Electrophysiology and Behavioral Assessment of the New Molecule SMe1EC2M3 as a Representative of the Future Class of Triple Reuptake Inhibitors

 ,
,  and
and
Abstract
:1. Introduction
2. Results
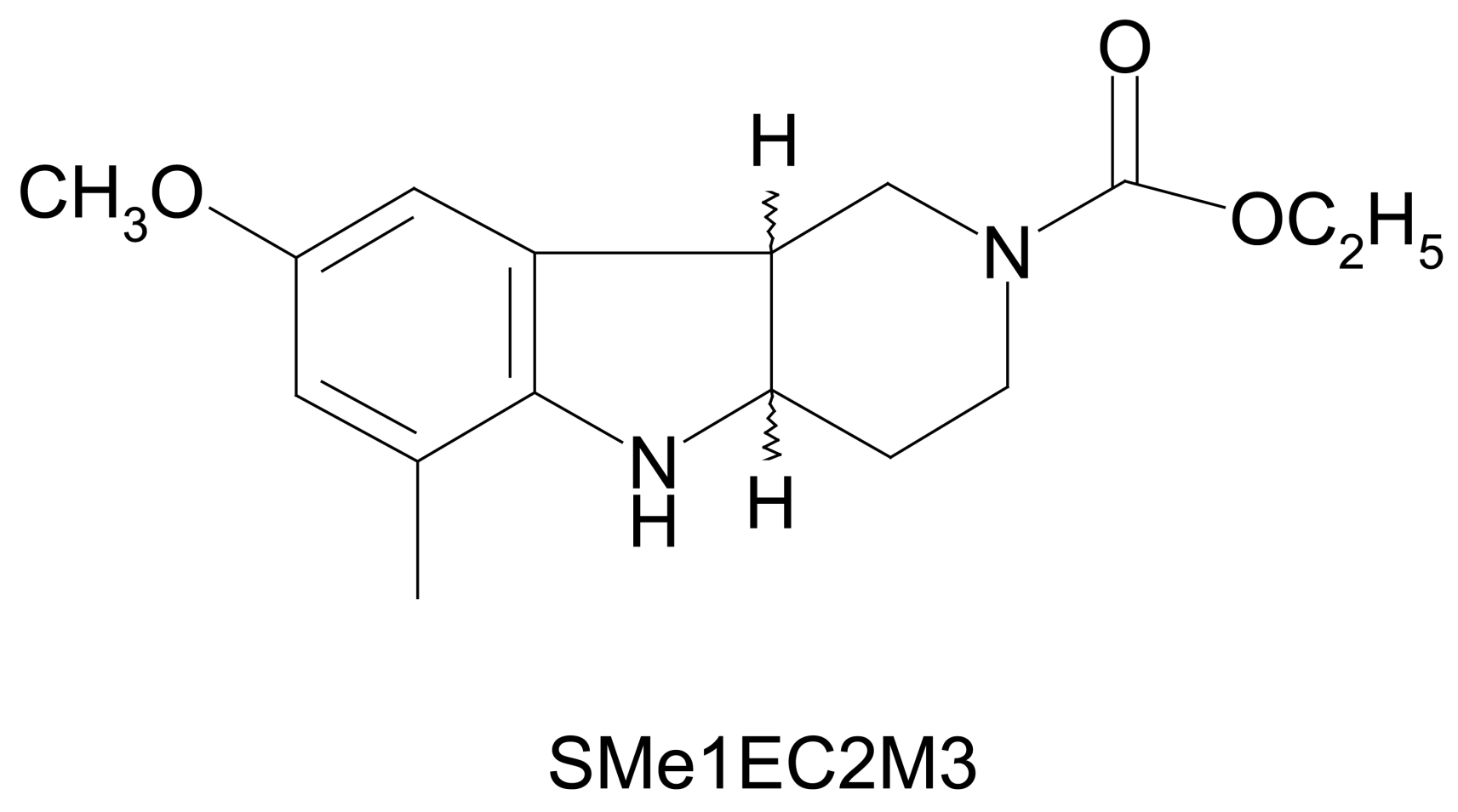
2.1. In Silico Study
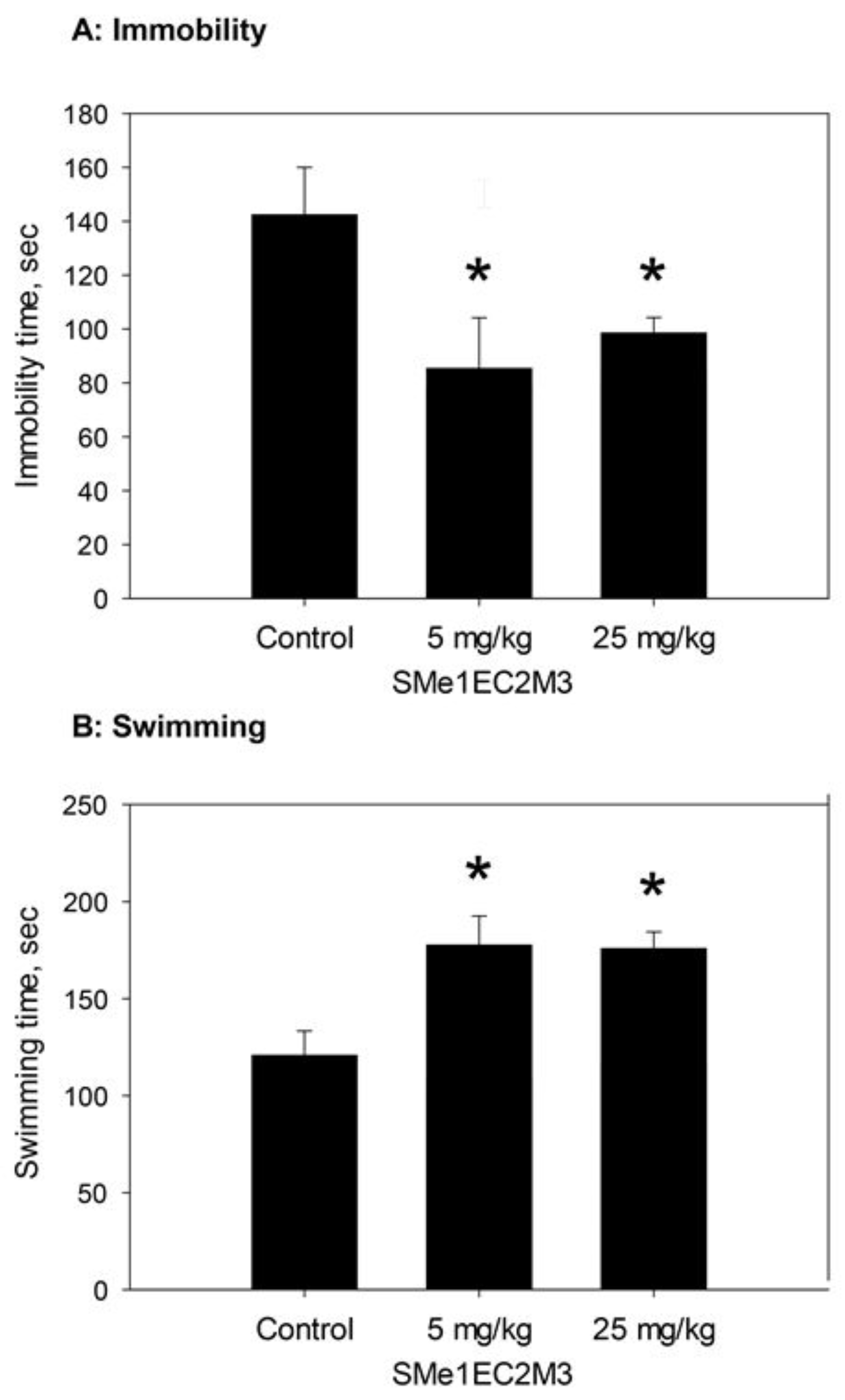
2.2. Behavioral Experiments
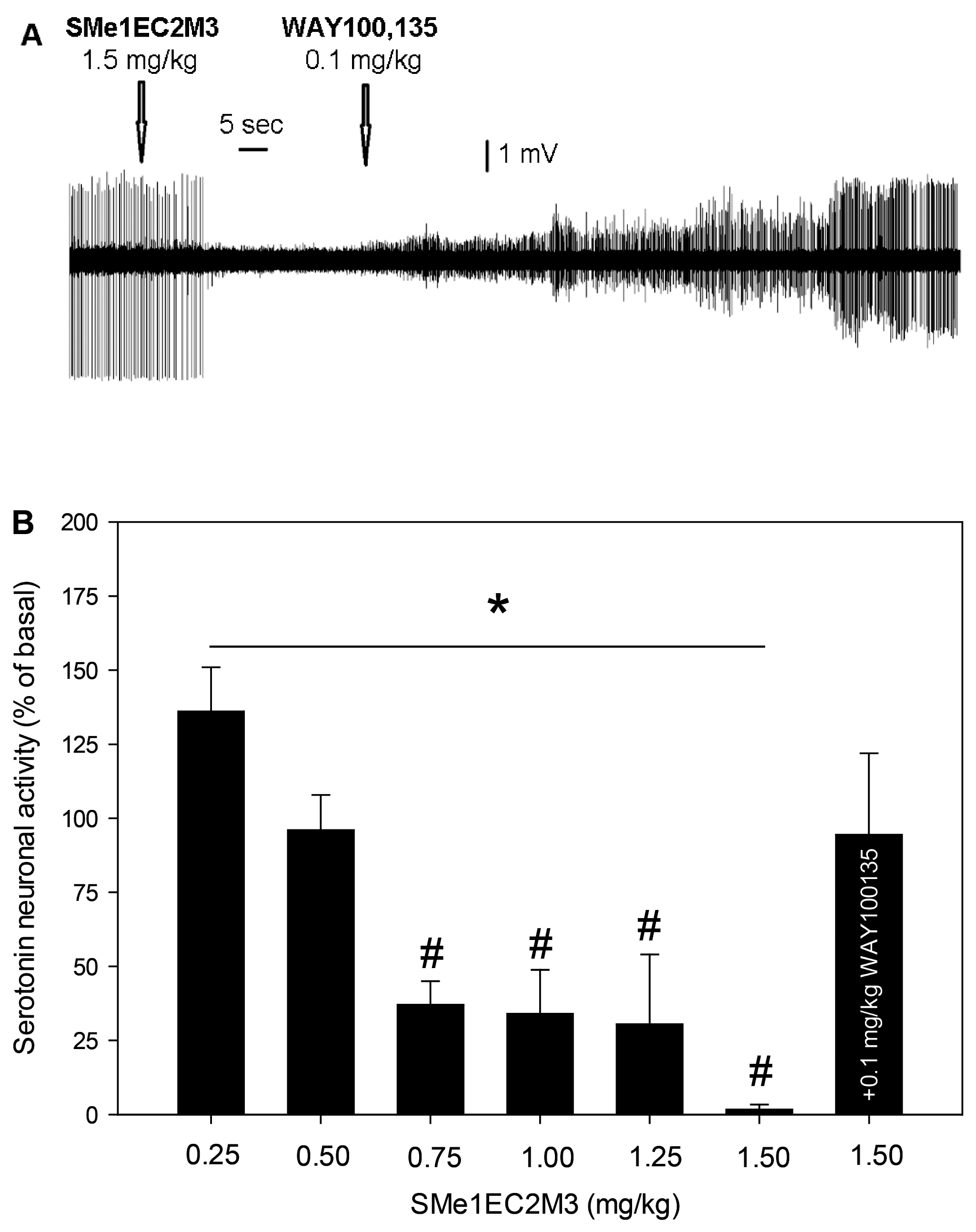
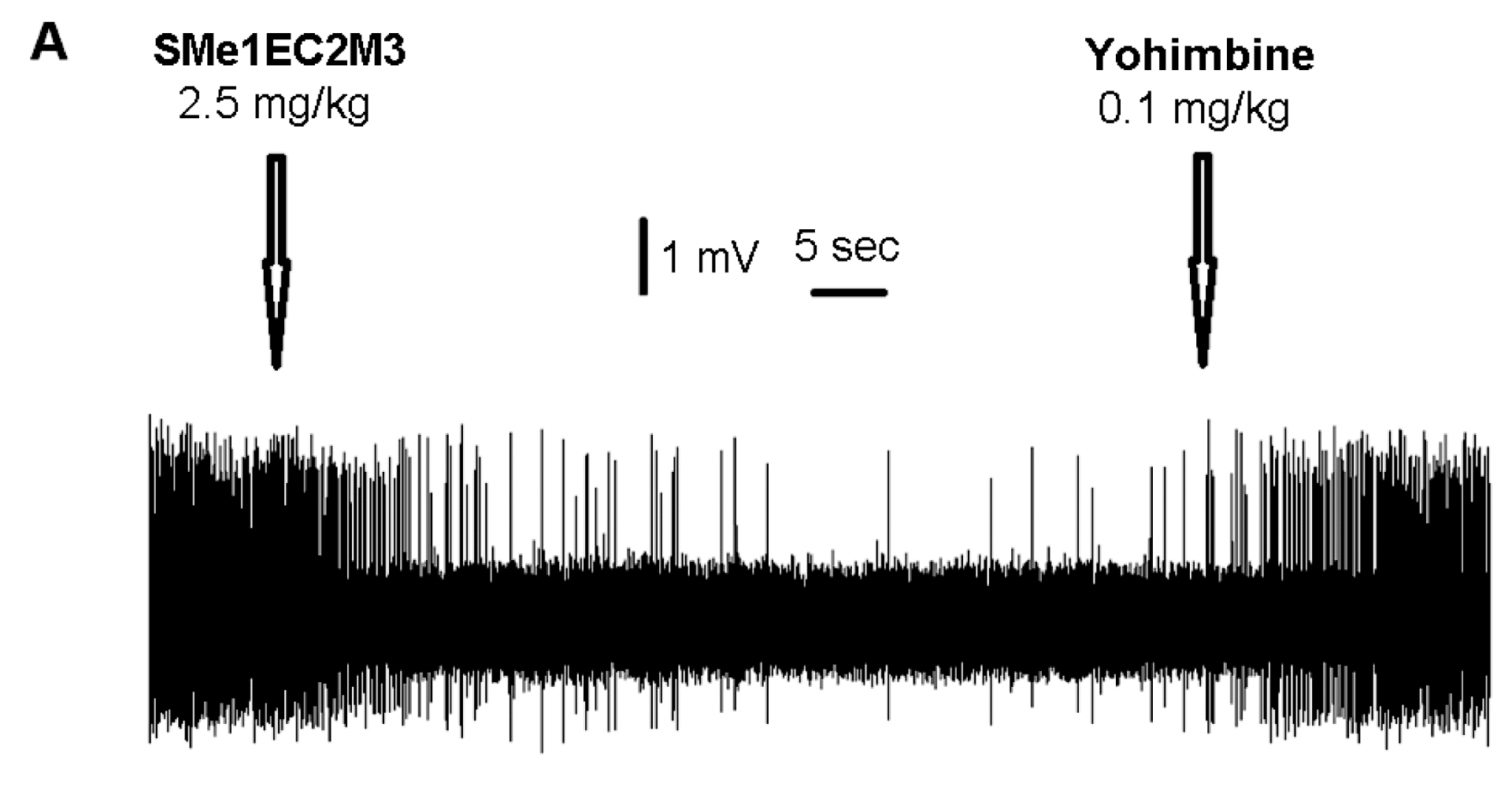
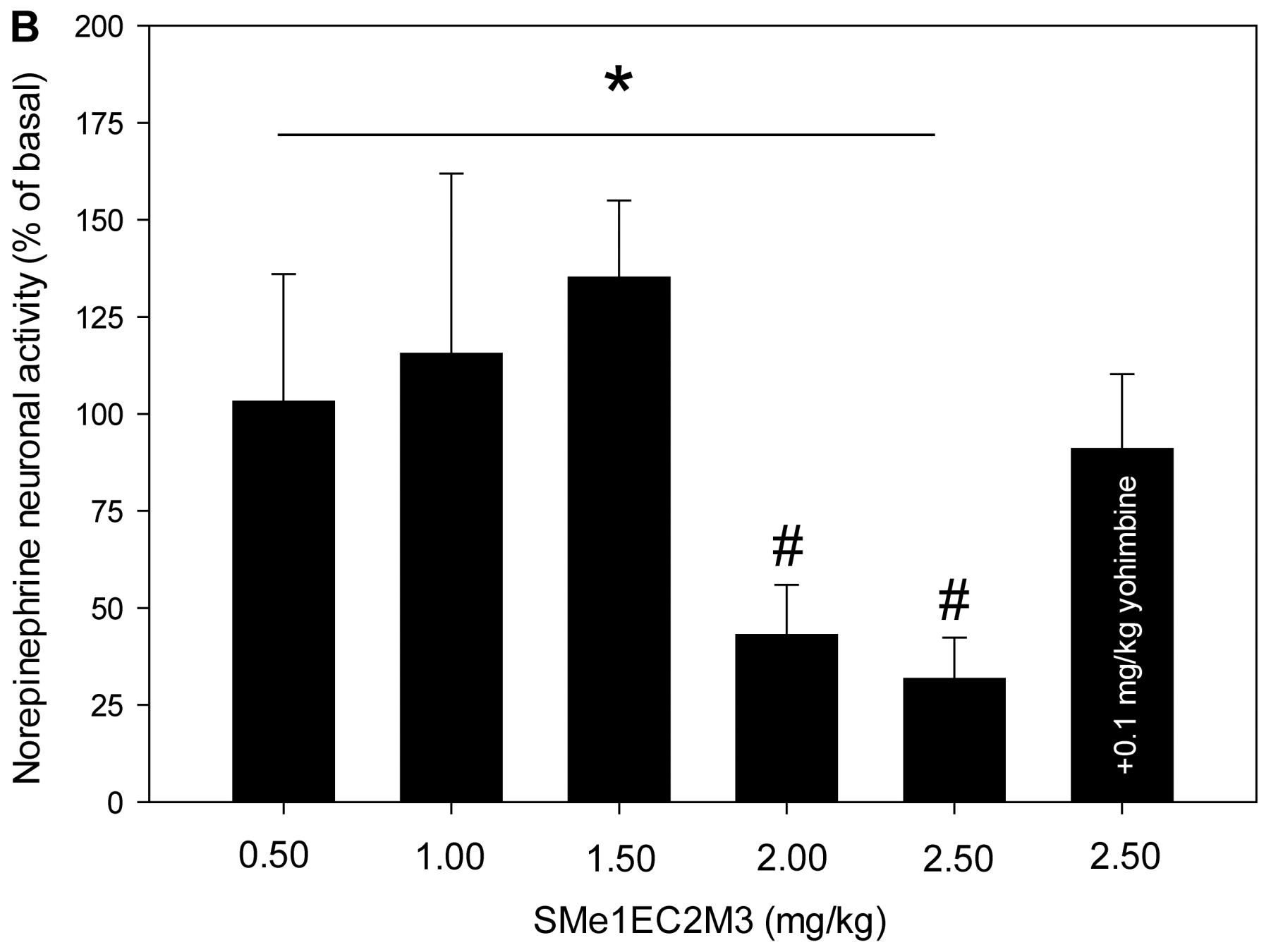
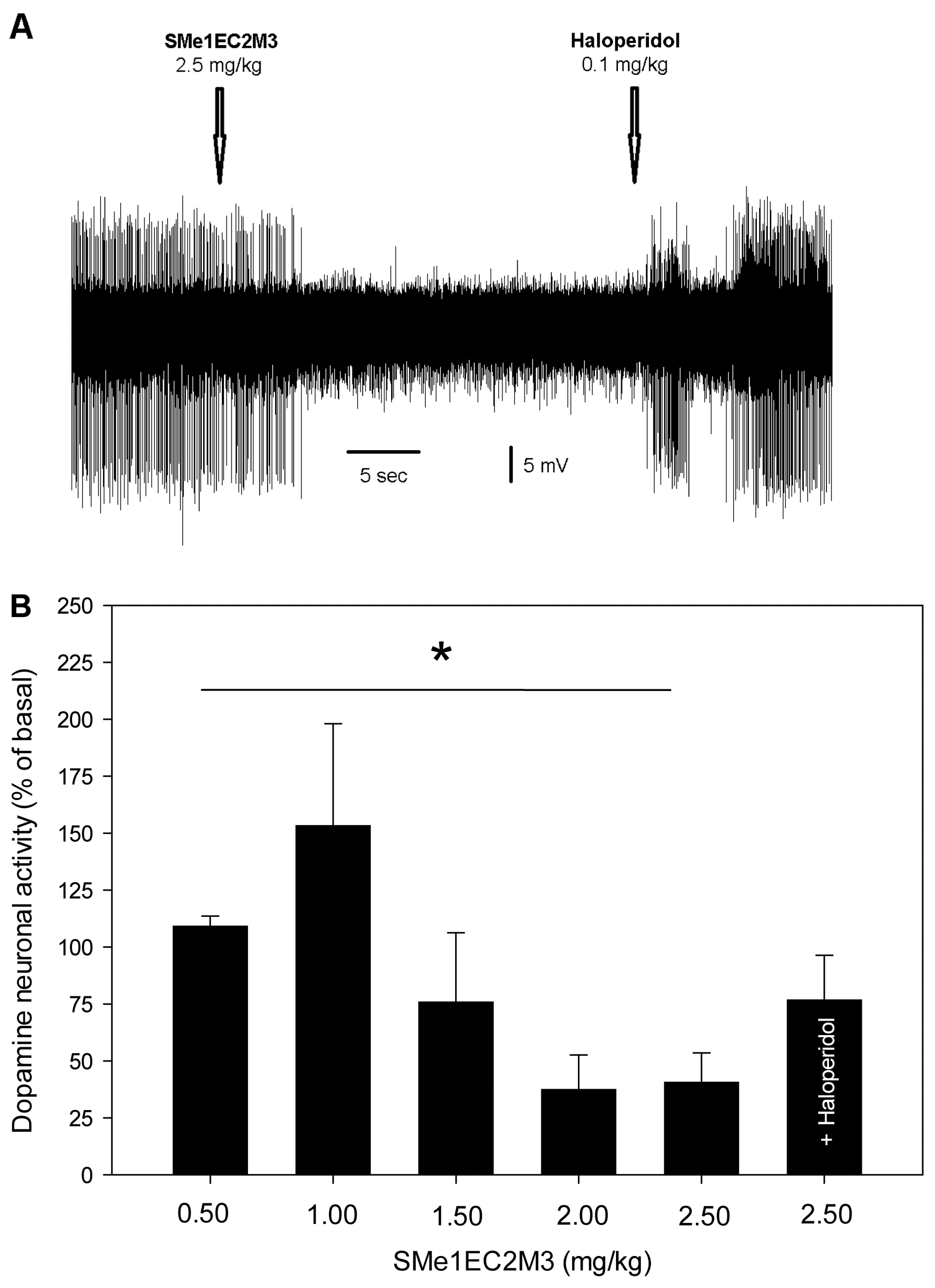
2.3. Electrophysiological Experiments
3. Discussion
4. Materials and Methods
4.1. In Silico Study
4.2. In Vivo Studies
4.2.1. Animals
4.2.2. Drugs
4.2.3. Behavioral Experiments
4.2.4. Electrophysiological Experiments
4.3. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andersen, I.; Thielen, K.; Bech, P.; Nygaard, E.; Diderichsen, F. Increasing prevalence of depression from 2000 to 2006. Scand J. Public Health 2011, 39, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.H.; Lam, R.W.; Mcintyre, R.S.; Tourjman, S.V.; Bhat, V.; Blier, P.; Hasnain, M.; Jollant, F.; Levitt, A.J.; Macqueen, G.M.; et al. Canadian network for mood and anxiety treatments (canmat) 2016 clinical guidelines for the management of adults with major depressive disorder: Section 3. pharmacological treatments. Can. J. Psychiatry 2016, 61, 540–560. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.H.; Young, A.H.; Blier, P. Strategies to achieve clinical effectiveness: Refining existing therapies and pursuing emerging targets. J. Affect Disord. 2011, 132, S21–S28. [Google Scholar] [CrossRef] [PubMed]
- Blier, P. Neurotransmitter targeting in the treatment of depression. J. Clin. Psychiatry 2013, 74, 19–24. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013; ISBN 978-08-9042-554-1. [Google Scholar]
- Liu, H.; Zakiniaeiz, Y.; Cosgrove, K.P.; Morris, E.D. Toward whole-brain dopamine movies: A critical review of pet imaging of dopamine transmission in the striatum and cortex. Brain Imaging Behav. 2019, 13, 314–322. [Google Scholar] [CrossRef]
- Guiard, B.P.; El Mansari, M.; Blier, P. Prospect of a dopamine contribution in the next generation of antidepressant drugs: The triple reuptake inhibitors. Curr. Drug Targets 2009, 10, 1069–1084. [Google Scholar] [CrossRef]
- Sharma, H.; Santra, S.; Dutta, A. Triple reuptake inhibitors as potential next-generation antidepressants: A new hope? Future Med. Chem. 2015, 7, 2385–2406. [Google Scholar] [CrossRef]
- Delorenzo, C.; Lichenstein, S.; Schaefer, K.; Dunn, J.; Marshall, R.; Organisak, L.; Kharidia, J.; Robertson, B.; Mann, J.J.; Parsey, R.V. Sep-225289 serotonin and dopamine transporter occupancy: A pet study. J. Nucl. Med. 2011, 52, 1150–1155. [Google Scholar] [CrossRef]
- Guiard, B.P.; Chenu, F.; El Mansari, M.; Blier, P. Characterization of the electrophysiological properties of triple reuptake inhibitors on monoaminergic neurons. Int. J. Neuropsychopharmacol. 2011, 14, 211–223. [Google Scholar] [CrossRef]
- Hache, G.; Guiard, B.P.; Nguyen, T.H.; Quesseveur, G.; Gardier, A.M.; Peters, D.; Munro, G.; Coudore, F. Antinociceptive activity of the new triple reuptake inhibitor ns18283 in a mouse model of chemotherapy-induced neuropathic pain. Eur. J. Pain 2015, 19, 322–333. [Google Scholar] [CrossRef]
- Štolc, S.; Považanec, F.; Bauer, V.; Májeková, M.; Wilcox, A.; Šnirc, V.; Račková, L.; Sotníková, R.; Štefek, M.; Gáspárová, Z.; et al. Pyridoindolové deriváty s antioxidačnými vlastnosťami, spôsob ich prípravy a použitia v liečebnej praxi a farmaceutické prostriedky. Industrial Property Office of the Slovak Republic. Patent 287506, 22 November 2010. [Google Scholar]
- Bachurin, S.; Bukatina, E.; Lermontova, N.; Tkachenko, S.; Afanasiev, A.; Grigoriev, V.; Grigorieva, I.; Ivanov, Y.; Sablin, S.; Zefirov, N. Antihistamine agent dimebon as a novel neuroprotector and a cognition enhancer. Ann. NY Acad. Sci. 2001, 939, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Samoylenko, V.; Rahman, M.M.; Tekwani, B.L.; Tripathi, L.M.; Wang, Y.H.; Khan, S.I.; Khan, I.A.; Miller, L.S.; Joshi, V.C.; Muhammad, I. Banisteriopsis caapi, a unique combination of mao inhibitory and antioxidative constituents for the activities relevant to neurodegenerative disorders and parkinson’s disease. J. Ethnopharmacol. 2010, 127, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Otto, R.; Penzis, R.; Gaube, F.; Winckler, T.; Appenroth, D.; Fleck, C.; Trankle, C.; Lehmann, J.; Enzensperger, C. Beta and gamma carboline derivatives as potential anti-alzheimer agents: A comparison. Eur. J. Med. Chem. 2014, 87, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Wang, F.; Malcolm, S.C.; Hewitt, M.C. Triple reuptake inhibitors and methods of their use. US Patent US8592608B2, 26 November 2013. [Google Scholar]
- Verhaar, H.J.; Solbe, J.; Speksnijder, J.; van Leeuwen, C.J.; Hermens, J.L.M. Classifying environmental pollutants: Part 3. External validation of the classification system. Chemosphere 2000, 40, 875–883. [Google Scholar] [CrossRef]
- Bogdanova, O.V.; Kanekar, S.; D’anci, K.E.; Renshaw, P.F. Factors influencing behavior in the forced swim test. Physiol. Behav. 2013, 118, 227–239. [Google Scholar] [CrossRef]
- Porsolt, R.D.; Anton, G.; Blavet, N.; Jalfre, M. Behavioural despair in rats: A new model sensitive to antidepressant treatments. Eur. J. Pharm. 1978, 47, 379–391. [Google Scholar] [CrossRef]
- Kaminska, K.; Rogoz, Z. The antidepressant- and anxiolytic-like effects following co-treatment with escitalopram and risperidone in rats. J. Physiol. Pharm. 2016, 67, 471–480. [Google Scholar]
- Koprdova, R.; Bogi, E.; Belovicova, K.; Sedlackova, N.; Okuliarova, M.; Ujhazy, E.; Mach, M. Chronic unpredictable mild stress paradigm in male wistar rats: Effect on anxiety- and depressive-like behavior. Neuro. Endocrinol. Lett. 2016, 37, 103–110. [Google Scholar]
- Skolnick, P.; Krieter, P.; Tizzano, J.; Basile, A.; Popik, P.; Czobor, P.; Lippa, A. Preclinical and clinical pharmacology of dov 216,303, a “triple” reuptake inhibitor. Cns. Drug Rev. 2006, 12, 123–134. [Google Scholar] [CrossRef]
- Caldarone, B.J.; Paterson, N.E.; Zhou, J.; Brunner, D.; Kozikowski, A.P.; Westphal, K.G.; Korte-Bouws, G.A.; Prins, J.; Korte, S.M.; Olivier, B.; et al. The novel triple reuptake inhibitor JZAD-IV-22 exhibits an antidepressant pharmacological profile without locomotor stimulant or sensitization properties. J. Pharm. Exp. 2010, 335, 762–770. [Google Scholar] [CrossRef]
- Zhang, R.; Li, X.; Shi, Y.; Shao, Y.; Sun, K.; Wang, A.; Sun, F.; Liu, W.; Wang, D.; Jin, J.; et al. The effects of LPM570065, a novel triple reuptake inhibitor, on extracellular serotonin, dopamine and norepinephrine levels in rats. PLoS ONE 2014, 9, E91775. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Markou, A.; Lucki, I. Assessing antidepressant activity in rodents: Recent developments and future needs. Trends Pharm. Sci. 2002, 23, 238–245. [Google Scholar] [CrossRef]
- Lucki, I. The Forced swimming test as a model for core and component behavioral effects of antidepressant drugs. Behav. Pharm. 1997, 8, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Valentino, R.J.; Lucki, I. Assessing substrates underlying the behavioral effects of antidepressants using the modified rat forced swimming test. Neurosci. Biobehav. Rev. 2005, 29, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Detke, M.J.; Lucki, I. Detection of serotonergic and noradrenergic antidepressants in the rat forced swimming test: The effects of water depth. Behav. Brain Res. 1996, 73, 43–46. [Google Scholar] [CrossRef]
- Reneric, J.P.; Lucki, I. Antidepressant behavioral effects by dual inhibition of monoamine reuptake in the rat forced swimming test. Psychopharmacol. 1998, 136, 190–197. [Google Scholar] [CrossRef]
- Guiard, B.P. New class of antidepressant drugs in the treatment of psychiatric disorders: The triple reuptake inhibitors. In Psychiatric Disorders: Trends and Developments; Uehara, T., Ed.; IntechOpen: London, UK, 2011; pp. 291–316. ISBN 978-953-307-745-1. [Google Scholar]
- Shaw, A.M.; Boules, M.; Zhang, Y.; Williams, K.; Robinson, J.; Carlier, P.R.; Richelson, E. Antidepressant-like effects of novel triple reuptake inhibitors, PRC025 and PRC050. Eur. J. Pharm. 2007, 555, 30–36. [Google Scholar] [CrossRef]
- Dremencov, E.; Csatlosova, K.; Durisova, B.; Moravcikova, L.; Lacinova, L.; Jezova, D. Effect of physical exercise and acute escitalopram on the excitability of brain monoamine neurons: In vivo electrophysiological study in rats. Int. J. Neuropsychopharmacol. 2017, 20, 585–592. [Google Scholar] [CrossRef]
- El Mansari, M.; Sanchez, C.; Chouvet, G.; Renaud, B.; Haddjeri, N. Effects of acute and long-term administration of escitalopram and citalopram on serotonin neurotransmission: An in vivo electrophysiological study in rat brain. Neuropsychopharmacology 2005, 30, 1269–1277. [Google Scholar] [CrossRef]
- Beique, J.C.; De Montigny, C.; Blier, P.; Debonnel, G. Venlafaxine: Discrepancy between in vivo 5-Ht and ne reuptake blockade and affinity for reuptake sites. Synapse 1999, 32, 198–211. [Google Scholar] [CrossRef]
- Palta, P.; Samuel, L.J.; Miller, E.R., 3rd; Szanton, S.L. Depression And Oxidative Stress: Results From A Meta-Analysis Of Observational Studies. Psychosom Med. 2014, 76, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhong, S.; Liao, X.; Chen, J.; He, T.; Lai, S.; Jia, Y. A meta-analysis of oxidative stress markers in depression. PLoS ONE 2015, 10, E0138904. [Google Scholar] [CrossRef] [PubMed]
- Stolc, S.; Snirc, V.; Majekova, M.; Gáspárová, Z.; Gajdosíková, A.; Stvrtina, S. Development of the new group of indole-derived neuroprotective drugs affecting oxidative stress. Cell Mol. Neurobiol. 2006, 26, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Tsuchiya, M.; Serbinova, E.; Packer, L.; Sies, H. Interaction of the pyridoindole stobadine with peroxyl, superoxide and chromanoxyl radicals. Biochem Pharmacol. 1993, 45, 393–400. [Google Scholar] [CrossRef]
- Yülek, F.; Or, M.; Ozoğul, C.; Isik, A.C.; Ari, N.; Stefek, M.; Bauer, V.; Karasu, C. Effects of stobadine and vitamin E in diabetes-induced retinal abnormalities: Involvement of oxidative stress. Arch. Med. Res. 2007, 38, 503–511. [Google Scholar] [CrossRef]
- Ghose, A.K.; Viswanadhan, V.N.; Wendoloski, J.J. A knowledge-based approach in designing combinatorial or medicinal chemistry libraries for drug discovery. 1. a qualitative and quantitative characterization of known drug databases. J. Comb. Chem. 1999, 1, 55–68. [Google Scholar] [CrossRef]
- Lee, S.; Lee, I.H.; Kim, H.J.; Chang, G.S.; Chung, J.E.; No, K.T. The preadme approach: Web-based program for rapid prediction of physico-chemical, drug absorption and drug-like properties. In Euro QSAR 2002-Designing Drugs and Crop Protectants: Processes Problems and Solutions; Ford, M.G., Livingstone, D., Dearden, J., van Waterbeemd, H., Eds.; Blackwell Publishing: Oxford, UK, 2003; pp. 418–420. ISBN 978-1405125161. [Google Scholar]
- Stolc, S.; Snirc, V.; Gajdosikova, A.; Gajdosik, A.; Gasparova, Z.; Ondrejickova, O.; Sotnikova, R.; Viola, A.; Rapta, P.; Jariabka, P.; et al. New pyridoindoles with antioxidant and neuroprotectve actions. In Trends in Pharmacological Research, 5th ed.; Bauer, V., Ed.; Institute of Experimental Pharmacology: Bratislava, Slovakia, 2008; pp. 118–136. ISBN 978-80-970003-7-0. [Google Scholar]
- Sedlackova, N.; Ponechalova, V.; Ujhazy, E.; Dubovicky, M.; Mach, M. Anxiolytic activity of pyridoindole derivatives sme1ec2 and sme1m2: Behavioral analysis using rat model. Interdiscip. Toxicol. 2011, 4, 211–215. [Google Scholar] [CrossRef]
- Slattery, D.A.; Cryan, J.F. Using the rat forced swim test to assess antidepressant-like activity in rodents. Nat. Protoc. 2012, 7, 1009–1014. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Academic Press: Boston, MA, USA, 2013; ISBN 978-01-2391-949-6. [Google Scholar]
- Dremencov, E.; El Mansari, M.; Blier, P. Distinct electrophysiological effects of paliperidone and risperidone on the firing activity of rat serotonin and norepinephrine neurons. Psychopharmacology 2007, 194, 63–72. [Google Scholar] [CrossRef]
- Flik, G.; Folgering, J.H.; Cremers, T.I.; Westerink, B.H.; Dremencov, E. Interaction between brain histamine and serotonin, norepinephrine, and dopamine systems: In vivo microdialysis and electrophysiology study. J. Mol. Neurosci. 2015, 56, 320–328. [Google Scholar] [CrossRef]
- Dremencov, E.; Lacinova, L.; Flik, G.; Folgering, J.H.; Cremers, T.I.; Westerink, B.H. Purinergic regulation of brain catecholamine neurotransmission: In vivo electrophysiology and microdialysis study in rats. Gen. Physiol. Biophys. 2017, 36, 431–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dremencov, E.; Nahshoni, E.; Levy, D.; Mintz, M.; Overstreet, D.H.; Weizman, A.; Yadid, G. Dimensional complexity of the neuronal activity in a rat model of depression. Neuroreport 2004, 15, 1983–1986. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.; Dremencov, E.; Crown, H.; Levy, D.; Mintz, M.; Overstreet, D.H.; Yadid, G. Variability of the mesolimbic neuronal activity in a rat model of depression. Neuroreport 2005, 16, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.; Deri, I.; Friedman, Y.; Dremencov, E.; Goutkin, S.; Kravchinsky, E.; Mintz, M.; Levi, D.; Overstreet, D.H.; Yadid, G. Decoding of dopaminergic mesolimbic activity and depressive behavior. J. Mol. Neurosci. 2007, 32, 72–79. [Google Scholar] [CrossRef]
- Friedman, A.; Friedman, Y.; Dremencov, E.; Yadid, G. Vta dopamine neuron bursting is altered in an animal model of depression and corrected by desipramine. J. Mol. Neurosci. 2008, 34, 201–209. [Google Scholar] [CrossRef]
- Dremencov, E.; El Mansari, M.; Blier, P. Effects of sustained serotonin reuptake inhibition on the firing of dopamine neurons in the rat ventral tegmental area. J. Psychiatry Neurosci. 2009, 34, 223–229. [Google Scholar]
Sample Availability: Samples of the compound SMe1EC2M3 are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted Property | Value | Source |
|---|---|---|
| BBB permeability (Cbrain/Cblood) | 0.13 | PreADMET |
| Human Intestinal Absorption (%) | 95.37 | |
| Plasma Protein Binding (%) | 80.97 | |
| SKlogD | 2.37 | |
| SKlogP | 2.37 | |
| BLTF96 (mmol/L) | −2.85 | Dragon |
| anti-depressant-50* | Yes | |
| anti-psychotic-50* | No | |
| hypnotic-50* | No |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koprdova, R.; Csatlosova, K.; Durisova, B.; Bogi, E.; Majekova, M.; Dremencov, E.; Mach, M. Electrophysiology and Behavioral Assessment of the New Molecule SMe1EC2M3 as a Representative of the Future Class of Triple Reuptake Inhibitors. Molecules 2019, 24, 4218. https://doi.org/10.3390/molecules24234218
Koprdova R, Csatlosova K, Durisova B, Bogi E, Majekova M, Dremencov E, Mach M. Electrophysiology and Behavioral Assessment of the New Molecule SMe1EC2M3 as a Representative of the Future Class of Triple Reuptake Inhibitors. Molecules. 2019; 24(23):4218. https://doi.org/10.3390/molecules24234218
Chicago/Turabian StyleKoprdova, Romana, Kristina Csatlosova, Barbora Durisova, Eszter Bogi, Magdalena Majekova, Eliyahu Dremencov, and Mojmir Mach. 2019. "Electrophysiology and Behavioral Assessment of the New Molecule SMe1EC2M3 as a Representative of the Future Class of Triple Reuptake Inhibitors" Molecules 24, no. 23: 4218. https://doi.org/10.3390/molecules24234218