicity of (−)-hinokinin a trypanosomicidal compound measured by Salmonella microsome and comet assays. BMC Complement. Altern. Med. 2012, 12, 203. [Google Scholar] [CrossRef] [PubMed]
- Masumura, M.; Okumura, F.S. Identity of hibalactone and savinin. J. Am. Chem. Soc. 1955, 77, 1906. [Google Scholar] [CrossRef]
- Hartwell, J.L.; Johnson, J.M.; Fitzgerald, D.B.; Belkin, M. Podophyllotoxin from Juniperus species; savinin. J. Am. Chem. Soc. 1953, 75, 235–236. [Google Scholar] [CrossRef]
- Woo, K.W.; Choi, S.U.; Park, J.C.; Lee, K.R. A new lignan glycoside from Juniperus rigida. Arch. Pharm. Res. 2011, 34, 2043–2049. [Google Scholar] [CrossRef] [PubMed]
- Badawi, M.M.; Seida, A.A.; Kinghorn, A.D.; Cordell, G.A.; Farnsworth, N.R. Potential anticancer agents. XVIII. Constituents of Amyris pinnata (Rutaceae). J. Nat. Prod. 1981, 44, 331–334. [Google Scholar] [CrossRef]
- Mansoor, T.A.; Borralho, P.M.; Luo, X.; Mulhovo, S.; Rodrigues, C.M.P.; Ferreira, M.-J.U. Apoptosis inducing activity of benzophenanthridine-type alkaloids and 2-arylbenzofuran neolignans in HCT116 colon carcinoma cells. Phytomedicine 2013, 20, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Park, J.; Kim, P.S.; Yoo, E.S.; Baik, K.U.; Park, M.H. Savinin, a lignan from Pterocarpus santalinus inhibits tumor necrosis factor-α production and T cell proliferation. Biol. Pharm. Bull. 2001, 24, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ban, H.S.; Kim, Y.P.; Kim, B.-K.; Cho, S.H.; Ohuchi, K.; Shin, K.H. Lignans from Acanthopanax chiisanensis having an inhibitory activity on prostaglandin E2 production. Phytother. Res. 2005, 19, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Mbaze, L.M.A.; Lado, J.A.; Wansi, J.D.; Shiao, T.C.; Chiozem, D.D.; Mesaik, M.A.; Choudhary, M.I.; Lacaille-Dubois, M.-A.; Wandji, J.; Roy, R.; et al. Oxidative burst inhibitory and cytotoxic amides and lignans from the stem bark of Fagara heitzii (Rutaceae). Phytochemistry 2009, 70, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.L.S.; Leite, K.C.D.S.; de Macêdo, I.Y.L.; de Morais, S.R.; Costa, E.A.; de Paula, J.R.; de Souza Gil, E. Electrochemical behavior and antioxidant activity of hibalactone. Int. J. Electrochem. Sci. 2017, 12, 7956–7964. [Google Scholar] [CrossRef]
- Oliveira, T.L.S.; Morais, S.R.; Sá, S.; Oliveira, M.G.; Florentino, I.F.; Silva, D.M.D.; Carvalho, V.V.; Silva, V.B.D.; Vaz, B.G.; Sabino, J.R.; et al. Antinociceptive, anti-inflammatory and anxiolytic-like effects of the ethanolic extract, fractions and Hibalactone isolated from Hydrocotyle umbellata L. (Acaricoba)—Araliaceae. Biomed. Pharmacother. 2017, 95, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Rzedowski, J.; Guevara-Féfer, F. Flora del Bajio y de Regiones Adyacentes; Instituto de Ecologia: Xalapa, Mexico, 1992; Volume 3. [Google Scholar]
- Chatterjee, A.; Basa, S.C.; Ray, A.B. Spectral properties of the lignin cubebin. J. Indian Chem. Soc. 1968, 45, 723–725. [Google Scholar]
- Bastos, J.K.; Albuquerque, S.; Silva, M.L.A. Evaluation of the trypanocidal activity of lignans isolated from the leaves of Zanthoxylum naranjillo. Planta Med. 1999, 65, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.T.M.; Rezende, K.C.S.; Evora, P.R.B.; Bastos, J.K.; Cunha, W.R.; Andrade e Silva, M.L.; Celotto, A.C. The lignan (−)-Cubebin inhibits vascular contraction and induces relaxation via nitric oxide activation in isolated rat aorta. Phytother. Res. 2013, 27, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Niwa, A.M.; Marcarini, J.C.; Sartori, D.; Maistro, E.L.; Mantovani, M.S. Effects of (−)-cubebin (Piper cubeba) on cytotoxicity, mutagenicity and expression of p38 MAP kinase and GSTa2 in a hepatoma cell line. J. Food Compos. Anal. 2013, 30, 1–5. [Google Scholar] [CrossRef]
- De Rezende, A.A.A.; Silva, M.L.A.; Tavares, D.C.; Cunha, W.R.; Rezende, K.C.S.; Bastos, J.K.; Lehmann, M.; de Andrade, H.H.R.; Guterres, Z.R.; Silva, L.P.; et al. The effect of the dibenzylbutyrolactolic lignan (−)-cubebin on doxorubicin mutagenicity and recombinogenicity in wing somatic cells of Drosophila melanogaster. Food Chem. Toxicol. 2011, 49, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha Pissurno, A.P.; De Laurentiz, R.D.S. Cubebin: A small molecule with great potential. Rev. Virtual Quim. 2017, 9, 656–671. [Google Scholar] [CrossRef]
- Cook, S. The Forest of the Lacandon Maya: An Ethnobotanical Guide; Springer: New York, NY, USA, 2016. [Google Scholar]
- Takaoka, D.; Imooka, M.; Hiroi, M. Studies of lignoids in Lauraceae. III. A new lignan from the heart wood of Cinnamomum camphora Sieb. Bull. Chem. Soc. Jpn. 1977, 50, 2821–2822. [Google Scholar] [CrossRef]
- Lopes, L.M.X.; Yoshida, M.; Gottlieb, O.R. The chemistry of Brazilian Myristicaceae. Part XIX. Dibenzylbutyrolactone lignans from Virola sebifera. Phytochemistry 1983, 22, 1516–1518. [Google Scholar] [CrossRef]
- Kato, M.J.; Yoshida, M.; Gottlieb, O.R. Lignoids and arylalkanones from fruits of Virola elongata. Phytochemistry 1990, 29, 1799–1810. [Google Scholar] [CrossRef]
- Gozler, T.; Gozler, B.; Patra, A.; Leet, J.E.; Freyer, A.J.; Shamma, M. Konyanin: A new lignan from Haplophyllum vulcanicum. Tetrahedron 1984, 40, 1145–1150. [Google Scholar] [CrossRef]
- Evcim, U.; Gozler, B.; Freyer, A.J.; Shamma, M. Haplomyrtin and (−)-haplomyrfolin: Two lignans from Haplophyllum myrtifolium. Phytochemistry 1986, 25, 1949–1951. [Google Scholar] [CrossRef]
- Sheriha, G.M.; Abouamer, K.; Elshtaiwi, B.Z.; Ashour, A.S.; Abed, F.A.; Alhallaq, H.H. Quinoline alkaloids and cytotoxic lignans from Haplophyllum tuberculatum. Phytochemistry 1987, 26, 3339–3341. [Google Scholar] [CrossRef]
- Rifai, Y. A new method for fast isolation of GLI inhibitory compounds. Int. J. Pharma Res. Rev. 2012, 1, 28–30. [Google Scholar]
- Ogungbe, I.V.; Erwin, W.R.; Setzer, W.N. Antileishmanial phytochemical phenolics: Molecular docking to potential protein targets. J. Mol. Gr. Modell. 2014, 48, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Sriwiriyajan, S.; Sukpondma, Y.; Srisawat, T.; Madla, S.; Graidist, P. (−)-Kusunokinin and piperloguminine from Piper nigrum: An alternative option to treat breast cancer. Biomed. Pharmacother. 2017, 92, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.G.; Estevez-Reyes, R.; Mato, C.; Estevez-Braun, A.M. Three lignans from Bupleurum salicifolium. Phytochemistry 1990, 29, 1981–1983. [Google Scholar] [CrossRef]
- Espinosa Organista, D.; Ramos Rivera, P.; Careaga Olvera, S.A. El Género Bursera en México. Parte II, 1.3 ed.; CONABIO: México City, México, 2017. [Google Scholar]
- Nakanishi, T.; Inatomi, Y.; Murata, H.; Shigeta, K.; Iida, N.; Inada, A.; Murata, J.; Farrera, M.A.P.; Iinuma, M.; Tanaka, T.; et al. A new and known cytotoxic aryltetralin-type lignans from stems of Bursera graveolens. Chem. Pharm. Bull. 2005, 53, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Provan, G.J.; Waterman, P.G. Picropolygamain: A new lignan from Commiphora incisa resin. Planta Med. 1985, 51, 271–272. [Google Scholar] [CrossRef]
- Peraza-Sanchez, S.R.; Pena-Rodriguez, L.M. Isolation of picropolygamain from the resin of Bursera simaruba. J. Nat. Prod. 1992, 55, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Mi, Q.; Pezzuto, J.M.; Farnsworth, N.R.; Wani, M.C.; Kinghorn, A.D.; Swanson, S.M. Use of the in Vivo Hollow Fiber Assay in Natural Products Anticancer Drug Discovery. J. Nat. Prod. 2009, 72, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Felger, R.S.; Johnson, M.B.; Wilson, M.F. The Trees of Sonora, Mexico; Oxford University Press: New York, NY, USA, 2001. [Google Scholar]
- Gentry, H.S. Rio Mayo Plants; Carnegie Institution: Washington, DC, USA, 1942. [Google Scholar]
- Garcia Martinez, L.E. Aspectos Socio-Ecologicos Para el Manejo Sustenable del Coal en el Ejido de Acateyahualco, Gro; UNAM: Morelia, Mexico, 2012. [Google Scholar]
- Montúfar, A. Los Copales Mexicanos y la Resina Sagrada del Templo Mayor de Tenochtitlan; Instituto Nacional de Antropología e Historia: Mexico City, Mexico, 2007.
- Dávila Aranda, P.; Ramos Rivera, P. La Flora útil de dos Comunidades Indígenas del Valle de Tehuacán-Cuicatlán: Coxcatlán y Zapotitlán de las Salinas, Puebla; Version 1.3; Comisión Nacional Para el Conocimiento y uso de la Biodiversidad: Mexico City, Mexico, 2017.
- Nieto-Yañez, O.J.; Resendiz-Albor, A.A.; Ruiz-Hurtado, P.A.; Rivera-Yañez, N.; Rodriguez-Canales, M.; Rodriguez-Sosa, M.; Juarez-Avelar, I.; Rodriguez-Lopez, M.G.; Canales-Martinez, M.M.; Rodriguez-Monroy, M.A. In vivo and in vitro antileishmanial effects of methanolic extract from bark of Bursera aptera. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Rzedowski, J.; Medina Lemos, R.; Calderón de Rzedowski, G. Las especies de Bursera (Burseraceae) en la cuenca superior del rio Papaloapan (Mexico). Acta Bot. Mex. 2004, 66, 23–151. [Google Scholar] [CrossRef]
- Canales, M.; Hernández, T.; Caballero, J.; Vivar, A.R.D.; Avila, G.; Duran, A.; Lira, R. Informant consensus factor and antibacterial activity of the medicinal plants used by the people of San Rafael Coxcatlán, Puebla, México. J. Ethnopharmacol. 2005, 97, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Gorgua Jiménez, G.; Nieto Yañez, O.; Ruiz Hurtado, P.A.; Rivera Yáñez, N.; Rodríguez Monroy, M.A.; Canales Martínez, M.M. Bursera arida. Evaluacion de su actividad antiparasitaria y caracterizacion fitoquimica del extracto metanolico. In 11 Reunion Internacional de Investigacion en Productos Naturales; Navarrete, A., Ed.; Revista Latinoamericana de Quimica: San Carlos, Mexico, 2015. [Google Scholar]
- Ionescu, F. Phytochemical Investigation of Bursera Arida Family Burseraceae; The University of Arizona: Tucson, Arizona, 1974. [Google Scholar]
- Hernández, J.D.; Román, L.U.; Espiñeira, J.; Joseph-Nathan, P. Ariensin, a New Lignan from Bursera ariensis. Planta Med. 1983, 47, 215–217. [Google Scholar] [CrossRef] [PubMed]
- LLIFLE Encyclopedias of Living Form. Available online: http://www.llifle.com/Encyclopedia/ (accessed on 5 August 2018).
- Avendaño Reyes, S.; Acosta Rosado, I. Plantas utilizadas como cercas vivas en el estado de Veracruz. Madera Bosques 2000, 6, 55–71. [Google Scholar] [CrossRef]
- Velazquez-Jimenez, R.; Torres-Valencia, J.M.; Cerda-Garcia-Rojas, C.M.; Hernandez-Hernandez, J.D.; Roman-Marin, L.U.; Manriquez-Torres, J.J.; Gomez-Hurtado, M.A.; Valdez-Calderon, A.; Motilva, V.; Garcia-Maurino, S.; et al. Absolute configuration of podophyllotoxin related lignans from Bursera fagaroides using vibrational circular dichroism. Phytochemistry 2011, 72, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Jolad, S.D.; Wiedhopf, R.M.; Cole, J.R. Cytotoxic agents from Bursera morelensis (Burseraceae): Deoxypodophyllotoxin and a new lignan, 5′-desmethoxydeoxypodophyllotoxin. J. Pharm. Sci. 1977, 66, 892–893. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Ishikawa, T.; Duh, C.Y.; Tsai, I.L.; Chen, I.S. New dimeric aporphine alkaloids and cytotoxic constituents of Hernandia nymphaeifolia. Planta Med. 1996, 62, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Novelo, M.; Cruz, J.G.; Hernández, L.; Pereda-Miranda, R.; Chai, H.; Mar, W.; Pezzuto, J.M. Cytotoxic Constituents from Hyptis verticillata. J. Nat. Prod. 1993, 56, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Fierro, C.S.; Tiezzi, A.; Ovidi, E.; Ceccarelli, D.; Triggiani, D.; Mastrogiovanni, F.; Taddei, A.R.; Pérez, C.; Becerra, J.; Silva, M.; Passarella, D. Antiproliferative activity of yatein isolated from Austrocedrus chilensis against murine myeloma cells: Cytological studies and chemical investigations. Pharm. Biol. 2015, 53, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Young, K.; Rakotondraibe, L.H.; Brodie, P.J.; Wiley, J.D.; Cassera, M.B.; Callmander, M.W.; Rakotondrajaona, R.; Rakotobe, E.; Rasamison, V.E.; Tendyke, K.; et al. Antiproliferative compounds from Cleistanthus boivinianus from the Madagascar dry forest. J. Nat. Prod. 2015, 78, 1543–1547. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Wu, X.-D.; Wang, S.-Y.; Wang, L.; He, J.; Li, G.-T.; Ding, L.-F.; Gong, X.; Dong, L.-B.; Song, L.-D.; Li, Y.; et al. Labdane diterpenoids and lignans from Calocedrus macrolepis. Fitoterapia 2013, 85, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Hung, H.C.; Sung, P.J.; Chen, I.S.; Kuo, W.L. Aporphine alkaloids and cytotoxic lignans from the roots of Illigera luzonensis. Phytochemistry 2011, 72, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Doussot, J.; Mathieu, V.; Colas, C.; Molinié, R.; Corbin, C.; Montguillon, J.; Moreno, Y.; Banuls, L.; Renouard, S.; Lamblin, F.; et al. Investigation of the Lignan Content in Extracts from Linum, Callitris and Juniperus Species in Relation toTheir In Vitro Antiproliferative Activities. Planta Med. 2017, 83, 574–581. [Google Scholar] [PubMed]
- Kuo, Y.-C.; Kuo, Y.-H.; Lin, Y.-L.; Tsai, W.-J. Yatein from Chamaecyparis obtusa suppresses herpes simplex virus type 1 replication in HeLa cells by interruption the immediate-early gene expression. Antivir. Res. 2006, 70, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Usia, T.; Watabe, T.; Kadota, S.; Tezuka, Y. Metabolite-cytochrome P450 complex formation by methylenedioxyphenyl lignans of Piper cubeba: Mechanism-based inhibition. Life Sci. 2005, 76, 2381–2391. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Chang, Y.L.; Teng, C.M.; Chen, I.S. Anti-platelet aggregation alkaloids and lignans from Hernandia nymphaeifolia. Planta Med. 2000, 66, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Picking, D.; Chambers, B.; Barker, J.; Shah, I.; Porter, R.; Naughton, D.P.; Delgoda, R. Inhibition of cytochrome P450 activities by extracts of Hyptis verticillata Jacq.: Assessment for potential HERB-drug interactions. Molecules 2018, 23, 430. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Sepúlveda, A.M.; Mendieta-Serrano, M.; Mojica, M.Y.A.; Salas-Vidal, E.; Marquina, S.; Villarreal, M.L.; Puebla, A.M.; Delgado, J.I.; Alvarez, L. Cytotoxic podophyllotoxin type-lignans from the steam bark of Bursera fagaroides var. fagaroides. Molecules 2012, 17, 9506. [Google Scholar] [CrossRef] [PubMed]
- Puebla-Pérez, A.M.; Huacuja-Ruiz, L.; Rodríguez-Orozco, G.; Villaseñor-García, M.M.; Miranda-Beltrán, M.D.L.L.; Celis, A.; Sandoval-Ramírez, L. Cytotoxic and antitumour activity from Bursera fagaroides ethanol extract in mice with L5178Y lymphoma. Phytother. Res. 1998, 12, 545–548. [Google Scholar] [CrossRef]
- Guerram, M.; Jiang, Z.-Z.; Zhang, L.-Y. Podophyllotoxin, a medicinal agent of plant origin: Past, present and future. Chin. J. Nat. Med. 2012, 10, 161–169. [Google Scholar] [CrossRef]
- Acton, Q.A. Advances in Podophyllotoxin Research and Application: 2012 Edition: ScholarlyBrief; Scholarly Editions: Atlanta, GA, USA, 2012. [Google Scholar]
- Kamal, A.; Ali Hussaini, S.M.; Malik, M.S. Recent Developments Towards Podophyllotoxin Congeners as Potential Apoptosis Inducers. Anti-Cancer Agents Med. Chem. 2015, 15, 565–574. [Google Scholar] [CrossRef]
- Lee, K.-H.; Xiao, Z. Podophyllotoxins and Analogs; CRC Press: Boca Raton, FL, USA, 2012; pp. 95–122. [Google Scholar]
- Ionkova, I. Podophyllotoxin and Related Lignans: Biotechnological Production by In Vitro Plant Cell Cultures; CAB International: Wallingford, Oxfordshire, UK, 2010; pp. 138–155. [Google Scholar]
- Khaled, M.; Jiang, Z.-Z.; Zhang, L.-Y. Deoxypodophyllotoxin: A promising therapeutic agent from herbal medicine. J. Ethnopharmacol. 2013, 149, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Guerram, M.; Jiang, Z.-Z.; Sun, L.; Zhu, X.; Zhang, L.-Y. Antineoplastic effects of deoxypodophyllotoxin, a potent cytotoxic agent of plant origin, on glioblastoma U-87 MG and SF126 cells. Pharmacol. Rep. 2015, 67, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Morales-Serna, J.A.; Cruz-Galicia, E.; Garcia-Rios, E.; Madrigal, D.; Gavino, R.; Cardenas, J.; Salmon, M. Three new diarylbutane lignans from the resin of Bursera fagaroides. Nat. Prod. Res. 2013, 27, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Mojica, M.A.; Leon, A.; Rojas-Sepulveda, A.M.; Marquina, S.; Mendieta-Serrano, M.A.; Salas-Vidal, E.; Villarreal, M.L.; Alvarez, L. Aryldihydronaphthalene-type lignans from Bursera fagaroides var. fagaroides and their antimitotic mechanism of action. RSC Adv. 2016, 6, 4950–4959. [Google Scholar] [CrossRef]
- Gu, J.-Q.; Park, E.J.; Totura, S.; Riswan, S.; Fong, H.H.S.; Pezzuto, J.M.; Kinghorn, A.D. Constituents of the twigs of Hernandia ovigera that inhibit the transformation of JB6 murine epidermal cells. J. Nat. Prod. 2002, 65, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Arreguin, P.; Arteaga-Nieto, P.; Reynoso-Orozco, R.; Villagomez-Castro, J.C.; Sabanero-Lopez, M.; Puebla-Perez, A.M.; Calvo-Mendez, C. Bursera fagaroides, effect of an ethanolic extract on ornithine decarboxylase (ODC) activity in vitro and on the growth of Entamoeba histolytica. Exp. Parasitol. 2008, 119, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Gutierrez, F.; Puebla-Perez, A.M.; Gonzalez-Pozos, S.; Hernandez-Hernandez, J.M.; Perez-Rangel, A.; Alvarez, L.P.; Tapia-Pastrana, G.; Castillo-Romero, A. Antigiardial Activity of Podophyllotoxin-Type Lignans from Bursera fagaroides var. fagaroides. Molecules 2017, 22, 799. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Bianchi, E.; Trumbull, E.R. Antitumor agents from Bursera microphylla (Burseraceae) II: Isolation of a new lignan—Burseran. J. Pharm. Sci. 1969, 58, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, K.; Ishiguro, T.; Mizuguchi, H.; Komeshima, N.; Koga, K.; Tsukagoshi, S.; Tsuruo, T.; Tashiro, T.; Tanida, S.; Kishi, T. Stereoselective reactions. XVII. Absolute structure-cytotoxic activity relationships of steganacin congeners and analogs. J. Med. Chem. 1991, 34, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, K.; Koga, K. Stereoselective total synthesis of optically active trans- and cis-burseran. Determination of the stereochemistry of natural antitumor burseran. Heterocycles 1979, 12, 1523–1528. [Google Scholar] [CrossRef]
- Gigliarelli, G.; Zadra, C.; Cossignani, L.; Robles Zepeda, R.E.; Rascon-Valenzuela, L.A.; Velazquez-Contreras, C.A.; Marcotullio, M.C. Two new lignans from the resin of Bursera microphylla A. gray and their cytotoxic activity. Nat. Prod. Res. 2017, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Tian, X.; Fan, P.C.; Zhan, Y.J.; Shen, D.W.; Jin, Y. Separation, determination and identification of the diastereoisomers of podophyllotoxin and its esters by high-performance liquid chromatography/tandem mass spectrometry. J. Chromatogr. A 2008, 1210, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Alina, C.-M.C.; Rocio, R.-L.; Aurelio, R.-M.M.; Margarita, C.-M.M.; Angelica, R.-G.; Ruben, J.-A. Chemical composition and in vivo anti-inflammatory activity of Bursera morelensis Ramirez essential oil. J. Essent. Oil-Bear. Plants 2014, 17, 758–768. [Google Scholar] [CrossRef]
- Rzedowski, J.; Medina Lemos, R.; Calderón de Rzedowski, G. Segunda restauración de Bursera ovalifolia y nombre nuevo para otro componente del complejo de B. simaruba (Burseraceae). Acta Bot. Mex. 2007, 81, 45–70. [Google Scholar] [CrossRef]
- Buckingham, J. Dictionary of Natural Products; Taylor & Francis: Abingdon, UK, 1993. [Google Scholar]
- Chang, C.-C.; Lien, Y.-C.; Liu, K.C.S.C.; Lee, S.-S. Lignans from Phyllanthus urinaria. Phytochemistry 2003, 63, 825–833. [Google Scholar] [CrossRef]
- Ito, C.; Itoigawa, M.; Ogata, M.; Mou, X.Y.; Tokuda, H.; Nishino, H.; Furukawa, H. Lignans as anti-tumor-promoter from the seeds of Hernandia ovigera. Planta Med. 2001, 67, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ohigashi, H.; Koshimizu, K. Anti-tumor promotion with food phytochemicals: A strategy for cancer chemoprevention. Biosci. Biotechnol. Biochem. 1996, 60, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Meng, Y.; Gearing, R.P.; Herald, D.L.; Pettit, R.K.; Doubek, D.L.; Chapuis, J.-C.; Tackett, L.P. Antineoplastic Agents. 522. Hernandia peltata (Malaysia) and Hernandia nymphaeifolia (Republic of Maldives). J. Nat. Prod. 2004, 67, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Noguera, B.; Diaz, E.; Garcia, M.V.; Feliciano, A.S.; Lopez-Perez, J.L.; Israel, A. Anti-inflammatory activity of leaf extract and fractions of Bursera simaruba (L.) Sarg (Burseraceae). J. Ethnopharmacol. 2004, 92, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Maldini, M.; Montoro, P.; Piacente, S.; Pizza, C. Phenolic compounds from Bursera simaruba Sarg. bark: Phytochemical investigation and quantitative analysis by tandem mass spectrometry. Phytochemistry 2009, 70, 641–649. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |
Figure 1.
Structures of dibenzyl butyrolactone lignans isolated from Bursera spp.

Figure 2.
Aryltetraline lignans (picro series) isolated from Bursera spp.

Figure 3.
Aryltetraline lignans isolated from Bursera spp.

Figure 4.
7′,8′-Dehydro-aryltetraline lignans isolated from Bursera spp.

Figure 5.
Dibenzylbutane diol lignans isolated from Bursera spp.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marcotullio, M.C.; Curini, M.; Becerra, J.X. An Ethnopharmacological, Phytochemical and Pharmacological Review on Lignans from Mexican Bursera spp. Molecules 2018, 23, 1976. https://doi.org/10.3390/molecules23081976
AMA Style
Marcotullio MC, Curini M, Becerra JX. An Ethnopharmacological, Phytochemical and Pharmacological Review on Lignans from Mexican Bursera spp. Molecules. 2018; 23(8):1976. https://doi.org/10.3390/molecules23081976
Chicago/Turabian StyleMarcotullio, Maria Carla, Massimo Curini, and Judith X. Becerra. 2018. "An Ethnopharmacological, Phytochemical and Pharmacological Review on Lignans from Mexican Bursera spp." Molecules 23, no. 8: 1976. https://doi.org/10.3390/molecules23081976