
Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides



1
Food Science and Technology Program, Beijing Normal University–Hong Kong Baptist University United International College, Zhuhai 519087, China
2
Department of Anaesthesiology, Laboratory and Clinical Research Institute for Pain, The University of Hong Kong, Hong Kong, China
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(14), 2556; https://doi.org/10.3390/molecules24142556
Submission received: 21 June 2019
/
Revised: 11 July 2019
/
Accepted: 12 July 2019
/
Published: 13 July 2019
(This article belongs to the Special Issue Discovery and Development of Antidiabetic Agents from Natural Products)
Abstract
:Diabetes mellitus is a multifactorial, heterogeneous metabolic disorder, causing various health complications and economic issues, which apparently impacts the human’s life. Currently, commercial diabetic drugs are clinically managed for diabetic treatment that has definite side effects. Dietary polysaccharides mainly derive from natural sources, including medicinal plants, grains, fruits, vegetables, edible mushroom, and medicinal foods, and possess anti-diabetic potential. Hence, this review summarizes the effects of dietary polysaccharides on diabetes and underlying molecular mechanisms related to inflammatory factors, oxidative stress, and diabetes in various animal models. The analysis of literature and appropriate data on anti-diabetic polysaccharide from electronic databases was conducted. In vivo and in vitro trials have revealed that treatment of these polysaccharides has hypoglycemic, hypolipidemic, antioxidant, and anti-inflammatory effects, which enhance pancreatic β-cell mass and alleviates β-cell dysfunction. It enhances insulin signaling pathways through insulin receptors and activates the PI3K/Akt pathway, and eventually modulates ERK/JNK/MAPK pathway. In conclusion, dietary polysaccharides can effectively ameliorate hyperglycemia, hyperlipidemia, low-grade inflammation, and oxidative stress in type 2 diabetes mellitus (T2DM), and, thus, consumption of polysaccharides can be a valuable choice for diabetic control.
1. Introduction
Diabetes mellitus is a multifactorial metabolic disorder described by chronic hyperglycemia due to insulin resistance or insulin insufficiency [1]. It is a heterogeneous disorder that can potentially cause insulin resistance in the peripheral tissues, including adipose, liver, and muscle, as well as progressive β-cell dysfunction in the islets of the pancreas, resulting in hyperglycemia [2]. In a patient with type 2 diabetes mellitus (T2DM), it is estimated that almost 90% of all the patients with diabetes show hyperglycemia, impaired glucose tolerance, dyslipidemia, hyperinsulinemia, and persistent insulin deficiency [3]. Moreover, the long-lasting complications of T2DM cause extraordinary healthcare costs with high morbidity and mortality [2,4]. Administration of oral hypoglycemic agents, such as biguanides, thiazolidinediones, and sulfonylureas, can meritoriously regulate hyperglycemia. Nevertheless, they have noticeable side-effects, including hypoglycemia and gastrointestinal troubles [5]. Thus, there is an urgent requirement for effective substitutions to reduce the complications of diabetes with lower side-effects. In recent years, the search for alternate medications has drawn great attention to combat diabetes. Therefore, dietary active principles progressively come into the field of investigator’s vision [4]. Dietary active principles, such as polysaccharides, alkaloids, steroids, polyphenol, terpenoids, glycoproteins, and saponins, have medicinal value and have been connected with various pharmacological effects, such as improving immune function, anti-diabetic, antioxidant, anti-cancer, anti-aging, antimicrobial, and anti-inflammatory properties [6,7,8,9].
Dietary polysaccharides are natural edible polysaccharides that are essential for our day to day life. Polysaccharides generally extracted from medicinal plants, grains, fruits, vegetables, edible mushroom, and medicinal foods have shown more consideration from investigators due to their low toxicity and numerous pharmacological activities [10,11,12]. They are composed of the monosaccharide unit and linked by glycosidic bonds. The assessment of the effects of polysaccharides with anti-diabetic properties has emerged as an important research field [10]. Various investigations have presented that polysaccharides purified from pumpkin, sea cucumber, goji berry, mushroom, bean, tea, and oat exert favorable effects on glucose homeostasis (Figure 1), reduce the complications of diabetes through the defensive mechanism against oxidative stress injury, and eventually improve insulin sensitivity [13,14,15]. In addition, dietary non-digestible polysaccharides obtained from plants and foodstuff are recognized potent modulators of gut microbiota that can nourish certain useful microbes in the human gut. This has also led to increased attention to the isolation of novel bioactive polysaccharides and their practices as functional components, which modulate the gut microbiota to improve the host’s metabolism and health [1]. Hence, polysaccharides are considered prebiotic substances, which are being speculated to confer positive effects in managing metabolic diseases like diabetes. In this comprehensive review, we summarize some of the most common dietary polysaccharides from medicinal plants, grains, fruits, vegetables, edible mushroom, and medicinal foods that impact metabolic health and discuss underlying molecular mechanisms, related to oxidative stress and inflammatory factors, which could be supportive in ameliorating type-2 diabetes.
2. Anti-Diabetic Potentials of Polysaccharides
Dietary polysaccharides are largely obtained from natural sources, including medicinal plants, grains, fruits, vegetables, mushroom, medicinal foods, algae, and fungi. They have potential anti-diabetic effects with underlying various molecular mechanisms to combat diabetic complications. Polysaccharides proceed to regulate hyperglycemia primarily established on their sources, composition, and preparation. Polysaccharides have been documented to have potent anti-diabetic activity. β-d-(1→6)-glucan can improve the insulin level and hepatic glycogen accumulation, decreasing the blood glucose level in streptozotocin (STZ)-induced diabetic mice [14]. A polysaccharide purified from Lycium barbarum has a composition of mainly mannose, rhamnose, and glucose. It can inhibit the absorption of glucose in a dose-dependent manner [16]. β-d-glucan of Agaricus blazei (MW: 136.05 kDa), Trametes gibbosa (MW: 3.872, 2.761, 8.526, and 5.659 kDa), Saccharina japonica (MW: 7.28 kDa), and Lachnum calyculiforme (MW: 445.363 kDa) has been documented to have hypoglycemic, hypotriglyceridemic, and hypocholesterolemic activities in diabetic rats. Moreover, the oligosaccharides hydrolyzed from β-d-glucan show diabetic improved activities [17,18,19,20]. A crude extract of Talinum triangulare polysaccharide contains rhamnose, arabinose, mannose, and galactose, which demonstrated an anti-diabetic effect in STZ diabetic rats [21]. Similarly, aqueous-soluble polysaccharides (MW: 60, 350, and 3000 kDa) obtained from Schisandra chinensis (Turcz.) Baill (ESCPs) is composed of L-rhamnose, L-arabinose, D-xylose, D-glucose, D-galactose, and D-mannose that evidently decreased the blood glucose level in alloxan-induced diabetic mice after 21-day oral administration [13]. A sulfated polysaccharide purified from Saccharina japonica primarily comprises fucose, sulfate uronic acid, galactose, mannose, glucose, and arabinose, which exhibited a potential hypoglycemic effect by markedly decreasing blood glucose and augmenting insulin levels in alloxan-induced diabetic mice [19]. Two polysaccharides (MW: 0.7, 3.5 kDa) obtained from Inula britannica (MW: 0.7, 3.5 kDa) contains mannose, glucuronic acid, rhamnose, galacturonic acid, glucose, galactose, and arabinose that markedly decreased the plasma glucose level and increased the liver glycogen content in alloxan-induced diabetic mice [22]. The anti-diabetic effects of various grains, fruits, vegetables, edible mushrooms, and medicinal foods have been listed in Table 1.
3. Mechanism of Dietary Polysaccharides on Anti-Diabetic Activities
Polysaccharides are generally extracted from dietary materials by various physical, chemical, or enzymatic digestion treatments that can be found to have anti-diabetic potentials. The previous study showed that polysaccharides consumption could alleviate diabetes through mechanisms of action on gastrointestinal viscosity, gastrointestinal satiety, colon fermentation, and anti-gastrointestinal inflammation [68]. Similarly, the present study aimed to identify the various in vivo and in vitro trials in which dietary polysaccharides have hypoglycemic, antioxidant, and anti-inflammatory effects. Dietary polysaccharides enhance pancreatic β-cell mass, trigger insulin signaling pathways through insulin receptors, and activate the PI3K/Akt pathway. They modulate ERK/JNK/MAPK pathways and, thus, alleviate β-cell dysfunction.
3.1. Hypoglycemic and Hypolipidemic Effects
Impaired glucose tolerance generally leads to permanent loss of β-cell function, which has been recognized by the occurrence of glucose toxicity and lipotoxicity. Hyperglycemia often produces an elevated reactive oxygen species in β-cells, providing succeeding impairment to cellular mechanisms [69,70]. The pancreatic lipid contents generally not relate with β-cell dysfunction in young-onset T2DM [71] and, however, pancreatic islet lipotoxicity is known as a major factor for the onset and progression of T2DM. The disorder of lipid metabolism or increasing fatty acid levels in the blood cause β-cell dysfunction, which are primary threat factors for T2DM [39]. An investigation of animal study connected with streptozotocin-induced diabetes succeeding β-cell dysfunction suggested that the polysaccharide obtained from gum exudates (Acacia tortilis), comprising of the polymer compounds (D-glucose, D-galactose, L-rhamnose, and D-glucuronic acid), showed a potential anti-diabetic effect. These extracts remarkably decreased fasting blood glucose (FBG), glycated hemoglobin (HbA1c), total cholesterol (TC), triglyceride (TG), low-density lipoprotein (LDL), very LDL (VLDL), and an elevated concentration of high-density lipoprotein (HDL). Furthermore, reduced hepatic markers, such as aspartate transaminase (AST) and alanine transaminase (ALT), were noticed in gum extract treated groups, indicating improved lipid metabolism in the liver [72]. Various animal studies reported that well characterized, low to high molecular weight polysaccharides obtained from various edible sources showed greater anti-diabetic effects [13,18,73]. A polysaccharide purified and characterized by Lachnum calyculiforme demonstrated a significant hypoglycemic effect (p < 0.01) at dose-dependent manners in alloxan-induced diabetic mice [18]. Similarly, corn silk, pumpkin, Mactra veneriformis, Trametes gibbosa, Inula britannica, Saccharina japonica, Phellinus linteus, Talinum triangulare, and Schisandra chinensis (Turcz.) Baill containing different ranges of molecular weight polysaccharides (60, 180, 220, 350, 449.6, 3000, 3172.9 kDa) exhibited a significant anti-diabetic effect (p < 0.05) as proved by a remarkable decrease of blood glucose and improvement of OGTT, serum insulin, and lipid metabolism outcomes in STZ as well as alloxan-induced diabetic mice [13,22,73,74,75].
The fruit polysaccharide of Morus alba L. has significant anti-hyperglycemic and anti-hyperlipidemic effects that can undoubtedly relieve symptoms of diabetes in the STZ-induced T2DM rat model. After seven weeks’ treatment, the fruit polysaccharides significantly diminished FBG, OGTT, glycated serum protein, and lipid profiles and improved insulin levels in the blood. Furthermore, the polysaccharides-treated groups enhanced the insulin-signaling pathway, and their high protein expression levels of InsR (insulin receptor), IRS-2 (insulin receptor substrate 2,), Akt (serine/threonine-specific protein kinase), and GLUT4 (glucose transporter 4) were identified when compared to that of the T2DM groups [76]. High-fat diet treated with STZ-induced hyperglycemia was significantly reduced by the administration of mushroom polysaccharides [77,78]. The oral administration of mushroom extracellular polysaccharides obtained from Pleurotus tuber-regium (20 mg/kg b.w. (body weight)) and Grifola frondosa (100 or 300 mg/kg b.w.) could decline the levels of FBG, TC, TG, lipid profiles, fatty acid composition, and expression of liver peroxisome proliferator-activated receptor alpha (PPAR-α) in obese-diabetic rats. The parallel restoration and elevated HDL-C levels occurred with supplementation of mushroom polysaccharides [77,78]. These hypolipidemic properties might be connected with up-regulated expression of liver PPAR-α mRNA and protein levels [78]. All these outcomes strongly suggest that polysaccharides exert potential hypoglycemic and hypolipidemic effects in STZ-induced diabetic animal models. Therefore, polysaccharides could be considered as a nutritional supplement to treat diabetic complications.
3.2. Increasing β-Cell Mass and Reducing β-Cell Dysfunction
A recent study described that lean and obese human, with T2DM, had a 45% and 70% decreased relative β-cell mass; 10- and 3-fold elevated β-cell apoptosis, respectively, compared with the respective nondiabetic control group [79,80,81]. This research outcome suggested that the decreased β-cell mass along with elevated β-cell apoptosis rate is relatively common in T2DM. Though the underlying mechanism of β-cell apoptosis in T2DM is complex and debated [82,83], the prevention of β-cell apoptosis and connected elements are the vital approach for treating T2DM.
Several animal studies reported that purified, characterized low to high molecular weight polysaccharides obtained from dietary sources showed elevating β-cell mass and reducing β-cell dysfunction [84,85]. The oral administration of mulberry leaf containing polysaccharides (MW: 8.1 kDa) significantly prevented β-cell apoptosis and elevated insulin secretion in STZ-induced diabetic rats. These polysaccharides significantly up-regulated Bcl-2 (B-cell lymphoma 2) and PDX-1 (insulin promoter factor 1) and down-regulated mRNA expression of Bax (BCL2 associated X protein). In addition, they markedly prevented caspase-3 activation in the islets of the pancreas of STZ- diabetic rats [84]. These results suggested that polysaccharides could play a critical function in pancreatic islet cell protection from apoptosis by increasing the ratio of Bcl-2/Bax and improving insulin secretion through the restoration of PDX-1 in diabetic animals [84]. Similarly, a polysaccharide from Ganoderma atrum (MW: 1013 kDa) administration in diabetic animals significantly reduced FBG, plasma insulin, and expression of Bax and improved expression of Bcl2 as well as lipid profiles in the high-fat diet STZ-induced diabetic rats [85]. Histopathological studies also confirmed that polysaccharides from G. atrum showed elevated β-cell mass, pancreatic islets expansion, and restoration, representing that polysaccharides protected the islets of the pancreas from HFD- and STZ-induced damage [86]. Another study from fruit bodies of Ganoderma lucidum containing protein-bound polysaccharide (MW: 8.849 kDa) exhibited potential anti-hyperglycemic and anti-hyperlipidemic effects on STZ-induced diabetic rats [87]. The underlying mechanism of this study observed that G. lucidum significantly up-regulated Bcl-2 and down-regulated Bax and caspase in the pancreatic cells compared to that of STZ diabetic animals. The results strongly suggested that polysaccharide from G. lucidum exerted an anti-diabetic potential by inhibiting the β-cell apoptosis in diabetic rats [88].
Zhu et al. [89] demonstrated a low molecular weight and well-characterized polysaccharide from pumpkin fruit (MW: 115 kDa) that prevented β-cell apoptosis by regulating the mRNA expression of Bcl-2 and Bax in STZ-induced damage of pancreatic islet cells. They found that polysaccharides from pumpkin possessed strong antioxidant capacities and eventually decreased the NO level and restore the β-cells. Zhang et al. [34] also presented that water-soluble polysaccharide purified from pumpkin restored the damaged pancreatic islets via triggering β-cell multiplication. This investigation further observed that intragastric treatment of polysaccharide from pumpkin significantly decreased blood glucose, TC, TG, and HbA1c in alloxan-induced diabetic animals and restored the normalization within 21 days’ treatment of polysaccharides. A low molecular weight polysaccharide purified from Sea cucumber (Cucumaria frondosa, MW: 14.76 kDa) and Lycium barbarum L (goji berry, MW: 212.14 kDa) prevented β-cell apoptosis and increased β-cell mass in pancreatic islets of mice by down-regulating the mitochondrial signaling pathway, eventually showing significant insulin-sensitizing and anti-hyperglycemic effects [86,90]. All these findings recommend that polysaccharides aid in increasing β-cell mass and reducing β-cell dysfunction. Hence, polysaccharides could be considered as a dietary supplement involved in the pathogenesis of diabetes, leading to reduce the degree of β-cell damage in pancreatic islets.
3.3. Antioxidant Effects
Diabetes is generally caused by the impairment or insufficient β-cells in the pancreas that diminishes insulin biosynthesis and gradually deteriorates whole body functions. In contrast to physiological glucose concentration, these glucose levels negatively affect a greater number of organs and tissues. Due to chronic hyperglycemia, decreasing insulin secretion, as well as up-surging insulin resistance, provides glucose toxicity [81,91,92]. It has been recognized that glucose toxicity mainly participates in the deterioration of diabetes by distressing the synthesis of β-cells. The underlying mechanism of glucotoxicity is chiefly mediated by oxidative stress, which has been associated as a primary risk factor in the onset and advancement of T2DM. Oxidative stress is usually formed by an excess free radical formation and decreases the antioxidant defense system in the body [81]. Generally, the living system generates two major forms of reactive species, viz. reactive oxygen species and reactive nitrogen species. Pancreatic β-cells are susceptible to those reactive species due to their low concentration of free-radical scavenging enzymes. They can readily injure to cellular macromolecules, such as lipids, DNA, and proteins [93]. However, the antioxidant agents derived from dietary sources are promising elements to scavenge cell generating free radicals and protects β-cells. These antioxidant agents generally augment cellular antioxidant enzymes and inhibit β-cell apoptosis, which has been demonstrated to improve β-cell dysfunction and protect β-cells against glucotoxicity in diabetic animal models [5,94,95].
In vivo, animal studies demonstrated that bioactive polysaccharide could inhibit the development of T2DM by decreasing oxidative stress. For instance, the polysaccharide obtained from Grifola frondosa (MW: 400–450 kDa) and Salvia miltiorrhiza Bunge (MW: 119.5 kDa) showed substantial defensive and antioxidative ability against the oxidative damage and increased the activities of antioxidant enzymes, such as SOD (superoxide dismutase), CAT (catalase), and GSH-Px (glutathione peroxidase) and decreased level of malondialdehyde (MDA), NO synthase, and inducible NOS (Nitric oxide synthase) in blood and liver [96,97]. MDA is measured as a fundamental chain reaction of lipid peroxidation, which produces injury to the cell membrane, necrosis, and inflammation [92]. Additionally, G. frondosa and S. miltiorrhiza improved the insulin sensitivity index and attenuated STZ-induced structural changes to the pancreas and liver [96,97]. Similarly, the low molecular weight polysaccharides from Catathelasma ventricosum (MW: 160 kDa) and Ophiopogon japonica demonstrated anti-diabetic, anti-obesity, and antioxidant activities in STZ-induced diabetic mice. Oral administration of both plants decreased the MDA levels and increased vitamin E contents, SOD, CAT, and GSH-Px activities in the hepatic and renal cells of STZ-induced diabetic mice [14,98]. Simultaneously, oral administration of these polysaccharides significantly decreased blood glucose and markedly elevated serum insulin levels. Microscopic observation in the pancreas, kidneys, and liver assay confirmed that polysaccharides protected the organs from lipid peroxidation injury and conserved tissue integrity [99]. The enhancement of antioxidant enzyme activity in the treated group designated that polysaccharides inhibited the cell injury by scavenging free radicals produced by chain reactions of lipid peroxidation [70].
Mulberry fruit polysaccharide, produced by Fructus Mori, is a biopolymer that exhibited hypoglycemic and antioxidant activities in vitro as well as in STZ-induced diabetic mice. In vitro, hypoglycemic experiments exhibited that a noteworthy insulin-sensitizing and increased insulin synthesis was observed when the treatment with polysaccharides stimulated pancreatic β cell proliferation and serum insulin levels. Oral administration of fruit polysaccharide could markedly decrease blood glucose and MDA levels and increase SOD, CAT, and GSH-Px in the hepatocytes of STZ-induced diabetic mice. Histopathological observation exhibited that fruit polysaccharide could significantly improve the tissue damage to the pancreas, liver, and kidney [100]. Likewise, ginseng polysaccharides from Panax ginseng C.A. Meyer showed significant hypoglycemic and antioxidant activities in STZ-induced diabetic mice. Oral administration of ginseng polysaccharides significantly decreased blood glucose and lipid peroxidation levels and enhanced SOD levels [101].
Another study in white oyster culinary-medicinal mushroom polysaccharide, which was obtained from Pleurotus florida (MW: 155 kDa), showed decreased blood glucose, HbA1c, lipid profiles, and urinary glucose in STZ- induced diabetic rats. P. florida decreased the levels of MDA and nitric oxide and restored the levels of GSH, SOD, and CAT in diabetic rats. These findings recommended that administration of P. florida could attenuate diabetic complications along with hyperglycemia and hypercholesteremic effects [102]. Based on the investigations, all these results strongly suggest that polysaccharides possess antioxidant properties that can be applied as an adjunct therapy and control the effect of T2DM.
3.4. Anti-Cholesterolemic and Anti-Triglyceridemic Effects
Various in vitro and in vivo studies demonstrated that dietary polysaccharides potentially have lipid-lowering effects and eventually reduce the effects of diabetic complications. Polysaccharides activate serine/threonine protein kinase (AMPK) pathway to regulate lipid metabolism by decreasing the levels of triglycerides and cholesterol. Studies have revealed that AMPK switches off anabolic processes, including the biosynthesis of fatty acids, triglyceride, and cholesterol, through repressing the expression of genes, such as Acetyl-CoA carboxylase (ACC), sterol regulatory element binding protein -1c (SREBP-1c), and 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) reductase [103,104,105].
Normally, the reduction of triglycerides occurs in the human body through activation of an enzyme, adipose triglyceride lipase, or up-regulation of peroxisome proliferator-activated receptor-α (PPAR-α) and PPAR gamma coactivator-1 alpha pathways [106,107]. These signaling pathways are highly connected with energy expenditure as well as reduce the uptake of energy substrates. Furthermore, triglyceride levels can be decreased by triggering another enzyme, ACC, and up-regulation of SREBP-1c or down-regulation of FAS (fatty acid synthase)-carnitine palmitoyltransferase-1 (CPT1) signaling pathways. Studies have shown that over-expression of CPT1 can elevate fatty acid oxidation, lessen cellular triglyceride accumulation, and reduce high-fat-diet-induced insulin resistance [107,108]. The activity of CPT1 is generally regulated by ACC, through the manufacturing of malonyl-CoA, which acts as an inhibitor of CPT1 [108].
The consumption of dietary polysaccharides can be another effective treatment or can prevent hypercholesterolemia. The dietary polysaccharides exert cholesterol lowering effects via activation of sterol regulatory element binding protein -2 (SREBP-2) or inhibition of rate-limiting enzyme, 3-hydroxy-3-methylglutaryl-CoA reductase (HMG-CoA reductase). The expression of HMG-CoA reductase gene is controlled by the SREBP, which play vital roles in managing the biosynthesis of cholesterol and fatty acids. Dietary polysaccharides have inhibitory effects on HMG-CoA reductase through the activation of SREBP-2 [107,109,110,111]. Increasing plasma LDL-cholesterol (LDL-C) level is a major cardiovascular risk for diabetic patients [112,113]. Uptake of these LDL-C through LDL receptor-mediated endocytosis is an essential step for the regulation of cholesterol homeostasis [114]. Hence, the activation of the LDL receptor reduces the plasma LDL-C levels. Proprotein convertase subtilisin/kexin type 9 is a class of proteinase, that can usually degrade LDL receptor resulting in the elevation of LDL-C in the blood [115]. Dietary polysaccharides are known to activate LDL receptors via inhibiting this enzyme and prevent the elevated LDL-C levels in diabetic patients [111]. Anti-cholesterolemic and anti-triglyceridemic effects of dietary polysaccharides are listed in Table 2. Based on the studies, all these outcomes strongly suggest that polysaccharides possess anti-cholesterolemic and anti-triglyceridemic effects that can be applied as an adjunct therapy for CVD (Cardiovascular diseases) and control the effect of T2DM.
3.5. Anti-Inflammatory Effects
In general, oxidative stress is connected with chronic inflammation in T2DM. During inflammation, several pro-inflammatory cytokines, namely, interleukin (IL)-1, IL-6, IL-8, IL-12, IL-18, interferon gamma (IF-γ), and tumor necrosis factor alpha (TNF-α), serve key functions in the dysfunction of islet cells and insulin receptors in the pancreas and eventually β-cell death [3,4,95,121]. Studies have shown that treatment using anti-inflammatory agents, including IL-1 and receptor antagonists, normalize glucose in the blood, improve insulin secretion, and decrease inflammation causing islet fibrosis. These findings suggested the improvement of β-cell dysfunction and cell survival [122].
Various preclinical studies validated that bioactive polysaccharide can inhibit the progression of T2DM by decreasing pro-inflammatory factors. A high molecular weight polysaccharide (MW: 72.9 kDa), which was obtained from the roots of Angelica sinensis (Oliv.) Diels, exhibited hypoglycemic and hypolipidemic effects in STZ-induced diabetic mice. These root polysaccharides markedly exhibited anti-inflammatory effects by decreasing the insulin receptor-associated inflammatory factors, including IL-6 and TNF-α, in STZ-induced diabetic mice [123]. Generally, IL-6 is synthesized by macrophages and T cells for triggering immune responses to the host. IL-6 can also certainly avert insulin synthesis, and excess IL-6 leads to severe pancreatic islet cytotoxicity and cause insulin resistance [4]. TNF-α is a well-recognized cell signaling protein, which is certainly correlated with the insulin receptor and β-cell dysfunction and vastly articulated in adipose cells [124,125]. Animal studies connected with T2DM exhibited that the polysaccharide from Rehmannia glutinosa (Gaertn.) DC (MW: 63.5 kDa) meritoriously improved hyperglycemia, vascular inflammation, hyperlipidemia, and oxidative stress [124].
A low molecular weight polysaccharide (MW: 50–210 kDa) acquired from Pseudostellaria heterophylla demonstrated a hypoglycemic potential in STZ-induced type 2 diabetic rats. These polysaccharides improved insulin sensitivity and markedly decreased lipid profiles and TNF-α expression and elevated IL-10 concentration. IL-10 has generally pleiotropic properties on inflammation, in which the polysaccharides avert the inflammatory mechanism by condensing the secretion and activities of proinflammatory cytokines. These findings clearly exhibited that polysaccharides attenuated low-grade inflammation connected with T2DM [126]. Similarly, the fruit body of mushroom polysaccharide from Pleurotus sajor-caju and Ramulus mori and the polysaccharide extracted from Morus alba L. reduced blood glucose and attenuated hyperglycemia and hyperinsulinemia in diabetic mice. In addition, the polysaccharides decreased the expression of various proinflammatory cytokines, including IL-6, IL-8, COX-2 (cyclooxygenase-2), TNF-α, by down-regulating the signaling of nuclear factor kappa B (NF-kB) [127,128]. Preventing these inflammatory factors are normally positive strategies for averting or alleviating pancreatic islet damage and reducing T2DM development. Hence, polysaccharides greatly normalize the pancreatic function from STZ-induced damage, and this normalization could be connected with a reduction of inflammatory factors and oxidative stress in pancreatic islets [129].
Various low molecular weight polysaccharides extracted and purified from Misgurnus anguillicaudatus (MW: 130 kDa), Anoectochilus roxburghii, Vigna radiata L., and Hedysarum polybotrys demonstrated anti-hyperglycemic, antioxidant, anti-inflammatory, and anti-hyperlipidemic effects in diabetic animals, which was reflected by decreased blood glucose, MDA, MCP-1 (Monocyte chemoattractant protein 1), TNF-α, IL-6, lipid profiles, and boosted the synthesis of insulin and elevated activities of SOD and GSH-Px in STZ induced diabetic animals [124,125,130,131]. Taken together, polysaccharides obtained from various dietary sources can effectively ameliorate hyperglycemia, hyperlipidemia, low-grade inflammation, and oxidative stress in T2DM, and, therefore, intake of polysaccharides can be a potential beneficial choice for diabetes.
3.6. Inhibition of α-Amylase and α-Glucosidase
Generally, an experimental indicator of T2DM is hyperglycemia; it is well-defined as abnormally elevated fasting and postprandial glucose levels in the blood. Hence, managing postprandial hyperglycemia is a main beneficial strategy for the management of diabetes. Dietary carbohydrates are naturally digested into monosaccharides, such as glucose and fructose; these monosaccharides can be readily uptaken by the small intestine and transfer into the blood circulation. The human body normally has several dynamic carbohydrates–digestive enzymes, of which α-amylase (saliva or pancreas) and α-glucosidase (small-intestine) are most distinct. α-amylases are present in saliva and pancreas that degrade polysaccharide into glucose. Likewise, α-glucosidases are vital for assimilating oligosaccharides to monosaccharides in the small intestine. Hence, restraining of these digestive enzymes notably inhibit the conversion of polysaccharides into blood glucose, which serves as an effective step to control the blood glucose in diabetic patients [132]. In addition, the hypoglycemic effect of polysaccharides has been achieved by changing the small intestine transit time and preventing the carbohydrate digestion by suppression of digestive enzyme. These inhibitions are generally accomplished by dietary components, such as inulin, tannin, and phytic acid [133]. All these steps can be an effective strategy in diabetes for controlling the blood glucose level.
Various in vitro studies showed that low-molecular-weight bioactive polysaccharides obtained from the fruits of blackcurrant (Ribes nigrum L.), an alkaline soluble polysaccharide from Coreopsis tinctoria; fucoidan polysaccharide from Turbinaria conoides; polysaccharide fraction from Diaphragma juglandis fructus exhibited higher antioxidant, α-amylase, and α-glucosidase inhibitory activities that showed higher bioactive, with hypoglycemic, potential [134,135,136,137]. Furthermore, these polysaccharides significantly prevented the synthesis of NO, TNF-α, and IL-6 in LPS (Lipopolysaccharide)-stimulated BV2 (raf/myc-immortalised murine neonatal microglial cell line) microglial cells [134]. Hence, all these investigations recommended that bioactive polysaccharides could serve as potential hypoglycemic agents to be applied as functional foods or alternative supplements. Various molecular weight polysaccharides isolated from the pulp of apricot (Armeniaca sibirica L. Lam., MW: 25.93 kDa) [138]; seeds of Plantago asiatica L. (MW: 1894 kDa) [139]; Fucoidan from sea cucumber [67], Turbinaria ornate [140], Fucus vesiculosus [17], and Sargassum wightii [141] demonstrated significant inhibition of α-glucosidase and α-amylase activities in vitro. All these crude polysaccharide extracts exhibited a significant α-amylase and α-glucosidase inhibitory effect in a dose-dependent manner. Based on the observation in in vitro studies, all these results strongly suggest that polysaccharides remarkably inhibit carbohydrate digesting enzymes, α-glucosidase, and α-amylase activities, which regulate blood glucose levels. Hence, polysaccharides serve as an effective component to control the hyperglycemic conditions in diabetic patients.
3.7. Increasing Insulin Signaling Pathways
Elevated blood glucose normally elicits the synthesis of insulin in the pancreatic β-cells. Secretion of insulin instantly binds to its membrane receptor, which stimulates a cascade sequence of mechanism. This series of mechanism subsequently aids to increase glucose influx and metabolic effects, including glycolysis, glycogenesis, and avert glycogenolysis. Furthermore, insulin triggers regular cellular and physiological functions, comprising the cell division, apoptosis, and autophagy [3,142]. This cascade mechanism starts with the autophosphorylation of tyrosine residues in the intracellular components of the insulin receptor, which phosphorylate various substrates, including IRS1 and IRS2. Both substrates fix and activate the PI3K (PI3K: phosphoinositide 3-kinase)/Akt pathway as well as the MAPK (MAPK: mitogen-activated protein kinase) pathway. Akt is a main mediator to activate the most biochemical mechanism in glucose metabolism via activating phosphofructokinase and deactivating glycogen synthase kinase, resulting in stimulation of glucose transporter system translocation [3]. MAPK is a specific protein kinase involved in various physiological and biochemical mechanisms, including cell differentiation, proliferation, apoptosis, and cell endurance. ERK1/2 (extracellular-signal-regulated kinase 1/2) and JNK (c-Jun N-terminal kinase) are other cell signaling kinases co-task with MAPK, involved in cell growth, differentiation, inflammatory response, and apoptosis [4] (Figure 2). Overstimulation of MAPK generally provides the failure of insulin synthesis linked with apoptosis process in pancreatic islet cells [3].
3.7.1. Activation of the PI3K/Akt Pathway
The different molecular weight of polysaccharides acquired from Ophiopogon japonicas (MW: 3.47, 6.746, 35.2, 124.3, and 324.6 kDa), Acaudina molpadioides (MW: 1614.1 kDa), and mulberry leaf (MW: 289) reduced hyperglycemia and hyperinsulinemia in STZ-induced diabetic mice. Polysaccharide triggers the PI3K/Akt signaling pathway through IRS1, PI3K-p85, and phosphorylated Akt develops insulin sensitivity [26,123,143] and improves diabetic-associated renal disease [144]. Moreover, treatment of these polysaccharides increased GLUT4 levels in pancreas and decreased glycogen synthase kinase-3β levels in most of the cells. This observation showed that polysaccharides demonstrated anti-diabetic agents by triggering the signaling pathways of PI3K/Akt/GSK-3/GLUT-4 [144].
Polysaccharide derived from Ganoderma atrum, Enteromorpha prolifera, and Liriope spicata var. prolifera (MW: 3.2 and 4.29 kDa) markedly decreased FBG and significantly improved plasma lipid profiles and glucose tolerance in diabetes-induced endothelial dysfunction in animal models. In addition, administration of polysaccharides remarkably inhibited the expression of GSK-3β (glycogen synthase kinase-3β) and elevated expressions of the insulin receptor, IRS1, PI3K, AKT, eNOS, and GLUT4 in the liver of diabetic rats [85,99,145,146,147]. An in vitro study also showed that the polysaccharide from Grifola frondosa significantly increased glucose metabolism and glycogen synthesis in HepG2 cells. Western blot findings demonstrated that polysaccharide triggered insulin receptor and elevated Akt expression, thereby inhibiting GSK-3β expression [148]. Another in vitro study demonstrated that the fruit of high molecular weight polysaccharide Lycium barbarum L. (MW: 33.867 kDa) elevated expressions of PI3K, p38 MAPK, and glucose uptake by GLUT4 in isolated adipocytes and reduced insulin receptors in obese and diabetic rats [74,149].
Sea cucumber containing polysaccharides consisted of a chondroitin sulfate E backbone, which decreased the level of glucose in the blood by stimulating PI3K/GLUT4 and elevated the phosphorylation of insulin receptors, IRS1, and p85-PI3K in the skeletal muscles of T2DM [90,150,151]. Western blot findings demonstrated that polysaccharides enhanced the protein expressions of IRS2, PI3K, and glycogen synthase and lowered that of GSK-3β in the liver of type 2 diabetic mice. At the end of the experiment, these findings suggested that sea cucumber polysaccharides increased glucose metabolism through the PI3K/GLUT4/GSK-3β signaling pathway [90,150,151]. A low molecular weight polysaccharide (120 kDa) extracted from tea (Camellia sinensis L.) demonstrated hypoglycemic, hypolipidemic, and insulin-sensitizing effects in obese and diabetic mice. In addition, these polysaccharides improved SOD, CAT, and GSH-Px activities in liver and kidney tissue of diabetic mice. Tea polysaccharides increased the expressions of PI3K/AKT p-AKT and GLUT4 signaling pathway [27,88,152]. All these above findings proved that administration of polysaccharides remarkably inhibited the expression of GSK-3β and elevated expressions of the insulin receptor, IRS1, PI3K, and AKT in type 2 diabetic animal models.
3.7.2. Modulation of the MAPK Pathway
Sea cucumber polysaccharide (MW: 21.53 kDa) purified from Acaudina Molpadioides (MW: 20.53 kDa,) and pumpkin polysaccharides (MW: 749.3, 727.0, and 607.6 kDa) and sulfated rhamnose polysaccharides (MW: 4.57 kDa) from Enteromorpha prolifera increased insulin-stimulated glucose uptake, GLUT4 translocation, and Akt/ERK activation in TNF-α-induced insulin-resistant 3T3-L1 adipocytes. This finding strongly suggested that polysaccharide enhanced glucose uptake by activating the PI3K/Akt pathway and MAPK–ERK pathway [90,150,151,152]. Another in vitro study connected with active polysaccharides on LPS-induced RAW 264.7 cells; polysaccharides derived from Agaricus blazei Murill decreased the expression of JNK, ERK, and p38 [132,153,154]. Xu et al. [155] purified polysaccharide (MW: 460 kDa) from Ramulus mori (M. alba L.) that decreased FBG and HbA1c levels and augmented insulin levels in STZ-induced type 2 diabetic mice. Western blot studies in pancreatic tissue exhibited that polysaccharide down-regulated the expression of p-JNK, p-p38, Bax, and cleaved-caspase-3 and increased Bcl-2 expression. This study strongly suggested that polysaccharide had a hypoglycemic effect by down-regulating the JNK/p38 pathway to inhibit pancreatic cell apoptosis [155].
4. Conclusions
Diabetes is currently a serious health issue worldwide producing significant morbidity and mortality, and there is no route to cure diabetes completely. The commercial oral hypoglycemic and anti-hyperglycemic drugs have their self-limitations, adverse effects, high cost, and secondary failure. In addition, these oral diabetic drugs cause serious complications, such as hypoglycemia, weight gain, abdominal pain, nausea, vomiting, edema, diarrhea, gas trouble, bloating, and an increase levels of LDL-C. Hence, screening active anti-diabetic agents from natural sources, including polysaccharide, is of greater attraction due to its lesser side effects. Various in vivo, in vitro, and clinical experiments in this review clearly showed that oral administration of polysaccharides reduced hyperglycemia and hyperlipidemia through underlying various molecular mechanisms. Anti-diabetic effects are mediated primarily by their antioxidant properties, as well as the succeeding methods include inhibition of α-amylase and α-glucosidase activity, improving glucose metabolism, increasing β-cell mass, and reducing β-cell dysfunction. Insulin signaling pathways are also increased through activating PI3K/Akt pathway and modulating MAPK/JNK/ERK pathway. Hypercholesterolemic and hyperlipidemic activities are also associated with diabetes in which dietary polysaccharides have a vital function in the activation of AMPK pathway and down-regulation of ACC, SREBP-1c, and HMG-CoA reductase that leads to reduce the levels of triglycerides and cholesterol. Hence, dietary polysaccharides could be considered as anti-diabetic agents and involved in alleviating the pathogenesis, leading to reduce the degree of β-cell damage in the pancreas.
Author Contributions
K.G. and B.X. conceived and designed the review; K.G. wrote the paper; B.X. critically revised and improved the manuscript.
Funding
This research was funded by one research grant from Zhuhai Higher Education Construction Project (Zhuhai Key Laboratory of Agricultural Product Quality and Food Safety).
Conflicts of Interest
The authors declared no conflicts of interest.
Abbreviations
| ACC | acetyl-CoA carboxylase |
| ACE | angiotensin converting enzyme |
| AGEs | advanced glycation end products |
| Akt | serine/threonine-specific protein kinase |
| ALP | alkaline phosphatase |
| ALT | alanine transaminase |
| AMPK | serine/threonine protein kinase |
| AST | aspartate transaminase |
| Bax | BCL2 associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| BUN | blood urea nitrogen |
| CAT | catalase |
| CPT1 | carnitine palmitoyltransferase-1 |
| CRE | creatinine |
| CVD | cardiovascular diseases |
| CRP | c-reactive protein |
| eNOS | endothelial nitric oxide synthase |
| ERK | extracellular-signal-regulated kinase |
| FBG | fasting blood glucose |
| FFA | free fatty acids |
| GLP-1 | glucagon-like peptide-1 |
| GLUT4 | glucose transporter 4 |
| GSH-Px | glutathione peroxidase |
| GSH-R | glutathione reductase |
| GSK- 3 | glycogen synthase kinase-3 |
| HbA1c | glycated hemoglobin |
| HDL-C | high-density lipoprotein –C |
| HepG2 | human liver cancer cell line |
| HFD | high-fat diet |
| IF-γ | interferon γ |
| IL | interleukin |
| IL-6 | interleukin-6 |
| InsR | insulin receptor |
| IPITT | Intraperitoneal Insulin Tolerance Test |
| IRS1 | Insulin receptor substrate 1 |
| IRS-1,2 | insulin receptor-1,2 |
| JNK | c-Jun N-terminal kinases |
| K | potassium |
| KDa | kilodaltons |
| LDL-C | low-density lipoprotein -C |
| LPO | lipid peroxidation |
| LPS | lipopolysaccharides |
| MAPK | mitogen-activated protein kinases |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MDA | malondialdehyde |
| mRNA | messenger RNA |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium |
| Na | sodium |
| NAG | n-acetyl-β-d-glucosaminidase |
| NO | nitric oxide |
| OGTT | oral glucose tolerance test |
| OGTT | oral glucose tolerance test |
| PBG | postprandial glucose |
| PDX-1 | insulin promoter factor 1 |
| PI3K | Phosphoinositide 3-kinases |
| PK | pyruvate kinase |
| PKC | protein kinase C |
| PPAR-α | peroxisome proliferator-activated receptor-alpha |
| PYY | peptide YY hormone |
| SD | Sprague-Dawley |
| SOD | superoxide dismutase |
| SREBP-1c | sterol regulatory element binding protein -1c |
| SREBP-2 | sterol regulatory element binding protein -2 |
| STZ | streptozotocin |
| T2DM | T2DM |
| TC | total cholesterol |
| TG | triglycerides |
| TGF-β1 | transforming growth factor-β1 |
| TNF-α | tumor necrosis factor-α |
| UA | uric acid |
| γGT | gamma-glutamyltransferase |
References
- Ganesan, K.; Chung, S.K.; Vanamala, J.; Xu, B. Causal relationship between diet-induced gut microbiota changes and diabetes: A novel strategy to transplant Faecalibacterium prausnitzii in preventing diabetes. Int. J. Mol. Sci. 2018, 19, 3720. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Sharmila Banu, G.; Murugesan, A.G. Attenuation of Helicteres isora L. bark extracts on streptozotocin-induced alterations in glycogen and carbohydrate metabolism in albino rats. Hum. Exp. Toxicol. 2009, 28, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Zhang, T.; Ganesan, K.; Xu, B.; Chung, S.S.M. Isoquercetin ameliorates hyperglycemia and regulates key enzymes of glucose metabolism via insulin signaling pathway in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2018, 829, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, M.; Wu, Z.; Ganesan, K.; Khalid, S.; Chung, S.M.; Xu, B. Isoquercetin upregulates antioxidant genes, suppresses inflammatory cytokines and regulates AMPK pathway in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2019, 303, 62–69. [Google Scholar] [CrossRef]
- Sukalingam, K.; Ganesan, K.; Ponnusamy, K. Evaluation of antidiabetic activity of polyherbal formulations on type 2 diabetic patients: A single blinded randomized study. Int. J. Integ. Med. Sci. 2015, 2. [Google Scholar] [CrossRef]
- Ganesan, K.; Jayachandran, M.; Xu, B. A critical review on hepatoprotective effects of bioactive food components. Crit. Rev. Food Sci. Nutr. 2017, 58, 1165–1229. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Molecular targets of vitexin and isovitexin in cancer therapy: A critical review. Ann. N. Y. Acad. Sci. 2017, 1401, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Polyphenol-rich dry common beans (Phaseolus vulgaris L.) and their health benefits. Int. J. Mol. Sci. 2017, 18, 2331. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Polyphenol-rich lentils and their health promoting effects. Int. J. Mol. Sci. 2017, 18, 2390. [Google Scholar] [CrossRef]
- Wang, D.; Li, C.; Fan, W.; Yi, T.; Wei, A.; Ma, Y. Hypoglycemic and hypolipidemic effects of a polysaccharide from Fructus Corni in streptozotocin-induced diabetic rats. Int. J. Biol. Macromol. 2019, 133, 420–427. [Google Scholar] [CrossRef]
- Xiao, H.; Chen, C.; Li, C.; Huang, Q.; Fu, X. Physicochemical characterization, antioxidant and hypoglycemic activities of selenized polysaccharides from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 132, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.; Yu, M.; Fang, Z.; Xiao, B.; Guo, L.; Wang, W.; Li, J.; Wang, S.; Zhang, Y. Preparation of the controlled acid hydrolysates from pumpkin polysaccharides and their antioxidant and antidiabetic evaluation. Int. J. Biol. Macromol. 2019, 121, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Mao, G.-H.; Zhang, M.; Li, F.; Zou, Y.; Zhou, Y.; Zheng, W.; Zheng, D.-H.; Yang, L.-Q.; Wu, X.-Y. Anti-diabetic effects of polysaccharides from ethanol-insoluble residue of Schisandra chinensis (Turcz.) Baill on alloxan-induced diabetic mice. Chem. Res. Chin. Univ. 2012, 29, 99–102. [Google Scholar] [CrossRef]
- Liu, C.; Song, J.; Teng, M.; Zheng, X.; Li, X.; Tian, Y.; Pan, M.; Li, Y.; Lee, R.J.; Wang, D. Antidiabetic and antinephritic activities of aqueous extract of Cordyceps militaris fruit body in diet-streptozotocin-induced diabetic Sprague Dawley rats. Oxid. Med. Cell. Longev. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Sarker, M.M.R.; Yan, X.; Yang, C.; Zhao, L.; Lv, X.; Liu, B.; Zhao, C. Structural characterization and antidiabetic potential of a novel heteropolysaccharide from Grifola frondosa via IRS1/PI3K-JNK signaling pathways. Carbohydr. Polym. 2018, 198, 452–461. [Google Scholar] [CrossRef]
- Tang, H.-L.; Chen, C.; Wang, S.-K.; Sun, G.-J. Biochemical analysis and hypoglycemic activity of a polysaccharide isolated from the fruit of Lycium barbarum L. Int. J. Biol. Macromol. 2015, 77, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-T.; Rioux, L.-E.; Turgeon, S.L. Molecular weight and sulfate content modulate the inhibition of α-amylase by fucoidan relevant for type 2 diabetes management. Pharma. Nutr. 2015, 3, 108–114. [Google Scholar] [CrossRef]
- Ye, M.; Qiu, T.; Peng, W.; Chen, W.-X.; Ye, Y.-W.; Lin, Y.-R. Purification, characterization and hypoglycemic activity of extracellular polysaccharides from Lachnum calyculiforme. Carbohydr. Polym. 2011, 86, 285–290. [Google Scholar] [CrossRef]
- Wang, J.; Jin, W.; Zhang, W.; Hou, Y.; Zhang, H.; Zhang, Q. Hypoglycemic property of acidic polysaccharide extracted from Saccharina japonica and its potential mechanism. Carbohydr. Polym. 2013, 95, 143–147. [Google Scholar] [CrossRef]
- Ma, Y.; Mao, D.; Geng, L.; Wang, Z.; Xu, C. Production, fractionation, characterization of extracellular polysaccharide from a newly isolated Trametes gibbosa and its hypoglycemic activity. Carbohydr. Polym. 2013, 96, 460–465. [Google Scholar] [CrossRef]
- Xu, W.; Zhou, Q.; Yin, J.-J.; Yao, Y.; Zhang, J.-L. Anti-diabetic effects of polysaccharides from Talinum triangulare in streptozotocin (STZ)-induced type 2 diabetic male mice. Int. J. Biol. Macromol. 2015, 72, 575–579. [Google Scholar] [CrossRef]
- Hong, T.; Zhao, J.; Dong, M.; Meng, Y.; Mu, J.; Yang, Z. Composition and bioactivity of polysaccharides from Inula britannica flower. Int. J. Biol. Macromol. 2012, 51, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lai, Q.; Zhang, J.; Huang, C.; Jia, L. Antioxidant and hypoglycemic effects of acidic-extractable polysaccharides from Cordyceps militaris on type 2 diabetes mice. Oxid. Med. Cell. Longev. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Huang, Y.; Bian, Y.; Wong, J.H.; Ng, T.B.; Wang, H. Hypoglycemic activity of the fungi Cordyceps militaris, Cordyceps sinensis, Tricholoma mongolicum, and Omphalia lapidescens in streptozotocin-induced diabetic rats. Appl. Microbiol. Biotechnol. 2006, 72, 1152–1156. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Jing, T.; Meng, Q.; Liu, C.; Hu, S.; Ma, Y.; Liu, Y.; Lu, J.; Cheng, Y.; Wang, D.; et al. Studies on the antidiabetic activities of Cordyceps militaris extract in diet-streptozotocin-induced diabetic Sprague-Dawley rats. BioMed Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, D.; You, Y.; Zeng, S.; Hu, Y.; Duan, X.; Liu, A.; Chen, H.; Hu, X.; Chen, S.; et al. Structural characterization and antidiabetic activity of a glucopyranose-rich heteropolysaccharide from Catathelasma ventricosum. Carbohydr. Polym. 2016, 149, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Li, S.P.; Zhang, G.H.; Zeng, Q.; Huang, Z.G.; Wang, Y.T.; Dong, T.T.X.; Tsim, K.W.K. Hypoglycemic activity of polysaccharide, with antioxidation, isolated from cultured Cordyceps mycelia. Phytomedicine 2006, 13, 428–433. [Google Scholar] [CrossRef]
- Yu, S.-H.; Dubey, N.K.; Li, W.-S.; Liu, M.-C.; Chiang, H.-S.; Leu, S.-J.; Shieh, Y.-H.; Tsai, F.-C.; Deng, W.-P. Cordyceps militaris treatment preserves renal function in type 2 diabetic nephropathy mice. PLoS ONE 2016, 11, e0166342. [Google Scholar] [CrossRef]
- Wang, J.; Teng, L.; Liu, Y.; Hu, W.; Chen, W.; Hu, X.; Wang, Y.; Wang, D. Studies on the antidiabetic and antinephritic activities of Paecilomyces hepiali water extract in diet-streptozotocin-induced diabetic Sprague Dawley rats. J. Diabet. Res. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Xue, J.; Tong, S.; Wang, Z.; Liu, P. Chemical characterization and hypoglycaemic activities in vitro of two polysaccharides from Inonotus obliquus by submerged culture. Molecules 2018, 23, 3261. [Google Scholar] [CrossRef]
- Huang, H.; Wang, S.-L.; Nguyen, V.; Kuo, Y.-H. Isolation and identification of potent antidiabetic compounds from Antrodia cinnamomea—An edible Taiwanese mushroom. Molecules 2018, 23, 2864. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qian, L.; Wang, B.; Zhang, Z.; Liu, H.; Zhang, Y.; Liu, J. Synergistic hypoglycemic effects of pumpkin polysaccharides and puerarin on type II diabetes mellitus mice. Molecules 2019, 24, 955. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Kim, M.; Kwon, D.; Kim, D.; Zhang, T.; Ha, C.; Park, S. Combination of aronia, red ginseng, shiitake mushroom and nattokinase potentiated insulin secretion and reduced insulin resistance with improving gut microbiome dysbiosis in insulin deficient type 2 diabetic rats. Nutrients 2018, 10, 948. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, X.; Zhao, L.Q.; Zhao, J.; Qi, Z.; Wang, L.-A. A primary study of the antioxidant, hypoglycemic, hypolipidemic, and antitumor activities of ethanol extract of brown slimecap mushroom, Chroogomphus rutilus (Agaricomycetes). Int. J. Med. Mushrooms 2017, 19, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Nyam, K.L.; Chow, C.F.; Tan, C.S.; Ng, S.T. Antidiabetic properties of the Tiger’s Milk medicinal mushroom, Lignosus rhinocerotis (Agaricomycetes), in streptozotocin-induced diabetic rats. Int. J. Med. Mushrooms 2017, 19, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Vitak, T.Y.; Wasser, S.P.; Nevo, E.; Sybirna, N.O. Enzymatic system of antioxidant protection of erythrocytes in diabetic rats treated with medicinal mushrooms Agaricus brasiliensis and Ganoderma lucidum (Agaricomycetes). Int. J. Med. Mushrooms 2017, 19, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.; Huang, Y.; Liu, Y.; Huang, M.; Song, G.; Ming, Q.; Ma, X.; Yang, J.; Deng, S.; Wen, Y.; et al. Antidiabetic activity of ergosterol from Pleurotus ostreatus in KK-Ay mice with spontaneous type 2 diabetes mellitus. Mol. Nutr. Food Res. 2018, 62, 1700444. [Google Scholar] [CrossRef]
- Ikewuchi, C.C.; Ikewuchi, J.C.; Ifeanacho, M.O. Restoration of plasma markers of liver and kidney functions/integrity in alloxan-induced diabetic rabbits by aqueous extract of Pleurotus tuberregium sclerotia. Biomed. Pharmacother. 2017, 95, 1809–1814. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Li, S.; Li, W.; Yuan, G.; Pan, Y.; Chen, H. Anti-diabetic effects of Inonotus obliquus polysaccharides in streptozotocin-induced type 2 diabetic mice and potential mechanism via PI3K-Akt signal pathway. Biomed. Pharmacother. 2017, 95, 1669–1677. [Google Scholar] [CrossRef]
- Chen, P.-H.; Weng, Y.-M.; Lin, S.-M.; Yu, Z.-R.; Wang, B.-J. Molecular weight affected antioxidant, hypoglycemic and hypotensive activities of cold water extract from Pleurotus citrinopileatus. J. Food Sci. 2017, 82, 2456–2461. [Google Scholar] [CrossRef]
- Liu, Y.; You, Y.; Li, Y.; Zhang, L.; Yin, L.; Shen, Y.; Li, C.; Chen, H.; Chen, S.; Hu, B.; et al. The characterization, selenylation and antidiabetic activity of mycelial polysaccharides from Catathelasma ventricosum. Carbohydr. Polym. 2017, 174, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Bedi, G.K.; Shri, R. In vitro and in vivo antidiabetic evaluation of selected culinary-medicinal mushrooms (Agaricomycetes). Int. J. Med. Mushrooms 2017, 19, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Y.; Sha, O.; Xu, W.; Wang, S. Hypolipidaemic and hypoglycaemic activities of polysaccharide from Pleurotus eryngii in Kunming mice. Int. J. Biol. Macromol. 2016, 93, 1206–1209. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Yin, R.; Hou, D.; Xue, Y.; Zhang, M.; Diao, X.; Zhang, Y.; Wu, J.; Hu, J.; Hu, X.; et al. The glucose-lowering effect of Foxtail millet in subjects with impaired glucose tolerance: A self-controlled clinical trial. Nutrients 2018, 10, 1509. [Google Scholar] [CrossRef] [PubMed]
- Howard, B.V.; Aragaki, A.K.; Tinker, L.F.; Allison, M.; Hingle, M.D.; Johnson, K.C.; Manson, J.E.; Shadyab, A.H.; Shikany, J.M.; Snetselaar, L.G.; et al. A low-fat dietary pattern and diabetes: A secondary analysis from the Women’s Health Initiative Dietary Modification Trial. Diabetes Care 2017, 41, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Marventano, S.; Vetrani, C.; Vitale, M.; Godos, J.; Riccardi, G.; Grosso, G. Whole grain intake and glycaemic control in healthy subjects: A systematic review and meta-analysis of randomized controlled trials. Nutrients 2017, 9, 769. [Google Scholar] [CrossRef] [PubMed]
- Mofidi, A.; Ferraro, Z.M.; Stewart, K.A.; Tulk, H.M.F.; Robinson, L.E.; Duncan, A.M.; Graham, T.E. The acute impact of ingestion of sourdough and whole-grain breads on blood glucose, insulin, and incretins in overweight and obese men. J. Nutr. Met. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Soong, Y.Y.; Quek, R.Y.C.; Henry, C.J. Glycemic potency of muffins made with wheat, rice, corn, oat and barley flours: A comparative study between in vivo and in vitro. Eur. J. Nutr. 2015, 54, 1281–1285. [Google Scholar] [CrossRef]
- Zafar, T.A.; Al-Hassawi, F.; Al-Khulaifi, F.; Al-Rayyes, G.; Waslien, C.; Huffman, F.G. Organoleptic and glycemic properties of chickpea-wheat composite breads. J. Food Sci. Technol. 2013, 52, 2256–2263. [Google Scholar] [CrossRef] [Green Version]
- Luhovyy, B.L.; Mollard, R.C.; Yurchenko, S.; Nunez, M.F.; Berengut, S.; Liu, T.T.; Smith, C.E.; Pelkman, C.L.; Anderson, G.H. The effects of whole grain high-amylose maize flour as a source of resistant starch on blood glucose, satiety, and food intake in young men. J. Food Sci. 2014, 79, H2550–H2556. [Google Scholar] [CrossRef]
- Poquette, N.M.; Gu, X.; Lee, S.-O. Grain sorghum muffin reduces glucose and insulin responses in men. Food Funct. 2014, 5, 894–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappi, J.; Aura, A.-M.; Katina, K.; Nordlund, E.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Comparison of postprandial phenolic acid excretions and glucose responses after ingestion of breads with bioprocessed or native rye bran. Food Funct. 2013, 4, 972. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Cai, X.; Xu, M.; Li, Y. Effect of oat intake on glycaemic control and insulin sensitivity: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2014, 112, 457–466. [Google Scholar] [CrossRef] [PubMed]
- He, L.-X.; Zhao, J.; Huang, Y.-S.; Li, Y. The difference between oats and beta-glucan extract intake in the management of HbA1c, fasting glucose and insulin sensitivity: A meta-analysis of randomized controlled trials. Food Funct. 2016, 7, 1413–1428. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, J.C.; Björck, I.M.E.; Nilsson, A.C. Effects of whole grain rye, with and without resistant starch type 2 supplementation, on glucose tolerance, gut hormones, inflammation and appetite regulation in an 11–14.5 hour perspective; a randomized controlled study in healthy subjects. Nutr. J. 2017, 16. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Chu, Y. Whole grain oats, more than just a fiber: Role of unique phytochemicals. Mol. Nutr. Food Res. 2017, 61, 1600715. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Brunius, C.; Lindelöf, M.; Shameh, S.A.; Wu, H.; Lee, I.; Landberg, R.; Moazzami, A.A. Targeted metabolomics reveals differences in the extended postprandial plasma metabolome of healthy subjects after intake of whole-grain rye porridges versus refined wheat bread. Mol. Nutr. Food Res. 2017, 61, 1600924. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.J.A.; Kendall, C.W.C.; Vuksan, V.; Faulkner, D.; Augustin, L.S.A.; Mitchell, S.; Ireland, C.; Srichaikul, K.; Mirrahimi, A.; Chiavaroli, L.; et al. Effect of lowering the glycemic load with Canola oil on glycemic control and cardiovascular risk factors: A randomized controlled trial. Diabetes Care 2014, 37, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Pyo, Y.-H.; Lee, K.-W. Preventive effect of monascus-fermented products enriched with ubiquinones on Type 2 diabetic rats induced by a high-fructose plus high-fat diet. J. Med. Food 2014, 17, 826–829. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Kim, M.; Chae, J.; Lee, S.-H.; Lee, J. Consumption of whole grains and legumes modulates the genetic effect of the APOA5 -1131C variant on changes in triglyceride and apolipoprotein A-V concentrations in patients with impaired fasting glucose or newly diagnosed type 2 diabetes. Trials 2014, 15, 100. [Google Scholar] [CrossRef]
- Asemi, Z.; Samimi, M.; Tabassi, Z.; Esmaillzadeh, A. The effect of DASH diet on pregnancy outcomes in gestational diabetes: A randomized controlled clinical trial. Eur. J. Clin. Nutr. 2014, 68, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Jacobs, D.R.; Pins, J.J.; Raatz, S.K.; Gross, M.D.; Slavin, J.L.; Seaquist, E.R. Effect of whole grains on insulin sensitivity in overweight hyperinsulinemic adults. Am. J. Clin. Nutr. 2002, 75, 848–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhao, Y.; Wu, Q.; John, A.; Jiang, Y.; Yang, J.; Liu, H.; Yang, B. Structure characterisation of polysaccharides in vegetable “okra” and evaluation of hypoglycemic activity. Food Chem. 2018, 242, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Hong, S.M.; Ahn, I.S.; Kim, Y.S.; Shin, D.W.; Park, S. Kochujang, a Korean fermented red pepper plus soybean paste, improves glucose homeostasis in 90% pancreatectomized diabetic rats. Nutrition 2009, 25, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, H.M.; McDonald, M.R.; Sullivan, J.A.; Tsao, R.; Platt, M.; Simpson, J.; Meckling, K.A. The effect of anthocyanin-rich purple vegetable diets on metabolic syndrome in obese Zucker rats. J. Med. Food 2017, 20, 1240–1249. [Google Scholar] [CrossRef]
- Wu, T.; Luo, J.; Xu, B. In vitro antidiabetic effects of selected fruits and vegetables against glycosidase and aldose reductase. Food Sci. Nutr. 2015, 3, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Hu, J.-L.; Nie, S.-P.; Xie, M.-Y. Antidiabetic mechanism of dietary polysaccharides based on their gastrointestinal functions. J. Agric. Food Chem. 2018, 66, 4781–4786. [Google Scholar] [CrossRef]
- Kumar, G.; Sharmila Banu, G.; Ganesan Murugesan, A.; Pandian, M.R. Antihyperglycaemic and antiperoxidative effect of Helicteres isora L. bark extracts in streptozotocin-induced diabetic rats. J. Appl. Biomed. 2007, 5, 97–104. [Google Scholar] [CrossRef]
- Kumar, G.; Sharmila Banu, G.; Ganesan Murugesan, A. Effect of Helicteres isora bark extracts on heart antioxidant status and lipid peroxidation in streptozotocin diabetic rats. J. Appl. Biomed. 2008, 6, 89–95. [Google Scholar] [CrossRef]
- Wang, Q.; Wu, X.; Shi, F.; Liu, Y. Comparison of antidiabetic effects of saponins and polysaccharides from Momordica charantia L. in STZ-induced type 2 diabetic mice. Biomed. Pharmacother. 2019, 109, 744–750. [Google Scholar] [CrossRef]
- Kumar Bhateja, P.; Singh, R. Antidiabetic activity of Acacia tortilis (Forsk.) Hayne ssp. raddiana polysaccharide on streptozotocin-nicotinamide induced diabetic rats. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, H.; Chang, N.; Zhang, K. Anti-hyperglycemic effect of the polysaccharide fraction isolated from Mactra veneriformis. Front Chem. Sci. Eng. 2010, 5, 238–244. [Google Scholar] [CrossRef]
- Zhao, R.; Qiu, B.; Li, Q.; Zhang, T.; Zhao, H.; Chen, Z.; Cai, Y.; Ruan, H.; Ge, W.; Zheng, X. LBP-4a improves insulin resistance via translocation and activation of GLUT4 in OLETF rats. Food Funct. 2014, 5, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Zhang, Y.-J.; Jiang, J.-X.; Zhu, L.-Y.; Chen, P.; Li, J.; Yao, H.-Y. Studies on the extraction of pumpkin components and their biological effects on blood glucose of diabetic mice. J. Food Drug Anal. 2013, 21, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Wang, X.; Jiang, X.; Kong, F.; Wang, S.; Yan, C. Antidiabetic effects of Morus alba fruit polysaccharides on high-fat diet- and streptozotocin-induced type 2 diabetes in rats. J. Ethnopharmacol. 2017, 199, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Guo, S.; Han, J.; Wang, Q.; Zhang, X.; Wu, W. Hypoglycemic and hypolipidemic activities of MT-α-glucan and its effect on immune function of diabetic mice. Carbohydr. Polym. 2012, 89, 245–250. [Google Scholar] [CrossRef]
- Huang, H.-Y. Effect of Pleurotus tuber-regium polysaccharides supplementation on the progression of diabetes complications in obese-diabetic rats. Chin. J. Physiol. 2014, 57, 198–208. [Google Scholar] [CrossRef]
- Leslie, W.S.; Taylor, R.; Harris, L.; Lean, M.E.J. Weight losses with low-energy formula diets in obese patients with and without type 2 diabetes: Systematic review and meta-analysis. Int. J. Obesity. 2016, 41, 96–101. [Google Scholar] [CrossRef]
- Anoop, S.; Misra, A.; Bhatt, S.P.; Gulati, S.; Mahajan, H. High fasting C-peptide levels and insulin resistance in non-lean & non-obese (BMI > 19 to < 25 kg/m2) Asian Indians with type 2 diabetes are independently associated with high intra-abdominal fat and liver span. Diabet. Met. Synd. Clin. Res. Rev. 2019, 13, 708–715. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Anti-obesity effects of medicinal and edible mushrooms. Molecules 2018, 23, 2880. [Google Scholar] [CrossRef] [PubMed]
- Vasu, S.; McClenaghan, N.H.; McCluskey, J.T.; Flatt, P.R. Mechanisms of toxicity by proinflammatory cytokines in a novel human pancreatic beta-cell line, 1.1B4. Biochim. Biophy. Acta 2014, 1840, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Franz, M.J.; Boucher, J.L.; Rutten-Ramos, S.; VanWormer, J.J. Lifestyle weight-loss intervention outcomes in overweight and obese adults with type 2 diabetes: A systematic review and meta-analysis of randomized clinical trials. J. Acad. Nutr. Diet. 2015, 115, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ren, C.; Lu, G.; Mu, Z.; Cui, W.; Gao, H.; Wang, Y. Anti-diabetic effect of mulberry leaf polysaccharide by inhibiting pancreatic islet cell apoptosis and ameliorating insulin secretory capacity in diabetic rats. Int. Immunopharmacol. 2014, 22, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.-X.; Nie, S.-P.; Li, C.; Gong, D.; Xie, M.-Y. Ganoderma atrum polysaccharide improves aortic relaxation in diabetic rats via PI3K/Akt pathway. Carbohydr. Polym. 2014, 103, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Nie, S.; Li, C.; Lin, S.; Xing, M.; Li, W.; Gong, D.; Xie, M. A newly identified polysaccharide from Ganoderma atrum attenuates hyperglycemia and hyperlipidemia. Int. J. Biol. Macromol. 2013, 57, 142–150. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, B.; Yu, Y.; Chen, Q.; Huang, T.; Li, D. Ganoderma lucidum polysaccharides exert anti-hyperglycemic effect on streptozotocin-induced diabetic rats through affecting & beta-cells. Comb. Chem. High Throughput Screen. 2012, 15, 542–550. [Google Scholar] [CrossRef]
- Li, F.; Zhang, Y.; Zhong, Z. Antihyperglycemic Effect of Ganoderma lucidum polysaccharides on streptozotocin-induced diabetic mice. Int. J. Mol. Sci. 2011, 12, 6135–6145. [Google Scholar] [CrossRef]
- Zhu, H.-Y.; Chen, G.-T.; Meng, G.-L.; Xu, J.-L. Characterization of pumpkin polysaccharides and protective effects on streptozotocin-damaged islet cells. Chin. J. Natl. Med. 2015, 13, 199–207. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Xu, H.; Wang, Y.; Li, Z.; Xue, C. Fucosylated chondroitin sulphate from sea cucumber inhibits high-fat-sucrose diet-induced apoptosis in mouse pancreatic islets via down-regulating mitochondrial signaling pathway. J. Funct. Foods 2014, 7, 517–526. [Google Scholar] [CrossRef]
- Kumar, G.; Banu, G.S.; Murugesan, A.G.; Pandian, M.R. Hypoglycaemic effect of Helicteres isora bark extract in rats. J. Ethnopharmacol. 2006, 107, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Sharmila Banu, G.; Murugesan, A.G.; Rajasekara Pandian, M. Effect of Helicteres isora. Bark extracts on brain antioxidant status and lipid peroxidation in streptozotocin diabetic rats. Pharm. Biol. 2007, 45, 753–759. [Google Scholar] [CrossRef]
- Islam, T.; Ganesan, K.; Xu, B. New Insight into mycochemical profiles and antioxidant potential of edible and medicinal mushrooms: A review. Int. J. Med. Mushrooms 2019, 21, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Maheswaran, R.; Sharmila Banu, G. Antihyperlipideamic effect of Solanum trilobatum L. leaves extract on streptozotocin induced diabetic rats. Asian J. Biomed. Pharma. Sci. 2013, 3, 51–57. [Google Scholar]
- Zhang, T.; Jayachandran, M.; Ganesan, K.; Xu, B. Black Truffle aqueous extract attenuates oxidative stress and inflammation in STZ-induced hyperglycemic rats via Nrf2 and NF-κB pathways. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, L.; Zhang, L.; Wang, T.; Zhou, Y.; Ding, C.; Yang, R.; Wang, X.; Yu, L. Optimization of extraction and antioxidant activity of polysaccharides from Salvia miltiorrhiza Bunge residue. Int. J. Biol. Macromol. 2015, 79, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Kou, L.; Du, M.; Liu, P.; Zhang, B.; Zhang, Y.; Yang, P.; Shang, M.; Wang, X. Anti-diabetic and anti-nephritic activities of Grifola frondosa mycelium polysaccharides in diet-streptozotocin-induced diabetic rats via modulation on oxidative stress. Appl. Biochem. Biotechnol. 2018, 187, 310–322. [Google Scholar] [CrossRef]
- Chen, X.; Tang, J.; Xie, W.; Wang, J.; Jin, J.; Ren, J.; Jin, L.; Lu, J. Protective effect of the polysaccharide from Ophiopogon japonicus on streptozotocin-induced diabetic rats. Carbohydr. Polym. 2013, 94, 378–385. [Google Scholar] [CrossRef]
- Liu, Y.; Wan, L.; Xiao, Z.; Wang, J.; Wang, Y.; Chen, J. Antidiabetic activity of polysaccharides from tuberous root of Liriope spicata var. prolifera in KKAy Mice. Evid. Based Complement. Alternat. Med. 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Chen, C.; Huang, Q.; Li, C.; Fu, X. Hypoglycemic effects of a Fructus mori polysaccharide in vitro and in vivo. Food Funct. 2017, 8, 2523–2535. [Google Scholar] [CrossRef]
- Sun, C.; Chen, Y.; Li, X.; Tai, G.; Fan, Y.; Zhou, Y. Anti-hyperglycemic and anti-oxidative activities of ginseng polysaccharides in STZ-induced diabetic mice. Food Funct. 2014, 5, 845. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Kohli, S.; Rai, G. Antidiabetic potential of polysaccharides from the white oyster culinary-medicinal mushroom Pleurotus florida (Higher Basidiomycetes). Int. J. Med. Mushrooms 2014, 16, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.W.; Ouyang, K.H.; Zhao, J.; Chen, H.; Xiong, L.; Wang, W.J. Structural characterization and hypolipidemic effect of Cyclocarya paliurus polysaccharide in rat. Int. J. Biol. Macromol. 2016, 91, 1073–1080. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, F.; Jiang, H.; Wang, Z.; Hua, C.; Zhang, Y. Chicory (Cichorium intybus L.) polysaccharides attenuate high-fat diet-induced non-alcoholic fatty liver disease via AMPK activation. Int. J. Biol. Macromol. 2018, 118, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Li, W.; Li, J.; Li, Y.; Song, H.; Luan, Y.; Qi, H.; Ma, L.; Lu, X.; Yang, Y. Lycium barbarum polysaccharide attenuates high-fat diet-induced hepatic steatosis by up-regulating SIRT1 expression and deacetylase activity. Sci. Rep. 2016, 6, 36209. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Gong, J.; Zhao, Y.; Zhuang, X.; Ye, Y.; Huang, F.; Lin, W. Sulfated polysaccharide from Enteromorpha prolifera suppresses SREBP-1c and ACC expression to lower serum triglycerides in high-fat-diet-induced hyperlipidaemic rats. J. Funct. Foods 2018, 40, 722–728. [Google Scholar] [CrossRef]
- Nie, Y.; Luo, F.; Wang, L.; Yang, T.; Shi, L.; Li, X.; Shen, J.; Xu, W.; Guo, T.; Lin, Q. Anti-hyperlipidemic effect of rice bran polysaccharide and its potential mechanism in high-fat diet mice. Food Funct. 2017, 8, 4028–4041. [Google Scholar] [CrossRef]
- Bruce, C.R.; Hoy, A.J.; Turner, N.; Watt, M.J.; Allen, T.L.; Carpenter, K.; Cooney, G.J.; Febbraio, M.A.; Kraegen, E.W. Overexpression of carnitine palmitoyltransferase-1 in skeletal muscle is sufficient to enhance fatty acid oxidation and improve high-fat diet–induced insulin resistance. Diabetes 2009, 58, 550–558. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, Y.; Lei, L.; Li, F.; Zhang, Y.; Chen, J.; Zhao, G.; Wu, S.; Yin, R.; Ming, J. Carboxymethylation of polysaccharide from Morchella angusticepes Peck enhances its cholesterol-lowering activity in rats. Carbohydr. Polym. 2017, 172, 85–92. [Google Scholar] [CrossRef]
- Gil-Ramírez, A.; Caz, V.; Smiderle, F.R.; Martin-Hernandez, R.; Largo, C.; Tabernero, M.; Marín, F.R.; Iacomini, M.; Reglero, G.; Soler-Rivas, C. Water-soluble compounds from Lentinula edodes influencing the HMG-CoA reductase activity and the expression of genes involved in the cholesterol metabolism. J. Agric. Food Chem. 2016, 64, 1910–1920. [Google Scholar] [CrossRef]
- Park, J.; Yeom, M.; Hahm, D.H. Fucoidan improves serum lipid levels and atherosclerosis through hepatic SREBP-2-mediated regulation. J. Pharmacol. Sci. 2016, 131, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musunuru, K.; Kathiresan, S. Surprises from genetic analyses of lipid risk factors for atherosclerosis. Circ. Res. 2016, 118, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Prospective Studies Collaboration; Lewington, S.; Whitlock, G.; Clarke, R.; Sherliker, P.; Emberson, J.; Halsey, J.; Qizilbash, N.; Peto, R.; Collins, R. Blood cholesterol and vascular mortality by age, sex, and blood pressure: A meta-analysis of individual data from 61 prospective studies with 55,000 vascular deaths. Lancet 2007, 370, 1829–1839. [Google Scholar] [PubMed]
- Petroglou, D.; Kanellos, I.; Savopoulos, C.; Kaiafa, G.; Chrysochoou, A.; Skantzis, P.; Daios, S.; Hatzitolios, A.I.; Giannoglou, G. The LDL-receptor and its molecular properties: From theory to novel biochemical and pharmacological approaches in reducing LDL-cholesterol. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Farnier, M. PCSK9 inhibitors. Curr. Opin. Lipidol. 2013, 24, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, Y.; Wang, Q.; Yang, Y. Crude extracts from Lycium barbarum suppress SREBP-1c expression and prevent diet-induced fatty liver through AMPK activation. BioMed Res. Int. 2014, 2014, 196198. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, X.; Zeng, X.; Ou, Z.; Xue, M.; Gao, D.; Liu, S.; Li, X.; Yang, S. Rheum palmatum L. Attenuates High Fat Diet-Induced Hepatosteatosis by Activating AMP-Activated Protein Kinase. Am. J. Chin. Med. 2016, 44, 551–564. [Google Scholar] [CrossRef]
- Wang, C.M.; Yuan, R.S.; Zhuang, W.Y.; Sun, J.H.; Wu, J.Y.; Li, H.; Chen, J.G. Schisandra polysaccharide inhibits hepatic lipid accumulation by down-regulating the expression of SREBPs in NAFLD mice. Lipids Health Dis. 2016, 15, 195. [Google Scholar] [CrossRef]
- Huang, X.; Tang, J.; Zhou, Q.; Lu, H.; Wu, Y.; Wu, W. Polysaccharide from fuzi (FPS) prevents hypercholesterolemia in rats. Lipids Health Dis. 2010, 9, 9. [Google Scholar] [CrossRef]
- Kim, H.; Wang, Q.; Shoemaker, C.F.; Zhong, F.; Bartley, G.E.; Yokoyama, W.H. Polysaccharide gel coating of the leaves of Brasenia schreberi lowers plasma cholesterol in hamsters. J. Tradit. Complement. Med. 2014, 5, 56–61. [Google Scholar] [CrossRef]
- Jayachandran, M.; Xiao, J.; Xu, B. A critical review on health promoting benefits of edible mushrooms through gut microbiota. Int. J. Mol. Sci. 2017, 18, 1934. [Google Scholar] [CrossRef] [PubMed]
- Westwell-Roper, C.; Dai, D.L.; Soukhatcheva, G.; Potter, K.J.; van Rooijen, N.; Ehses, J.A.; Verchere, C.B. IL-1 Blockade attenuates islet amyloid polypeptide-induced proinflammatory cytokine release and pancreatic islet graft dysfunction. J. Immunol. 2011, 187, 2755–2765. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Cao, P.; Shui, W.; Yang, Q.; Tang, Z.; Zhang, Y. Angelica sinensis polysaccharide regulates glucose and lipid metabolism disorder in prediabetic and streptozotocin-induced diabetic mice through the elevation of glycogen levels and reduction of inflammatory factors. Food Funct. 2015, 6, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, G.; Yan, J.; Li, K.; Bai, Z.; Cheng, W.; Huang, K. Rehmannia glutinosa (Gaertn.) DC. polysaccharide ameliorates hyperglycemia, hyperlipemia and vascular inflammation in streptozotocin-induced diabetic mice. J. Ethnopharmacol. 2015, 164, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yan, J.; Bai, Z.; Li, K.; Huang, K. Hypoglycemic activity and potential mechanism of a polysaccharide from the loach in streptozotocin-induced diabetic mice. Carbohydr. Polym. 2015, 121, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Pang, W.; Chen, J.; Bai, S.; Zheng, Z.; Wu, X. Hypoglycemic effect of polysaccharides with different molecular weight of Pseudostellaria heterophylla. BMC Complement. Altern. Med. 2013, 13, 267. [Google Scholar] [CrossRef] [PubMed]
- Kanagasabapathy, G.; Kuppusamy, U.R.; Abd Malek, S.N.; Abdulla, M.A.; Chua, K.-H.; Sabaratnam, V. Glucan-rich polysaccharides from Pleurotus sajor-caju (Fr.) Singer prevents glucose intolerance, insulin resistance and inflammation in C57BL/6J mice fed a high-fat diet. BMC Complement. Altern. Med. 2012, 12, 261. [Google Scholar] [CrossRef]
- Guo, C.; Li, R.; Zheng, N.; Xu, L.; Liang, T.; He, Q. Anti-diabetic effect of ramulus mori polysaccharides, isolated from Morus alba L. on STZ-diabetic mice through blocking inflammatory response and attenuating oxidative stress. Int. Immunopharmacol. 2013, 16, 93–99. [Google Scholar] [CrossRef]
- Yu, W.; Chen, H.; Xiang, Z.; He, N. Preparation of polysaccharides from Ramulus mori, and their antioxidant, anti-inflammatory and antibacterial activities. Molecules 2019, 24, 856. [Google Scholar] [CrossRef]
- Zhang, J.-G.; Liu, Q.; Liu, Z.-L.; Li, L.; Yi, L.-T. Antihyperglycemic activity of Anoectochilus roxburghii polysaccharose in diabetic mice induced by high-fat diet and streptozotocin. J. Ethnopharmacol. 2015, 164, 180–185. [Google Scholar] [CrossRef]
- Hou, D.; Yousaf, L.; Xue, Y.; Hu, J.; Wu, J.; Hu, X.; Feng, N.; Shen, Q. Mung bean (Vigna radiata L.): Bioactive polyphenols, polysaccharides, peptides, and health benefits. Nutrients 2019, 11, 1238. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Yan, X.; Zhang, M.; Chang, M.; Yun, S.; Meng, J.; Liu, J.; Feng, C.-P. Regulation of RAW 264.7 cell-mediated immunity by polysaccharides from Agaricus blazei Murill via the MAPK signal transduction pathway. Food Funct. 2017, 8, 1475–1480. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Q.; Zang, Y.; Zhao, Y.; Liu, N.; Wang, Y.; Xu, X.; Liu, L.; Mei, Q. Apple polysaccharide inhibits microbial dysbiosis and chronic inflammation and modulates gut permeability in HFD-fed rats. Int. J. Biol. Macromol. 2017, 99, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhong, J.; Zhang, Q.; Qing, D.; Yan, C. Structural elucidation and bioactivities of a novel arabinogalactan from Coreopsis tinctoria. Carbohydr. Polym. 2019, 219, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Niu, X.; Liu, N.; Gao, Y.; Wang, L.; Xu, G.; Li, X.; Yang, Y. Characterization, antioxidant and hypoglycemic activities of degraded polysaccharides from blackcurrant (Ribes nigrum L.) fruits. Food Chem. 2018, 243, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Chen, F.; Xiao, T.; Zhang, L. Inhibitory effects of polysaccharide from Diaphragma juglandis fructus on α-amylase and α-d-glucosidase activity, streptozotocin-induced hyperglycemia model, advanced glycation end-products formation, and H2O2-induced oxidative damage. Int. J. Biol. Macromol. 2019, 124, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Senthil, S.; Chandrasekaran, R.; Arjun, H.A.; Anantharaman, P. In vitro and in silico inhibition properties of fucoidan against α-amylase and α-d-glucosidase with relevance to type 2 diabetes mellitus. Carbohydr. Polym. 2019, 209, 350–355. [Google Scholar] [CrossRef]
- Cui, J.; Gu, X.; Wang, F.; Ouyang, J.; Wang, J. Purification and structural characterization of an α-glucosidase inhibitory polysaccharide from apricot (Armeniaca sibirica L. Lam.) pulp. Carbohydr. Polym. 2015, 121, 309–314. [Google Scholar] [CrossRef]
- Hu, J.-L.; Nie, S.-P.; Li, C.; Xie, M.-Y. In vitro effects of a novel polysaccharide from the seeds of Plantago asiatica L. on intestinal function. Int. J. Biol. Macromol. 2013, 54, 264–269. [Google Scholar] [CrossRef]
- Lakshmanasenthil, S.; Vinothkumar, T.; Geetharamani, D.; Marudhupandi, T.; Suja, G.; Sindhu, N.S. Fucoidan—A novel α-amylase inhibitor from Turbinaria ornata with relevance to NIDDM therapy. Biocatal. Agric. Biotechnol. 2014, 3, 66–70. [Google Scholar] [CrossRef]
- Vinoth Kumar, T.; Lakshmanasenthil, S.; Geetharamani, D.; Marudhupandi, T.; Suja, G.; Suganya, P. Fucoidan—A α-d-glucosidase inhibitor from Sargassum wightii with relevance to type 2 diabetes mellitus therapy. Int. J. Biol. Macromol. 2015, 72, 1044–1047. [Google Scholar] [CrossRef] [PubMed]
- Moullé, V.S.; Ghislain, J.; Poitout, V. Nutrient regulation of pancreatic β-cell proliferation. Biochimie 2017, 143, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.; Xie, C.; Luo, X.; Bao, Y.; Wu, B.; Hu, Y.; Zhong, Z.; Liu, C.; Li, M. Prevention effects and possible molecular mechanism of mulberry leaf extract and its formulation on rats with insulin-insensitivity. PLoS ONE 2016, 11, e0152728. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-Y.; Wang, Y.; Xu, D.-S.; Ruan, K.-F.; Feng, Y.; Wang, S. MDG-1, a polysaccharide from Ophiopogon japonicus exerts hypoglycemic effects through the PI3K/Akt pathway in a diabetic KKAy mouse model. J. Ethnopharmacol. 2012, 143, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.-Q.; Wang, Y.-L.; Gan, S.-R.; Chen, J.-C. Polysaccharides from Liriopes radix ameliorates hyperglycemia via various potential mechanisms in diabetic rats. J. Sci. Food Agric. 2013, 94, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.-X.; Nie, S.-P.; Tan, L.-H.; Li, C.; Gong, D.-M.; Xie, M.-Y. A polysaccharide from Ganoderma atrum Improves liver function in type 2 diabetic rats via antioxidant action and short-chain fatty acids excretion. J. Agric. Food Chem. 2016, 64, 1938–1944. [Google Scholar] [CrossRef]
- Lin, W.; Wang, W.; Liao, D.; Chen, D.; Zhu, P.; Cai, G.; Kiyoshi, A. A polysaccharides from Enteromorpha prolifera improve glucose metabolism in diabetic rats. J. Diabet. Res. 2015, 2015, 1–12. [Google Scholar] [CrossRef]
- Ma, X.; Zhou, F.; Chen, Y.; Zhang, Y.; Hou, L.; Cao, X.; Wang, C. A polysaccharide from Grifola frondosa relieves insulin resistance of HepG2 cell by Akt-GSK-3 pathway. Glycoconj. J. 2014, 31, 355–363. [Google Scholar] [CrossRef]
- Zhao, C.; Liao, Z.; Wu, X.; Liu, Y.; Liu, X.; Lin, Z.; Huang, Y.; Liu, B. Isolation, purification, and structural features of a polysaccharide from Phellinus linteus and Its hypoglycemic effect in alloxan-induced diabetic mice. J. Food Sci. 2014, 79, H1002–H1010. [Google Scholar] [CrossRef]
- Hu, S.; Chang, Y.; Wang, J.; Xue, C.; Li, Z.; Wang, Y. Fucosylated chondroitin sulfate from sea cucumber in combination with rosiglitazone improved glucose metabolism in the liver of the insulin-resistant mice. Biosci. Biotechnol. Biochem. 2013, 77, 2263–2268. [Google Scholar] [CrossRef]
- Hu, S.; Xu, H.; Chen, R.; Wang, J.; Li, Z.; Xu, J. Activation of PKB and ERK, but not PI3K, is involved in fucosylated chondroitin sulphate from Acaudina molpadioides induced glucose uptake. J. Funct. Foods 2014, 10, 385–396. [Google Scholar] [CrossRef]
- Liao, Z.; Zhang, J.; Liu, B.; Yan, T.; Xu, F.; Xiao, F.; Wu, B.; Bi, K.; Jia, Y. Polysaccharide from okra (Abelmoschus esculentus (L.) Moench) improves antioxidant capacity via PI3K/AKT Pathways and Nrf2 translocation in a type 2 diabetes model. Molecules 2019, 24, 1906. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Shen, Z.; Cui, J.; Zhu, Y.; Li, Y.; Chi, Y.; Wang, J.; Wang, P. Hypoglycemic activity and mechanism of the sulfated rhamnose polysaccharides chromium(III) complex in type 2 diabetic mice. Bioorg. Chem. 2019, 88, 102942. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, D.; Wang, D.; Lai, S.; Zhong, R.; Liu, Y.; Yang, C.; Liu, B.; Sarker, M.R.; Zhao, C. Hypoglycemic activity and gut microbiota regulation of a novel polysaccharide from Grifola frondosa in type 2 diabetic mice. Food Chem. Toxicol. 2019, 126, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yang, F.; Wang, J.; Huang, H.; Huang, Y. Anti-diabetic effect mediated by Ramulus mori polysaccharides. Carbohydr. Polym. 2015, 117, 63–69. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Dietary polysaccharides exert favorable effects on glucose homeostasis and reduce insulin resistance. IR: insulin receptor, IRS: insulin receptor substrate, PI3K: phosphoinositide 3-kinase, AKT: serine/threonine-specific protein kinase, ROS: reactive oxygen species, RNS: reactive nitrogen species, LPO: lipid peroxidation.
Figure 1.
Dietary polysaccharides exert favorable effects on glucose homeostasis and reduce insulin resistance. IR: insulin receptor, IRS: insulin receptor substrate, PI3K: phosphoinositide 3-kinase, AKT: serine/threonine-specific protein kinase, ROS: reactive oxygen species, RNS: reactive nitrogen species, LPO: lipid peroxidation.

Figure 2.
Schematic representation of the action of dietary polysaccharides on the insulin signaling pathway. Dietary polysaccharides elevate the cascade signaling of IRS/PI3K/AKT/GLUT4 and inhibit GSK3/GS, eventually stimulating the synthesis of glycogen. Akt is a main mediator to activate ERK/MAPK pathways and stimulate various physiological and biochemical mechanisms, including cell differentiation, proliferation, apoptosis, and cell endurance. IRS: insulin receptor substrate, PI3K: phosphoinositide 3-kinase, PDK1/2: Phosphoinositide-dependent protein kinase 1 and 2, AKT: serine/threonine-specific protein kinase, GSK-3: glycogen synthase kinase-3, GS: glycogen synthase, GLUT4: glucose transporter type 4, ERK: extracellular-signal-regulated kinase, MAPK: mitogen-activated protein kinase.
Figure 2.
Schematic representation of the action of dietary polysaccharides on the insulin signaling pathway. Dietary polysaccharides elevate the cascade signaling of IRS/PI3K/AKT/GLUT4 and inhibit GSK3/GS, eventually stimulating the synthesis of glycogen. Akt is a main mediator to activate ERK/MAPK pathways and stimulate various physiological and biochemical mechanisms, including cell differentiation, proliferation, apoptosis, and cell endurance. IRS: insulin receptor substrate, PI3K: phosphoinositide 3-kinase, PDK1/2: Phosphoinositide-dependent protein kinase 1 and 2, AKT: serine/threonine-specific protein kinase, GSK-3: glycogen synthase kinase-3, GS: glycogen synthase, GLUT4: glucose transporter type 4, ERK: extracellular-signal-regulated kinase, MAPK: mitogen-activated protein kinase.


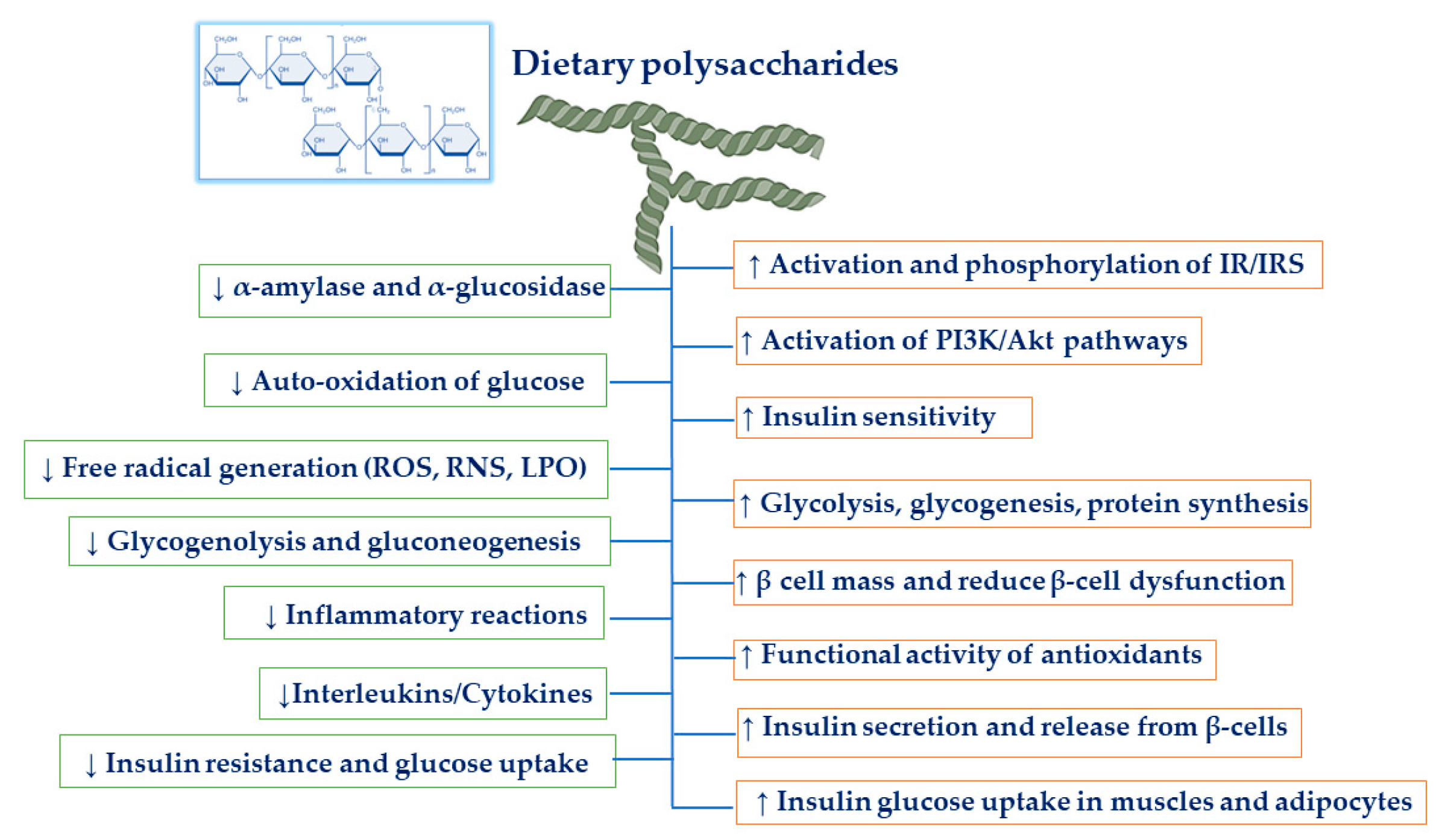{kind=link}
{kind=link}
Table 1.
In vitro and in vivo actions of dietary polysaccharides and their anti-diabetic potential.
| Source of Polysaccharides | Botanical Name/Composition | Model | Doses and Route of Administration | Negative Control | Investigation | Results | References |
|---|---|---|---|---|---|---|---|
| Mushroom | Cordyceps militaris | Wistar mice | 100 and 400 mg/kg, p.o. for 4 weeks | STZ (60 mg/kg, i.p) | FBG, Serum Insulin, OGTT, AST, ALT, BUN, CRE, LDL-C, TC, HDL-C, hepatic, renal, and pancreatic SOD, GSH-Px, CAT, and lipid peroxidation | Antioxidant and hypoglycemic effects | [23] |
| Mushroom | Cordyceps sinensis, Omphalia lapidescens, and Tricholomamongolicum | Wistar rats | 10 and 100 mg/kg, p.o. for 4 weeks | STZ (40 mg/kg, i.p) | FBG and PBG | Antioxidant and hypoglycemic effects | [24] |
| Mushroom | Cordyceps sinensis, Omphalia lapidescens, and Tricholomamongolicum. | SD male rats | 500 mg and 2000 mg/kg, p.o. for 3 weeks | STZ (40 mg/kg, i.p) | FBG, PK, SOD, GSH-Px, TG, TC, BUN, UA, CRE, and urine protein levels | Anti-diabetic and anti-nephropathic activities | [25] |
| Mushroom | Cordyceps militaris | Sprague-Dawley male rats | 0.5, 1.0, and 2.0 g/kg, p.o. for 4 weeks | STZ (40 mg/kg, i.p) | FBG, PK, SOD, GSH-Px, TG, TC, BUN, UA, CRE, urine protein, NAG, and MDA | Anti-diabetic and antinephritic activities | [26] |
| Mushroom | Cordyceps mycelia | Male BALB/c mice and male Sprague-Dawley rats | 200 mg, 400 mg/kg, p.o. for 1-week | STZ (60 mg/kg, i.p), Alloxan monohydrate (150 mg/kg, i.p.) | Blood glucose and insulin | Hypoglycemic activity | [27] |
| Mushroom | Cordyceps militaris | C57BL/6J mice | 360 mg/kg/p.o. for 8 weeks | HFD + STZ (60 mg/kg, i.p) + nicotinamide (180 mg/kg, i.p) | FBG, OGTT, IPITT, CRE, AGEs, TGF-β1, TC, TG, LDL-C, and HDL-C | Anti-diabetic and renoprotective activities | [28] |
| Mushroom | Paecilomyces hepiali | Sprague-Dawley male rats | 0.08, 0.4, and 2.0 g/kg/p.o. for 4 weeks | HFD + STZ (25 mg/kg, i.p) + nicotinamide (180 mg/kg, i.p) | Blood glucose, TC, LDL-C, insulin, PK, glycogen, SOD, MDA, GSH-Px IL-2, IL-6, IL-10, and TNF-α | Anti-diabetic and antinephritic Activities | [29] |
| Mushroom | Inonotus obliquus | HepG2 cells and insulin-resistant HepG2 cells | 10, 20, 40, 80, and 160 μg/mL, for 24 and 48 h. | - | Glucose, insulin | Hypoglycemic activity | [30] |
| Mushroom | Antrodia cinnamomea | - | 50 μL | - | α-glucosidase inhibitory activity | Anti-diabetic activity | [31] |
| Mushroom | Grifola frondosa | Male ICR mice, HepG2 | 75 and 150 mg/kg for 0, 14, and 28 days; 100 μg/mL | STZ (40 mg/kg, i.p) | Glucose, OGTT, insulin, IRS1, JNK1, PI3K, or GLUT4 | Anti-diabetic activity | [15,32] |
| Mushroom | Aronia melanocarpa, red ginseng, and shiitake mushroom | Male SD rats | 0.5, 1 g/kg bw | Pancreatectomy rats with 1 g dextrin/kg bw | Serum glucose, food intake, body weight, and OGTT | Anti-diabetic activity | [33] |
| Mushroom | Chroogomphus rutilus | Male SD rats | 1.0 and 2.0 g/kg bw, p.o. for 4 weeks | STZ (40 mg/kg, i.p) | α-glucosidase, blood glucose, SOD, GSH-Px, MDA, TC, TG, LDL-C, HDL-C, and MTT | Antioxidant, Hypoglycemic, Hypolipidemic, and Antitumor Activities | [34] |
| Mushroom | Lignosus rhinocerotis | Male SD rats | 0.5, 1.0, and 2.0 g/kg bw, p.o. for 8 weeks | STZ (35 mg/kg, i.v.) | Blood glucose, GSH, CAT, SOD, and LPO | Anti-diabetic activity | [35] |
| Mushroom | Agaricus brasiliensis and Ganoderma lucidum | Male SD rats | 1.0 and 2.0 g/kg bw, p.o. for 4 weeks | STZ (35 mg/kg, i.v.) | Blood glucose, GSH, CAT, SOD, LPO, TBARS, GSH-Px, and GSH-R | Anti-diabetic activity | [36] |
| Mushroom | Pleurotus Ostreatus | KK-Ay Mice | 1.0 and 2.0 g/kg bw, p.o. for 4 weeks | STZ (35 mg/kg, i.v.) | Blood glucose, AMPK, GLUT-4, Akt, and PKC | Anti-diabetic activity | [37] |
| Mushroom | Pleurotus Ostreatus | Rabbits | 100, 200, and 300 mg/kg for 4 weeks | Alloxan (120 mg/kg, p.o) | Blood glucose, ALP, γGT, ALT, AST, bilirubin, urea, BUN, CRE, Na, and K | Anti-diabetic activity | [38] |
| Mushroom | Inonotus obliquus | Male Kunming mic | 900 mg/kg for 4 weeks | STZ (60 mg/kg, i.p.) | Blood glucose, body weight, organ weight, glycogen, OGTT, TC, TG, LDL-C, HDL-C, PI3K, GLUT-4, and Akt | Anti-diabetic activity | [39] |
| Mushroom | Pleurotus citrinopileatus | In vitro | - | - | Pancreatic α-amylase, intestinal α-glucosidase, and ACE | Antioxidant, Hypoglycemic and Hypotensive Activities | [40] |
| Mushroom | Catathelasma ventricosum | Male ICR mice | 0.2 g/kg for 4 weeks | STZ (150 mg/kg, i.p.) | Blood glucose, TC, TG, LDL-C, and HDL-C | Anti-diabetic activity | [41] |
| Mushroom | Pleurotus ostreatus, Calocybe indica, and Volvariella volvacea | In vitro, in vivo (Male ICR mice) | 200 and 400 mg/kg for 6 weeks | STZ (150 mg/kg, i.p.) | α-amylase inhibition assay, glucose uptake by yeast cells, glucose adsorption capacity, and blood glucose | Anti-diabetic activity | [42] |
| Mushroom | Pleurotus eryngii | KKAy mice | 1 g/kg for 6 weeks | STZ (150 mg/kg, i.p.) | Blood glucose, insulin, FBS, OGTT, TC, TG, LDL-C, HDL-C, liver glycogen | Hypolipidemic and hypoglycemic activities | [43] |
| Grains | Foxtail Millet | Open-label, self-controlled clinical trial 64 subjects (27 male subjects and 37 female subjects) | 50–150 g of whole grain for week 6 and 12 | Diabetic patients | FBG, insulin, fructosamine, fasting C-peptide, TG, and TC HDL-C, LDL-C, apolipoprotein A1 and B, TNF-α, IL-6, leptin, GLP-1, blood pressure, body weight, waist circumference, and hip circumference | Anti-diabetic activity | [44] |
| Vegetable, fruit, and grain | Vegetable, fruit, and grain | 48,835 post-menopausal women | A 1:1:0.5–serving/day vegetable, fruit, food grains | Diabetic patients | Serum glucose, insulin, and waist circumference | Reduced the risk of diabetes | [45] |
| Whole Grain cereals | Whole grain cereals | A meta-analysis of randomized controlled trials | 50 g/day | Healthy Subjects | Serum glucose, insulin, and HbAlc | Improved the PBG and insulin homeostasis | [46] |
| Grain and Sprouted grain | Grain and sprouted grain | 12 male subjects | 50 g/day | Healthy Subjects | Serum glucose, insulin, and HbAlc | Only sprouted-grain improved PBG and insulin | [47] |
| Whole Grains muffins | Wheat, rice, corn, oat, and barley | 4 Male and 8 Female | 50 g/day | Healthy Subjects | Serum glucose, insulin, and HbAlc | Lowered the PBG | [48] |
| Whole grains bread | Chickpea-wheat composite bread | 13 female subjects | 50 g/day | Healthy Subjects | Serum glucose, insulin, and HbAlc | Reduced PBG | [49] |
| Whole grains bread | Maize | 30 male subjects | 50 g/day | Healthy Subjects | Serum glucose | Reduced PBG | [50] |
| Sorghum and Wheat muffin | Sorghum and wheat flour | 10 male subjects | 50 g/day | Healthy Subjects | Serum glucose, insulin | Improved the PBG and insulin | [51] |
| Whole rye bread | Whole rye with white wheat bread | 6 males and 9 females | 50 g/day | Healthy Subjects | Serum glucose, insulin | Improved the insulin response | [52] |
| Oat | Oat | A meta-analysis of randomized controlled trials | 50 g/day | Healthy Subjects | Serum glucose, insulin | Improved glucose and insulin response | [53] |
| Oat and beta-glucan | Oat and beta-glucan | A meta-analysis of randomized controlled trials | - | Healthy Subjects | Serum glucose, HbA1c, and insulin | Improved glucose and insulin and HbA1c response | [54] |
| Whole grain rye with starch | Whole grain rye flour and rye kernels bread | 21 subjects | 50 g/day | Healthy Subjects | Serum glucose, OGTT, insulin, PYY, FFA, and IL-6 | Improved cardiometabolic variables and glucose | [55] |
| Whole grain oats | Whole grain oats | A meta-analysis of randomized controlled trials | - | Healthy Subjects | Serum glucose, OGTT, insulin, and TC | Cholesterol-lowering and anti-diabetic effects | [56] |
| Whole-grain rye and wheat bread | Whole-grain rye porridges and refined wheat bread | 21 subjects | 40, 55 g/day | Healthy Subjects | Serum glucose, postprandial plasma amino acids and short chain fatty acids | Suppressed appetite and improved glucose metabolism. | [57] |
| Canola oil-enriched bread supplement | Canola oil-enriched bread | 141 subjects | 31 g/day | Diabetic patients | HbA1c, blood pressure, Framingham CVD risk score, and reactive hyperemia index ratio | Improved glycemic control in T2DM | [58] |
| Grains | Monascus-fermented grains | Male SD rats | 300 mg/kg bw. For 16 weeks | High-fructose (60%, w/w) plus high-fat (20%, w/w) diet | OGTT, Insulin, insulin sensitivity index, TBARS, SOD, CAT, and GPx | Anti-diabetic effect by improving insulin resistance and hepatic antioxidant enzymes. | [59] |
| Whole grains and legumes | Whole grains and legumes | 39 males, 146 females | 30–70 g for 16 weeks | Diabetic patients | BMI, waist and hip ratio, TC, TG, LDL-C, HDL-C, FBS, insulin FFA, Plasma apolipoprotein A-V, and CRP | Anti-diabetic effects | [60] |
| DASH diet | fruits, vegetables, whole grains, low-fat dairy products, low in saturated fats, cholesterol, refined grains, and sweets | 52 pregnant women | 40 g for 4 weeks | Gestational Diabetic patients | Length, weight, and head circumference of infants | Improved gestational diabetes mellitus | [61] |
| Whole grains | Cereal, bread, rice, pasta, and muffin | 11 subjects | 6–10 servings/day for 6 weeks | Diabetic/obese patients | Insulin, blood glucose, and OGTT | Reduce the risk of T2DM and heart disease. | [62] |
| Vegetables | Okra (Abelmoschus esculentus L. Moench) | Male C57BL/6 mice | 50 mg/kg, p.o for 10 days | STZ (45 mg/kg, i.p.) | blood glucose, OGTT | Hypoglycemic effect | [63] |
| Vegetables | Red pepper and soybeans | Male SD rats | 5% powder supplement | STZ (45 mg/kg, i.p.) | FBS, OGTT, body weight, visceral fat, and serum leptin | Improves glucose homeostasis by reducing insulin resistance | [64] |
| Fruits and vegetables | Fruits and vegetables | 550 children and adolescents | 257, 227 g/day for 30 days | Diabetic patients | FBS, insulin, and HbA1c | Anti-diabetic effect | [50] |
| Vegetables | Purple carrots and purple potatoes | Obese Zucker rats | Purple carrot and potatoes supplemented a high-fat diet for 8 weeks. | - | Intraperitoneal glucose and insulin tolerance test and invasive hemodynamic tests | Purple vegetables improve insulin resistance and hypertension | [65] |
| Apricot Lychee | Prunus armeniaca Lychee chinensis | In vitro | - | - | α-glycosidase, aldose reductase, and antioxidant activity | Anti-diabetic effects | [66] |
| Blueberry | Vaccinium cyanococcus | ||||||
| Plum | Prunus salicina | ||||||
| Kiwi | Kiwifruit c.v. hayward | ||||||
| Lemon pulp | Citrus limon | ||||||
| Lemon peel | Citrus limon | ||||||
| Pear | Pyrus bretschneider | ||||||
| Wolfberry | Lycium chinensis | ||||||
| Watermelon | Citrullus lanatusus | ||||||
| Lettuce | Lactuca sativa | In vitro | - | - | α-glycosidase, aldose reductase, and antioxidant activity | Anti-diabetic effects | [66] |
| Cucumber | Cucumis sativus | ||||||
| Red onion | Allium cepa | ||||||
| Bitter gourd | Momordica charantia | ||||||
| Eggplant | Solanum melongena | ||||||
| Celery | Apium graveolens | ||||||
| Kelp | Laminaria japonica | ||||||
| Wax gourd | Benincasa pruriens | ||||||
| Garlic | Allium sativum | ||||||
| Tomato | Solanum lycopeersicum | ||||||
| Vegetables | Momordica charantia | SD rats | 50 mg/kg, p.o for 10 days | STZ (45 mg/kg, i.p.) | FBS, insulin, and HbA1c | Anti-diabetic effects | [67] |
Abbreviations: ACE-angiotensin converting enzyme; AGEs-advanced glycation end products; Akt-serine/threonine-specific protein kinase; ALP-alkaline phosphatase; ALT-alanine transaminase; AST-aspartate transaminase; BUN-blood urea nitrogen; CAT-catalase; CRE-creatinine; CRP-C-reactive protein; FBG-fasting blood glucose; FFA-free fatty acids; GLP-1-glucagon-like peptide-1; GLUT4-glucose transporter 4; GSH-Px-glutathione peroxidase; GSH-R-glutathione reductase; HbA1c-glycated hemoglobin; HDL-C-high density lipoprotein–C; HepG2-human liver cancer cell line; HFD-high-fat diet; IL-interleukin; IL-6-interleukin-6; IPITT-intraperitoneal insulin tolerance test; IRS1-insulin receptor substrate 1; JNK-c-Jun N-terminal kinases; K-potassium; LDL-C-low density lipoprotein–C; LPO-lipid peroxidation; MDA-malondialdehyde; MTT-3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium; Na-sodium; NAG-n-acetyl-β-d-glucosaminidase; OGTT-oral glucose tolerance test; PBG-postprandial glucose; PI3K-phosphoinositide 3-kinases; PK-pyruvate kinase; PKC-protein kinase C; PYY-peptide tyrosine tyrosine hormone; SD-Sprague-Dawley; SOD-superoxide dismutase; TC-total cholesterol; T2DM-type 2 diabetes; TG-triglycerides; TGF-β1-transforming growth factor-β1; TNF-α-tumor necrosis factor-α; UA-uric acid; γGT-gamma-glutamyltransferase; i.v-intravenous; i.p-intraperitoneal; b.w-body weight; DASH-dietary approaches to stop hypertension.
Table 2.
Anti-cholesterolemic and anti-triglyceridemic effects of dietary polysaccharides.
| Sources of Polysaccharides | Monosaccharide Units/Active Compounds | Effects on Metabolism | Molecular Mechanisms | Results | References |
|---|---|---|---|---|---|
| Cyclocarya paliurus | Rhamnose, arabinose, xylose, mannose, glucose, and galactose | Triglyceride metabolism | ↑ATGL, ↑PPAR-α, ↑PPARɣ coactivator-1 α, ↓FAS, ↓HMG-CoA reductase | Anti-hyperlipidemic effects | [103] |
| Cichorium intybus L. | Sorbin, glucose, fructose, and glucitol | Triglyceride metabolism | ↑p-AMPK, ↑ATGL, ↑CAPT1, ↑p-ACC, ↓FAS, | Anti-hyperlipidemic effects | [104] |
| Lycium barbarum | Rhamnose, arabinose, xylose, mannose, glucose, galactose, and galacturonic acid | Triglyceride metabolism | ↑p-AMPK, ↑p-ACC, ↑ATGL, ↑CAPT1, ↓FAS | Anti-hyperlipidemic effects | [105] |
| Enteromorpha prolifera | Rhamnose, glucuronic acid, arabinose, fucose, xylose, and glucose | Cholesterol metabolism | ↓SREBP-2, ↓HMG-CoA reductase | Cholesterol-lowering effects | [106] |
| Oryza sativa L. | Xylose, rhamnose, mannose, galactose, arabinose, and glucose | Triglyceride and cholesterol metabolism | ↑PPAR-α, ↑PPARɣ coactivator-1 α, ↓SREBP-1c | Anti-hyperlipidemic effects | [107] |
| Morchella angusticepes | Arabinose, mannose, glucose, and galactose | Cholesterol metabolism | ↓HMG-CoA reductase | Cholesterol-lowering effects | [109] |
| Lentinula edodes | α- and β-glucans and fucomannogalactans | Cholesterol metabolism | ↓HMG-CoA reductase | Cholesterol-lowering effects | [110] |
| Fucus vesiculosus | Sulfated polysaccharide with fucose | Triglycerides and cholesterol metabolism | ↓FAS, ↓ACC, ↓SREBP -1c, ↓SREBP-2, ↓HMG-CoA reductase | Triglyceride and Cholesterol-lowering effects | [111] |
| Lycium barbarum | Rhamnose, arabinose, xylose, mannose, glucose, galactose, and galacturonic acid | Triglyceride and cholesterol metabolism | ↑p-AMPK, ↑PPARɣ coactivator-1 α, ↑p-ACC, ↓FAS, ↓SREBP-1c | Anti-hyperlipidemic effects | [116] |
| Rheum palmatum L. | Rhamnose, mannose, and galactose | Triglyceride metabolism | ↑p-AMPK, ↑p-ACC | Anti-hyperlipidemic effects | [117] |
| Schisandra Chinensis | Galactose, arabinose, and glucose | Triglyceride and cholesterol metabolism | ↓SREBP-1c, ↓SREBP-2, ↓FAS ↓ACC, ↓HMG-CoA reductase | Anti-hyperlipidemic effects | [118] |
| Aconiti Lateralis Radix Praeparata | α-d-glucan | Cholesterol metabolism | ↑LDL receptor, ↓HMG-CoA reductase | Cholesterol-lowering effects | [119] |
| Brasenia schreberi | Galactose, mannose, fucose, rhamnose, arabinose, xylose, glucose, and alduronic acids | Cholesterol metabolism | ↑LDL receptor, ↑PPAR-α | Cholesterol-lowering effects | [120] |
Abbreviations: ATGL-adipose triglyceride lipase; PPAR-α-peroxisome proliferator-activated receptor alpha; FAS-fatty acid synthase; HMG-CoA reductase-3-hydroxy-3-methylglutaryl-CoA reductase; p-AMPK-phosphorylated serine/threonine protein kinase; CAPT1-carnitine palmitoyltransferase-1; p-ACC-phosphorylated acetyl-CoA carboxylase; SREBP-2-sterol regulatory element binding protein-2; SREBP-1c-sterol regulatory element binding protein-1c, ↑ increase; ↓ decrease.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ganesan, K.; Xu, B. Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides. Molecules 2019, 24, 2556. https://doi.org/10.3390/molecules24142556
AMA Style
Ganesan K, Xu B. Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides. Molecules. 2019; 24(14):2556. https://doi.org/10.3390/molecules24142556
Chicago/Turabian StyleGanesan, Kumar, and Baojun Xu. 2019. "Anti-Diabetic Effects and Mechanisms of Dietary Polysaccharides" Molecules 24, no. 14: 2556. https://doi.org/10.3390/molecules24142556