
Recent Advances in Applications of Acidophilic Fungi to Produce Chemicals


1
Lignin Valorization & Nanomaterials Lab, Centre for Applied Molecular Biology (CAMB), University of the Punjab, 87-West Canal Bank Road, Thokar Niaz Baig, 53700 Lahore, Pakistan
2
Cell and Molecular Biology Lab, Department of Zoology, University of the Punjab Quaid-e Azam Campus, 54590 Lahore, Pakistan
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(4), 786; https://doi.org/10.3390/molecules24040786
Submission received: 14 June 2018
/
Revised: 28 July 2018
/
Accepted: 8 August 2018
/
Published: 22 February 2019
(This article belongs to the Special Issue Lignin for Energy, Chemicals and Materials)
Abstract
:Processing of fossil fuels is the major environmental issue today. Biomass utilization for the production of chemicals presents an alternative to simple energy generation by burning. Lignocellulosic biomass (cellulose, hemicellulose and lignin) is abundant and has been used for variety of purposes. Among them, lignin polymer having phenyl-propanoid subunits linked together either through C-C bonds or ether linkages can produce chemicals. It can be depolymerized by fungi using their enzyme machinery (laccases and peroxidases). Both acetic acid and formic acid production by certain fungi contribute significantly to lignin depolymerization. Fungal natural organic acids production is thought to have many key roles in nature depending upon the type of fungi producing them. Biological conversion of lignocellulosic biomass is beneficial over physiochemical processes. Laccases, copper containing proteins oxidize a broad spectrum of inorganic as well as organic compounds but most specifically phenolic compounds by radical catalyzed mechanism. Similarly, lignin peroxidases (LiP), heme containing proteins perform a vital part in oxidizing a wide variety of aromatic compounds with H2O2. Lignin depolymerization yields value-added compounds, the important ones are aromatics and phenols as well as certain polymers like polyurethane and carbon fibers. Thus, this review will provide a concept that biological modifications of lignin using acidophilic fungi can generate certain value added and environmentally friendly chemicals.
1. Introduction
The processing as well as the extraction of fossil fuels are the major prevailing environmental issues Therefore, it is the utmost need of the time to decrease the fossil fuels consumption as much as possible. The only reliable solution to this major issue is to replace the petroleum products with less costly and environmentally friendly (green) chemicals. Over 10 million tons of petrochemical materials (phenol and its derivatives) are generated annually. Thus, advancement is required to utilize new and natural raw substances for polyphenolic compounds biosynthesis [1]. Biomass is gaining much attention these days for being a renewable carbon source for chemicals, materials and energy production and hence acting as a resource to produce green products and replace fossil fuels that are decreasing day by day [2,3]. Among the plant derived raw materials, lignocellulosic biomass is most abundant and consists of three macromolecular constituents, cellulose, hemicellulose and lignin that constitutes plant cell walls. The quantities of each of these polymers are dependent on age, harvest season and plant species [4,5,6]. Identifying routes of production for both energy and value-added chemicals are imperative, and their idealistic pathways have been discussed in numerous reports [7,8].
1.1. Biomass Pretreatment Methods
A large proportion of lignocellulosic biomass pretreatment strategies had established that can be classified as physico-chemical, physical, chemical and biological methods [9,10]. Table 1 summarizes different pretreatment protocols for various feedstocks with their hydrolysis products, advantages and disadvantages [11].
1.2. Lignin
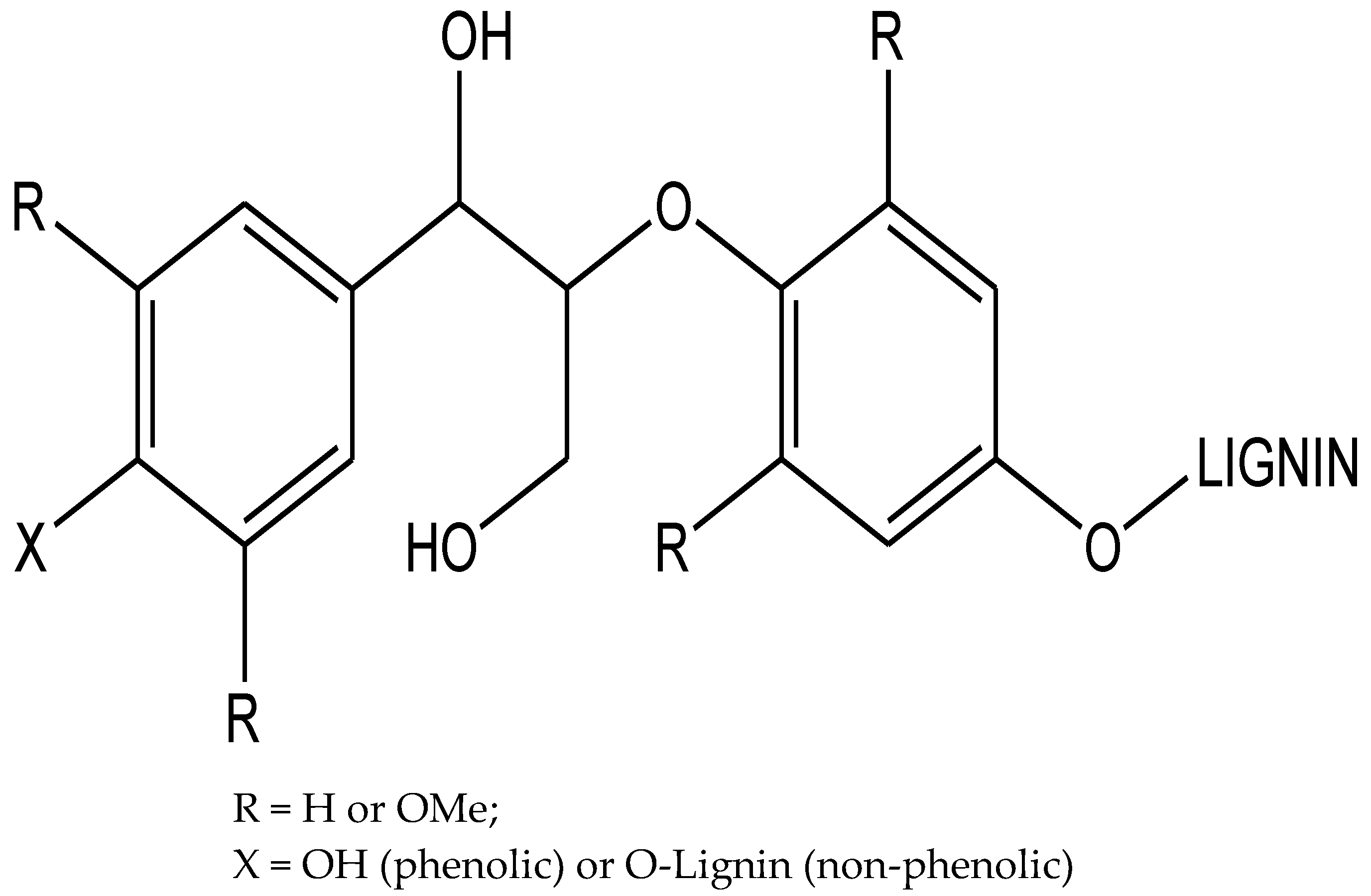
Lignin is primarily composed of three phenyl-propanoid monomers, namely sinapyl alcohol (S), 4-hydroxycinnamyl alcohol (H) and coniferyl alcohol (G) bonded by C-O or C-C linkages and is produced during cell wall biosynthesis by radical coupling reactions [23,24]. Plants utilize this high molecular weight, branched polymers for both water transport and defense. Around 50% of the inter-monomer linkages of lignin in most plants are the aryl ether β-O-4 bonds [23]. Lignin polymers are often terminated by a p-hydroxyl group, which is typically referred to as a “phenolic” group, whereas “nonphenolic” β-O-4 linkages internal to the lignin polymer can be connected to additional monomeric units (Figure 1) [25].
As the β-O-4 linkage is the most predominant in lignin, many fundamental studies have focused on understanding how this bond is cleaved in various physical and chemical environments or in the presence of homogeneous and heterogeneous catalysts [26,27]. Moreover, sophisticated NMR methods have been developed to fingerprint the presence of these aryl-ether linkages (and other linkages) in lignin to understand their fate as a function of treatment [28,29,30]. Biomass utilization efforts for both fuels and products from carbohydrates often focus on lignin removal from, or redistribution within biomass [31].
Concentrated acid hydrolysis can also be applied to depolymerize hemicellulose, cellulose, and lignin [32]. With the aim to improve carbohydrate yields from biomass, substantial efforts have also been expended to genetically modify plants to exhibit lower lignin contents or less recalcitrant lignin [33,34]. Other technologies are under development to fractionate the plant cell wall into its constituent polymers with novel solvents such as ionic liquids [35,36,37] or organic solvents (Organosolv processes) that typically utilize acid as well to depolymerize some of the lignin and hemicellulose [38,39,40].
2. Organic Acid Treatment
A broad scale laboratory investigations using a variety of organic solvents (ethanol, acetic acid, esters etc.) have been recognized to obtain remarkable results of both woody as well as non-woody pulping procedures [41,42,43].
2.1. Acetic Acid
Acetic acid, one of the first organic acids used for delignification of lignocellulosic raw material in laboratory studies, can be used as a pulping solvent uncatalyzed or catalyzed on woods [44,45]. The wood pulping properties of acetic acid is better compared with conventional chemical processes; it also possesses major benefits in contrast to other organosolv processes used at laboratory scales as reported by many researchers [46].
2.2. Formic Acid
Organosolv extraction is used as a substitute for delignification. Formic acid, a chemical agent for biomass fractionation, is readily available as well as a cheap organic solvent [47]. During formic acid pulping, lignin dissolves in black liquor due to cleavage of β-O-4 bonds of lignin, whereas solid cellulose remains in the residue after degradation of hemicellulose into both monosaccharides as well as oligosaccharides. Lignin precipitates out and separates out from the liquor by adding water. After pulping, formic acid can be easily recovered by distillation for reuse. Several techniques for biomass fractions in formic acid have been described including pulping in peroxy-formic acid mixtures, in aqueous formic acid and in acid-catalyzed aqueous formic acid [47,48,49,50].
Pulping of lignocellulose by formic acid is effective for delignification when formic acid concentration is higher than 80%, but delignification is inadequate at formic acid strength below 70% [47,51]. To establish potential applications of polymers resulting from formic acid pulping, the molecular characteristics and the structures of solid residues and lignin byproducts need to be characterized [52].
2.3. Fungal Acid Production
Low molecular weight organic acids production by filamentous fungi have attracted considerable attention due to their potential industrial applications as well as significant role in natural ecology [53,54]. Fungal natural organic acids production is thought to have many key roles in nature depending upon the type of fungi producing them. These roles are primarily either due to pH decrease consecutive to their secretion or due to direct interaction of organic acid with the environment [55,56]. The consecutive decrease in pH upon their secretion may give a competitive advantage to the acid-tolerant filamentous fungi. For ecto-mycorrhizal fungi, this decrease in pH also has been suggested to solubilize soil minerals thus releasing nutrient ions for plants and microorganisms uptake, enhancing mineral weathering [54]. For wood-decaying and saprophytic fungi, this pH acidification, caused by oxalic acid production, leads to an acid-catalyzed hydrolysis of holocellulose [57,58,59]. For this reason, Basidiomycota have been extensively studied for their ability to produce oxalic acid [60,61,62,63]. To better understand their role in the ecosystem; these studies have focused on both fungus and plant symbiosis or often growth on complex substrates [60,64]. In addressing the demand for sustainable alternatives to fossil fuels as energy source and chemicals, synthetic biology focuses on understanding how biological systems work and how to use them for the welfare of society. Organic acids can have multiple industrial applications as pharmaceutical, cosmetic excipients and food additives. They are fully degradable molecules and can be used as chemical intermediates or as synthons for biodegradable polymers synthesis; hence potentially replacing petroleum-based or synthetic chemicals [65].
A variety of useful organic acids are produced by fungi; citric, gluconic, malic and itaconic acids are synthesized by Aspergillus genera while lactic and fumaric acids are formed by Rhizopus genera. Large scale bio-processes can be used for certain organic acids like citric acid having the potential of fungi as organic acid production platforms [53,66]. Several acidophilic fungi are listed in Table 2 [67].
Lignin inhibits both enzymatic and microbial attack as it is the chief constituent of plant tissue’s mechanical support. By forming stable lignin-carbohydrate complexes (LCCs) with polysaccharides, lignin restricts the ruminal degradation and digestion of both cellulose and hemicellulose [68]. Some microorganisms, such as white-rot fungi and actinomycetes, can degrade LCCs [69,70], but depolymerization and subsequent metabolism of lignin seems unlikely under anaerobic conditions such as those in the rumen, because oxygen is thought to be essential for lignin breakdown [69,71,72]. Gaillard and Richards [73] found soluble compounds in the rumen that had the same ultraviolet and infrared spectra as lignin, and estimated that such soluble LCCs could correspond to as much as 40% of the total lignin intake. These soluble compounds might not be a direct result of the disruption of LCCs, but might originate instead from the microbial hydrolysis of surrounding structural polysaccharides on LCCs surfaces. On the other hand, direct degradation of lignin model compounds by ruminal microbes as well as the proposed pathways for their breakdown based on HPLC analysis of the end products were also reported [74,75,76]. Synthetic model compounds have been used to define the effect of the specific lignin binding structure on the degradability of plant cell walls. 4-Methylumbelliferone (4-MUF) is an analogue of lignin that fluoresces in free state. This compound could be very useful if it were incorporated into a lignin structure by a definite mode of linkage, since cleavage of the linkage could be detected by fluorescence [77].
Fungi can primarily degrade lignocellulosic biomass. However, augmenting the microbial activities is an array of soil macro-invertebrates, whose effects may range from simple comminution and dispersion of plant material to actual dissimilation of the structural polymer of lignocellulose [78,79]. Termites, being the most abundant and important of these invertebrates with their associated microbial symbionts have the capability to dissimilate a significant proportion of both cellulose (74–99%) and hemicellulose (65–87%) constituents of the ingested lignocellulosic plant material [80,81].
3. Linkages in Lignin
Lignin molecule possesses a variety of structurally correlated phenylpropanoid subunits having either C-C bonds or ether linkages known as core lignin [82]. The different linkages type in both softwood lignin and hardwood lignin together with the functional groups and their approximate proportions commonly present in a lignin macromolecule are listed in Table 3 and Table 4 [83,84].
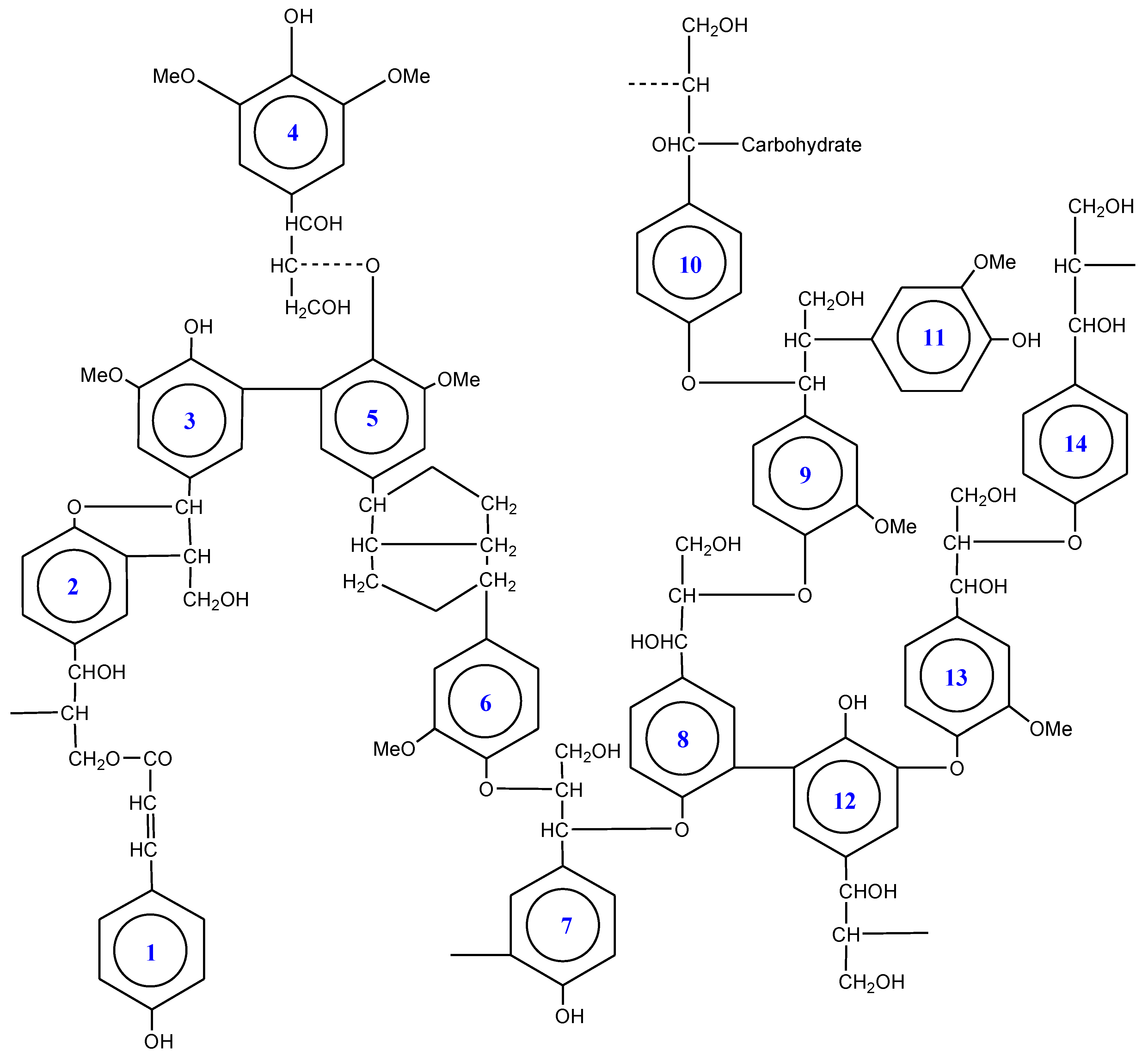
The major linkage in lignin is a phenylglycerol-β-aryl ether (e.g., ring 1→14), trailed by phenylcoumaran (ring→2), diary propane (ring→11), and biphenyl (ring→5) linkages. But diphenyl ethers (ring 12→13) and pinoresinol linkages (ring 5→6) are characteristically less common [85]. The breakdown of all these linkages by hydrolysis is difficult or not possible. The basis for the complicated, nonrepetitive structure of lignin lies in its biogenesis [86].
In 1951 Freudenberg and coworkers showed that the dehydrogenative polymerization of coniferyl alcohol yielded a high molecular weight dehydrogenation polymerizate (DHP) that closely resembled spruce lignin [87,88]. Further studies revealed the explanation of a complex reaction sequence that usually takes place in plant cell walls lignifications [82,85,89,90]. The production of majority of phenoxy radicals using extracellular peroxidases starts a new cycle of non-enzymatic polymerization reactions to form oligolignols being condensed further in parallel reactions by initiating from basic identical monomers (coniferyl, sinapyl, and p-coumaryl alcohol). A three-dimensional complex network of non-identical oligolignols constitutes the major final product, lignin. Lignin from dissimilar phylogeny has remarkable structural differences [85].
Softwood lignin possesses guaiacyl propane subunits (e.g., ring 13), being polymerizates of coniferyl alcohol monomers. Conversely, a mixture of sinapyl and coniferyl alcohol starts hardwood lignification that yields a characteristic mixture of syringyl and guaiacyl propane subunits (ring 4). Comparable to both types described, the grass lignin showed the greatest complication having 4-hydroxyphenylpropane subunits (ring 14). However, most grass lignin coupled with hardwoods have considerable percentage of chemically less recalcitrant linkages (5–10%) as aromatic acids being esterified to core lignin (ring 1→2) normally residing the primary hydroxyl groups at propyl side chains. Being covalently bonded with hemicellulose and possess carbohydrate polymer linkage, it is impossible to depolymerize lignin from lignocelluloses prior to partial denaturation. During the polymerization process in plant cell walls, many ethers and esters are formed by covalent linkages when several intermediates not only react with other oligolignols but also with glucuronic acids in hemicelluloses possessing both hydroxyl and carboxyl groups (Figure 2, ring 10) [85,86,89].
Symbiotic Fungi
A subfamily Macrotermitinae, having higher termites, plays a remarkable role by forming a fascinating symbiotic association with external basidiomycete fungi belonging to genus Termitomyces that are being cultured in greyish-brown convoluted dynamic combs. The fungal mycelium that fills these combs have plant materials being partially digested by fungus and develops mycotetes (round white nodules) consisting of many conidia (asexual spores). Plant material gets heavy with impregnation of fresh termite faeces that ultimately becomes permeated with Termitomyces spp. to develop new combs. Termites can easily utilize the older or more seasoned parts of the comb together with the fungal nodules. In 1989, researchers reviewed both biology and importance of this remarkable link that proved to be a key question for fungus role in termite nutrition [91]. Evidence suggests that Termitomyces spp. causes incomplete digestion of both plant polysaccharides and lignin within the comb [92,93].
4. Lignin Degradation
Depolymerization and aromatic ring cleavage are the key steps in lignin degradation. Certain steps are involved in oxidation of lignin due to extracellular fungal enzymes:
- Β-O-4 linkages are oxidized to arylglycerol compounds;
- Aromatic rings are cleaved that usually follows the β-ketoadipate pathway;
- Cleaved aromatic rings coupled with β-O-4 oxidation leads to the formation of cyclic carbonate structures [94].
4.1. Enzymatic Depolymerization
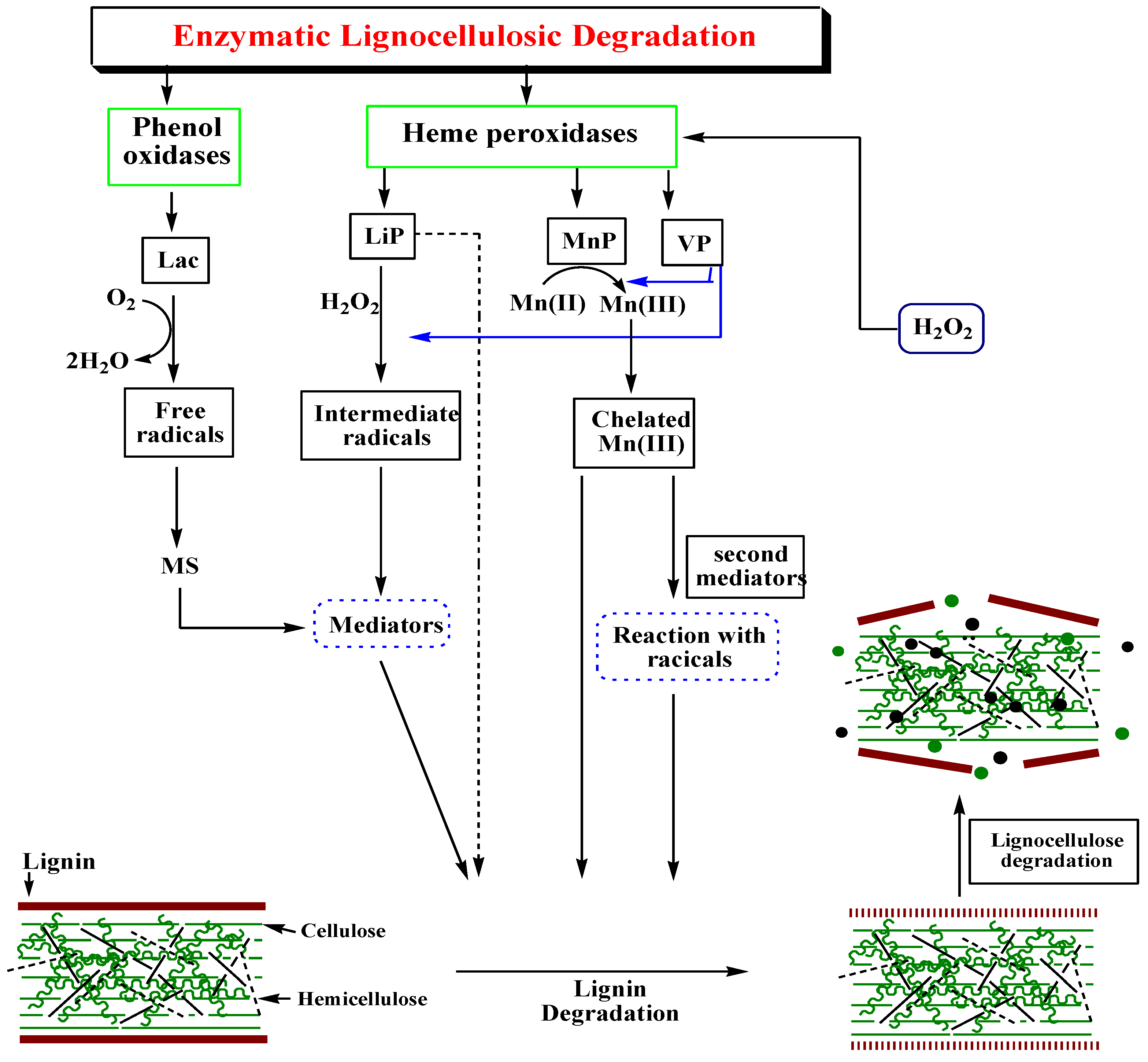
Ligninolytic enzymes that perform the conversion of lignosulphonate considered to be the main lignin degrading enzymes [95]. Enzymatic conversion of lignocellulosic is beneficial over other physiochemical processes because of enzymatic specificity in reactions. There has been an expanding literature focusing on the ligninolytic enzymes after their discovery from white rot fungi [96]. This method is a significant alternative to the other methods due to high product yield and lower environmental impact. White rot fungi produce main lignin-degrading enzymes including heme-containing lignin peroxidases (LiP), manganese peroxidase (MnP), versatile peroxidase (VP) and copper containing laccases (benzenediol: oxidoreductase) (Figure 3) [95].
4.1.1. Laccase
Laccases, being the core of interest since 19th century are one of the oldest enzymes obtained from Japanese tree, Rhus vernicifera as first extracted by Yoshida in 1883 [97]. For the first time in 1896, it was considered to be a fungal enzyme as demonstrated by Bertrand and Laborde [98]. These are the copper (Cu) containing proteins that contribute to oxidize a broad spectrum of inorganic as well as organic compounds but most specifically phenolic compounds by radical catalyzed mechanism [99].
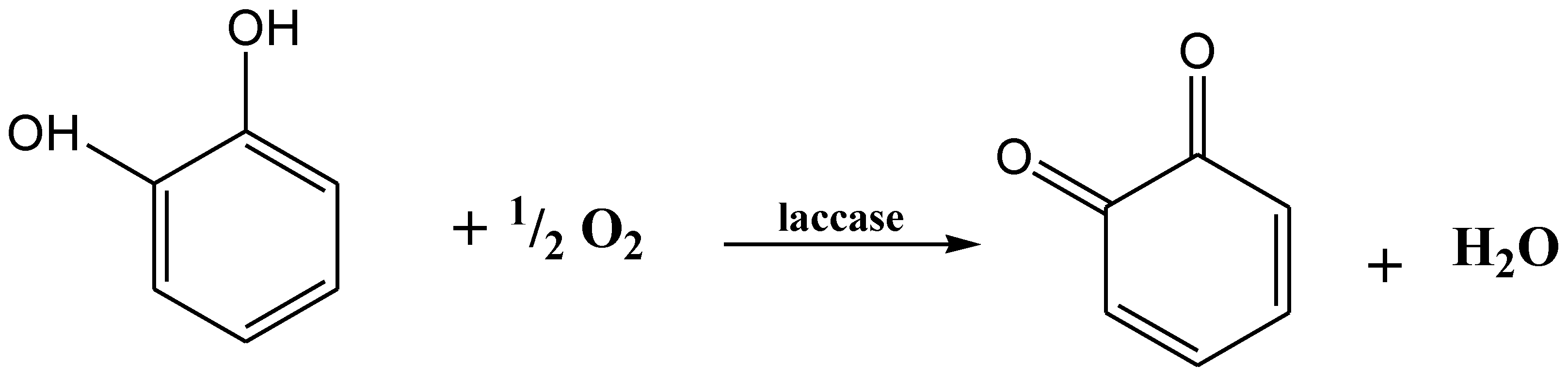
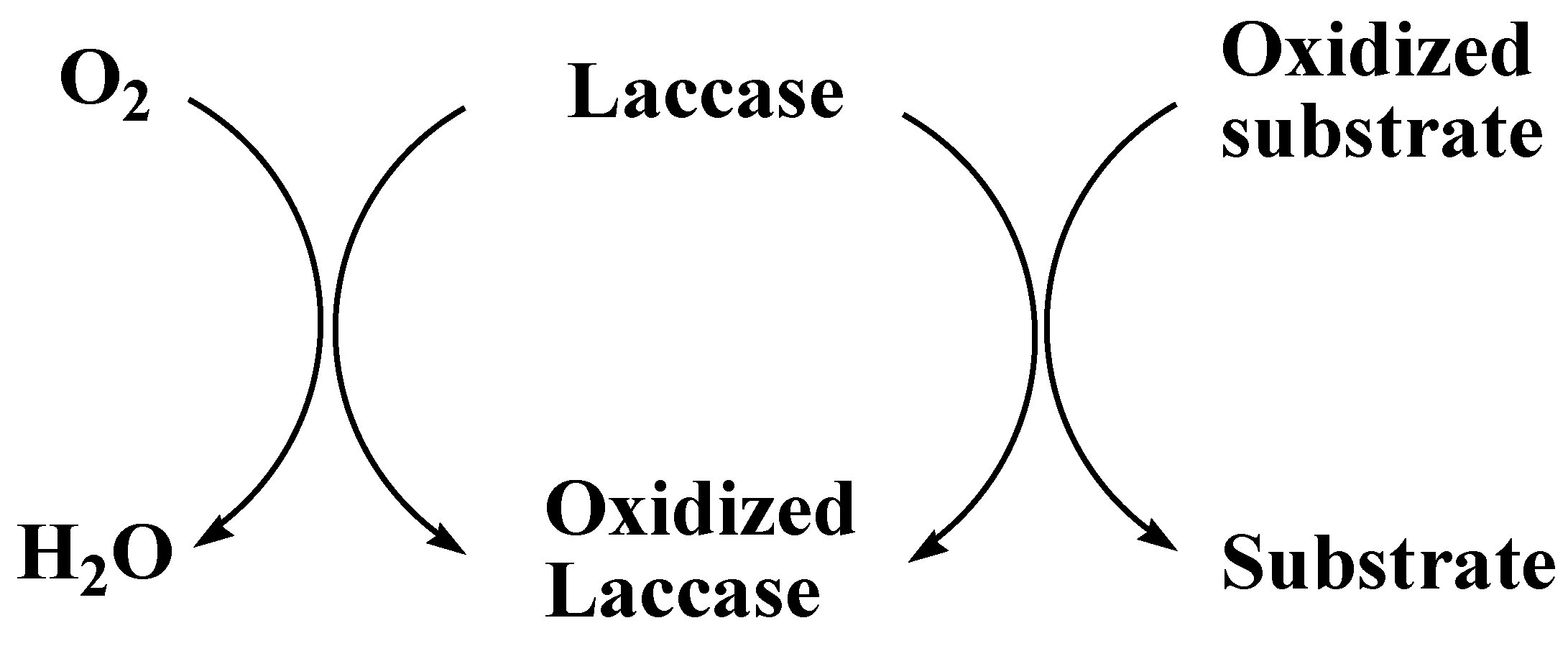
The production of enzymes has been improved by some specific compounds which act as protein synthesis inducers. The manufacturing of recombinant laccases at industrial level has been increased by the recent success in cellular engineering and fungal molecular technology. Laccases are relatively more stable because they do not use hydrogen peroxidases (H2O2) as a cofactor. They can produce water by reducing the molecular oxygen in the presence of substrate (Figure 4) [100].
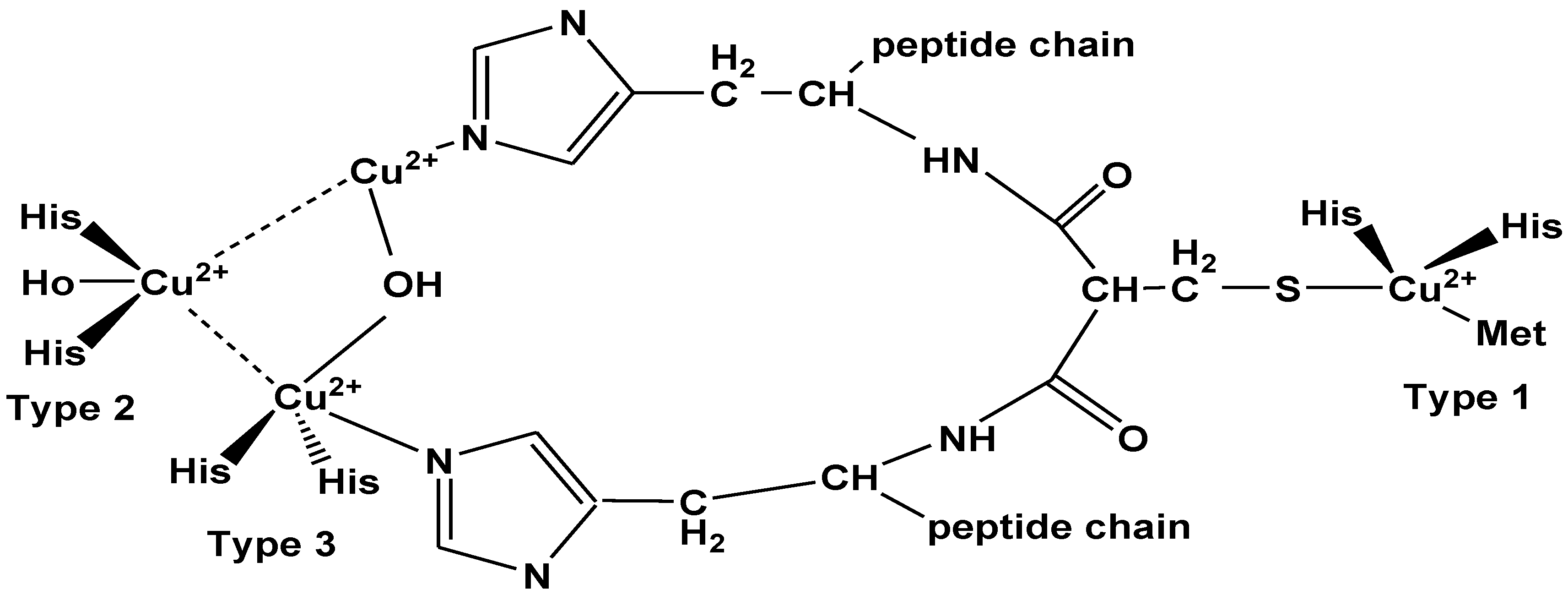
Laccases are multi-copper proteins that are characterized by their electron paramagnetic resonance (EPR) spectrum in three distinctive types:
- ▪
- Type-1 copper: attach to two amino acids (cysteine and methionine) and two histidine ligands, because of these enzymes show blue color.
- ▪
- Type-2 copper: attach via water and two histidine ligands.
- ▪
- Type-3 copper: contain two copper ions each of which attach to three histidine ligands.
Catalytic activity of laccases is performed both by type-2 and type-3 which form a trinuclear cluster (Figure 5) [101,102].
The catalytic activity is generally dependent on three binding sites with these four types of copper ions. Type-1 copper is the main primary electron acceptor and then electron transferred to the tri-nuclear cluster. The oxygen reduction into water also takes place on these binding sites. Laccases remove solely one electron to oxidize its substrate and laccase with its total reduced state contain four electrons consequently electrons gain by oxygen yielding water [103]. Substrate spontaneously forms free radical or a new compound after the removal of proton (Figure 6) [102].
An extensive amount of literature has examined the source of Laccases from fungi and plants. Its activity was also seen in bacteria viz. Streptomyces griseus, Azospirillum lipoferum, Marinomonas mediterranea, and Bacillus subtilis [104,105,106]. There are abundant types of fungi that show Laccases activity including Neurospora crassa, Pyricularia bryzae, Pleurotus, Pholiata, Polyporus versicolor A, B, and Aspergillus nidulans. However, researchers show much interest in basidiomycetes like Agaricus bisporus, Lentimus edodes, Trametes versicolor and Pleurotus ostreatus since they produce laccases that are involved in lignin degradation [107]. Laccases from Trametes versicolor (LTV) and Agaricus bisporus (LAB) are easily available commercially and have various applications in different fields including pulp and paper industry, textiles, environmental aspects, the food processing units, pharmaceutical business and nano-biotechnology [102].
Additionally, voluminous literature covers the LAB and LTV regarding their reactions and production [108]. Laccases synthesized specially from white rot fungus (LAB and LTV) can cause lignin degradation due to their ability to further rearrange the phenoxy radical by Cα-Cβ cleavage as well as the benzyl hydroxyls oxidation. Lignin polymer is too large to penetrate active site of laccase so it could not oxidize directly by laccase. Furthermore, a mediator; an additional compound is required to deal with this limitation [109].
4.1.2. Laccase-Mediator System (LMS)
For the depolymerization of lignin, laccases require a mediating agent known as intermediary substance or mediator. Mostly laccase mediators are low molecular weight and aromatic compounds. The combination of laccases with mediators increase the yields and rates in conversion of laccase-substrate as well as it adds new reactions to substrate without which enzyme shows no or just marginal activity. Consequently, LMS enhances the range of substrate to oxidize compounds with higher redox potential (E°) compared to laccases (LMS E° lies above +1100 mV but laccase allows to oxidize molecule in limited range of +475 to +790 mV) [110].
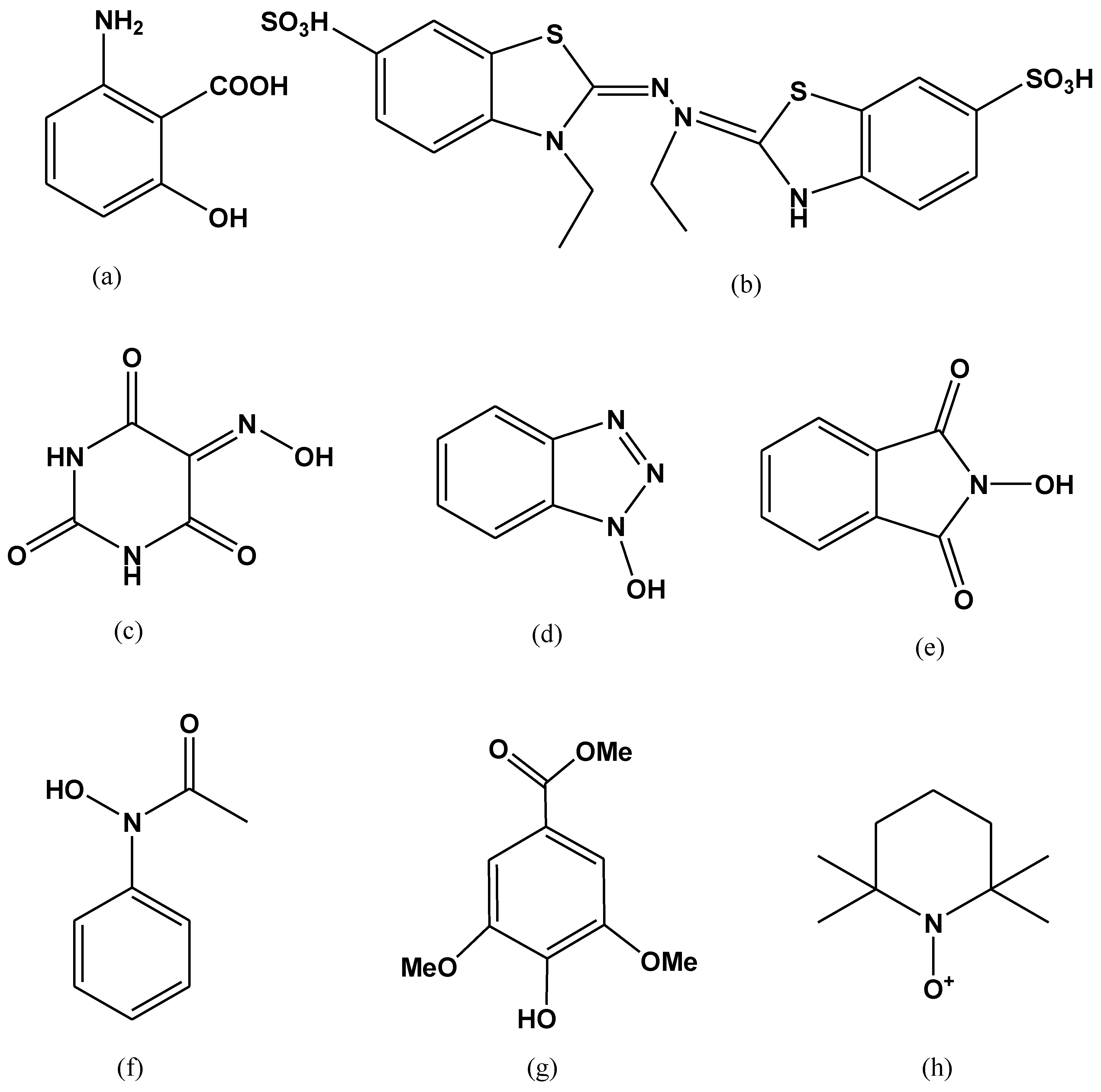
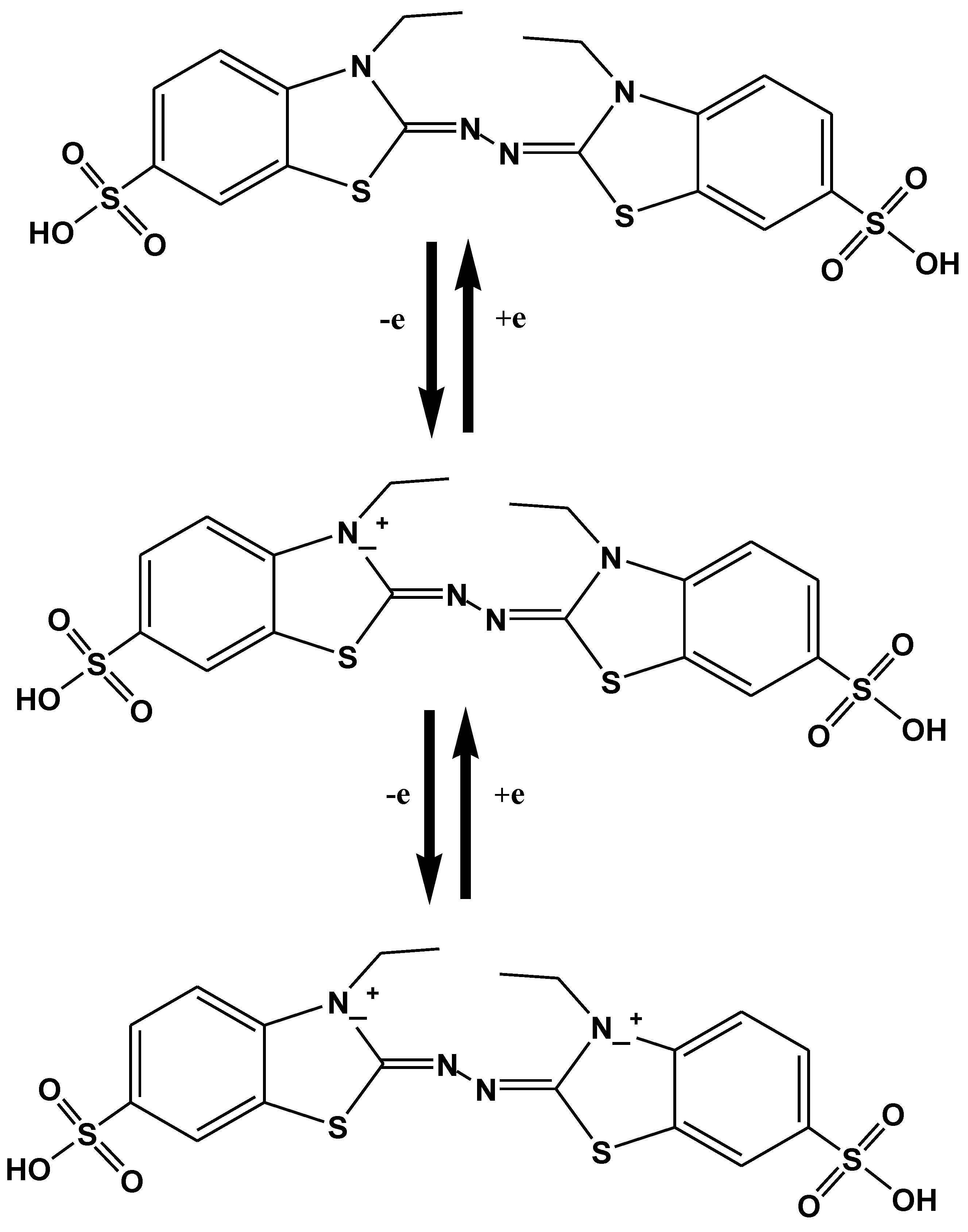
Numerous artificial mediators have been discovered oxidizing the non-phenolic structural moieties of lignin [111]. They remain the subject of wide range of study, from the very first described laccase-mediator, ABTS; to the synthetic mediators of -NOH- type (e.g., 1-hydroxybenzotriazole (HBT), N-hydroxyphtalimide (HPI), violuric acid (VLA), N-hydroxyacetanilide (HAA) and N-hydroxyacetanilide (NHA)) and the stable one 2,2,6,6-tetramethyl-1-piperidinyloxy free radical or TEMPO [111,112,113]. ABTS has been considered the best substrate-mediator laccase. It speeds up the rate of reaction by moving the electron towards electron accepting compounds from the donor substrate. Two stages are involved in the oxidation of ABTS. In the earlier stage, fast oxidation occurs and cation radical (ABTS+) is formed, after that di-cation (ABTS2+) formed by the slow oxidation of cation radical (Figure 7) [114].
A large body of literature has explained ABTS application of lignin degradation using laccase. The use of mediators, most probably ABTS is unique for the oxidation of lignin subunits. Many workers examined the Kraft lignin oxidation by Trametes versicolor (LTV) laccase and stated that ABTS coupled with laccase enhance the catalytic activity of laccase to generate lignin subunits having an average weight of 5300 g/mol [112]. The mechanism of ABTS oxidation indicates that ABTS2+ di-cation only act as an intermediate, for oxidation of non-phenolic structures. Conversely, ABTS+-cation radical accounts for phenolic structures [114]. In previous studies, researchers mostly concentrated on the oxidation mechanism of ethers, alcohols and lignin model compounds. Extensive research has described the effects of mediators and laccase enzyme on lignin model compounds to fully recognize the laccase reaction owing to the lignin structure complexity [112] (Figure 8).
4.1.3. Model Compounds of Lignin
The structural variability and complexity of lignin provoked the use of various lignin model compounds in its place to study the lignin depolymerization [115]. Such model compounds bear a resemblance to lignin polymer and investigation of their reactivity gives understanding about the reactivity of lignin polymer itself. Several factors lead to the use of lignin model compounds:
- to perceive the interaction between lignin and enzymes by using lignin model compounds in place of lignin due to their simple structure;
- many model compounds contain lignin-related linkages i.e., β-O-4, α-O-4, β-5, 4-O-5, etc. so their reactivity give the information relevant to lignin-enzyme interaction;
- the product and analysis of such model compounds are relatively easy as compared to lignin. Many publications give the idea about the interaction of lignin with laccase; though, the lignin degradation mechanism is much more difficult to understand [116].
4.1.4. Lignin Peroxidase (LiP)
Lignin peroxidases are heme containing proteins having an iron protoporphyrin prosthetic group, first isolated from Phanerochaete chrysosporium. These peroxidases catalyze the oxidation of a broad variety of aromatic compounds in the presence of H2O2 [117].
This enzyme had been completely characterized and its catalytic mechanism was studied previously in oxidizing substrate. Firstly, the enzyme is oxidized using hydrogen peroxide (H2O2) to LiPI (intermediate of LiP) and water. LiPI then converts to LiPII and substrate radical (VA+) by the oxidation of first molecule of veratryl-alcohol (VA). LiPII use the second veratryl alcohol (VA) by the reduction of the substrate and the enzyme recover in its original form as mentioned in Equation (1) [118].
Enzyme (LiP) + H202→ LiP I + H20
LiP I + VA → LiP II + VA+
LiP II + VA → Enzyme (LiP) + VA+
LiP I + VA → LiP II + VA+
LiP II + VA → Enzyme (LiP) + VA+
Since 1986, veratryl alcohol (VA) had been a redox mediator for LiP; it did not react with lignin in the absence of veratryl alcohol. So, lignin depolymerization via LiP was performed by adding veratryl alcohol [119].
4.1.5. Manganese Peroxidase (MnP)
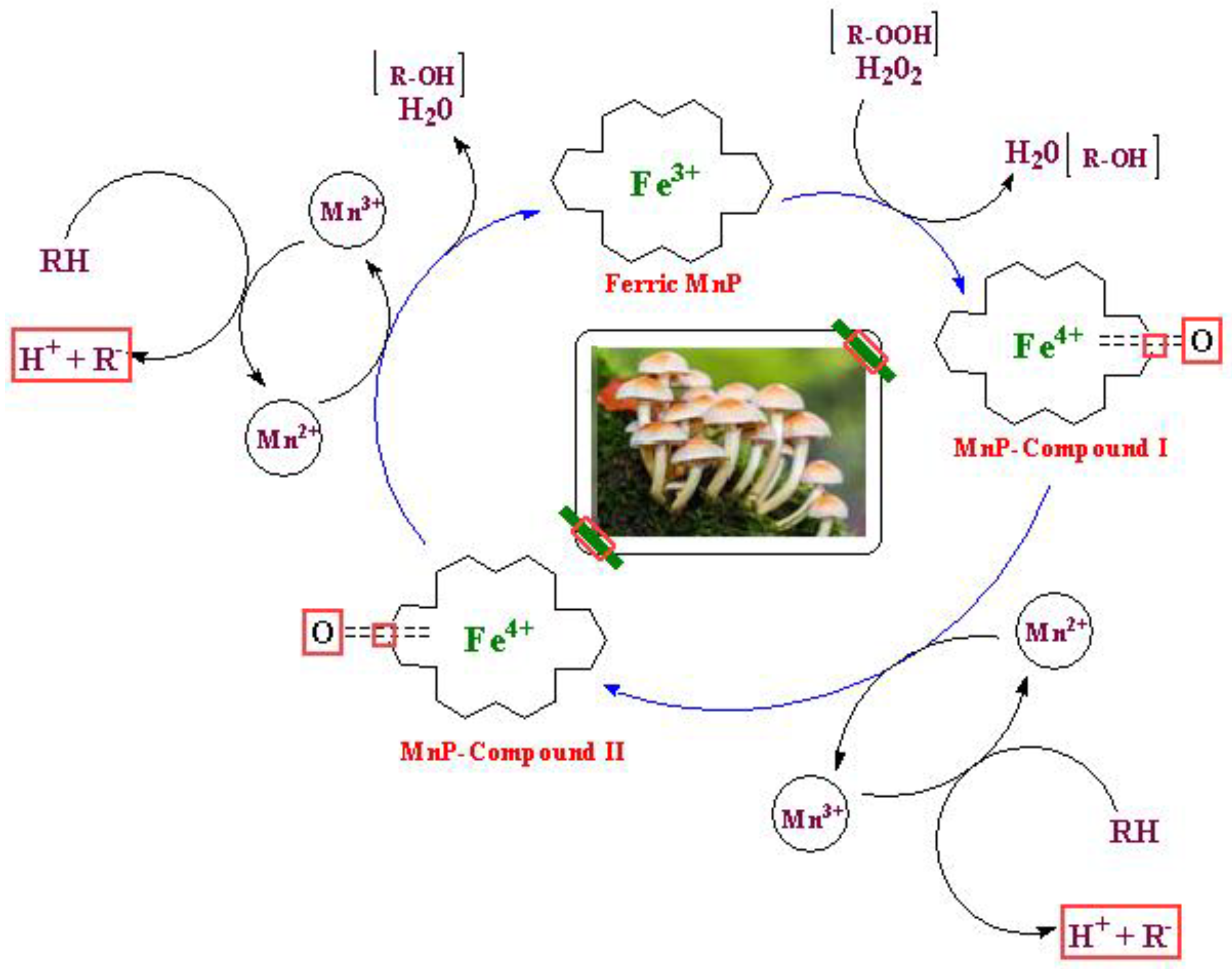
Manganese (Mn) is required for MnP synthesis. This enzyme has the pivotal role for earlier stages of degrading lignin polymer [120] and are produced by wide-ranging species of white rot basidiomycetes like Phanerochaete chryosporium [121]. For last 25 years, production of heme-peroxidases remained an interesting subject for researchers which include both manganese peroxidase (MnP) and lignin peroxidase (LiP) [122]. MnP, like LiP are heme containing proteins as well that use H2O2 as a co-substrate in the substrate oxidation. Like LiP, MnP also produces the intermediates (MnP-I and MnP-II) in its catalytic cycle [118].
The nature of substrate makes the main difference between MnP and LiP. Unlike LiP, primary substrate of MnP is Mn(II) instead of phenol and produces Mn(III) which is highly reactive and oxidizes a variety of phenolic compounds. Firstly, iron-peroxide complex is formed when native ferric MnP bound to H2O2. MnP-Compound-I along with a molecule of water produces by the transfer of electrons from MnP. Mn2+ oxidized to Mn3+ and transfer the electron to the porphyrin intermediate while MnP-Compound-I transformed to MnP-compound-II [123]. MnP-II reduces in a similar way and regenerate the native MnP along with a second water molecule. Mn3+ ion chelated with organic acids (malonate and lactate) makes possible Mn3+ release from the active site of MnP. This detachment increases the oxidation rate by stimulating the MnP activity. Chelates of Mn3+ ion cause the oxidation of many substrates or the removal of radicals (Figure 9) [124].
4.1.6. Versatile Peroxidases (VP)
A novel peroxidase from Pleurotus eryngii was reported and this peroxidase contains both main peroxidase properties (LiP and MnP) that can modify lignin molecule without the involvement of external mediator [125]. This enzyme, named as versatile peroxidase (VP), indicates that it has properties of both LiP and MnP and can oxidize various substrates including Mn2+, phenolic compounds and non-phenolic aromatic compounds e.g., veratryl alcohol [126]. Versatile peroxidase is isolated from white-rot fungi types like Bjerkandera spp. strain BOS55, Pleurotus ostreatus and Bjerkandera adusta [127]. Furthermore, VP are characterized by having extensive specificity for aromatic substrates, making them highly beneficial in certain applications including recalcitrant pollutants bioremediation as well as gaining major industrial interests these days [128].
In summary, heme containing peroxidases (LiP, MnP and VP) also have some drawbacks that limit their use as well. MnP, LiP and VP involve the use of H2O2 for their catalytic activity while laccases only require O2 which they absorb from the atmosphere directly. Peroxidases are extreme expensive and are not commercially available yet, in contrast, laccases are available at low prices. In comparison to peroxidases, laccases offer selection of mediator compounds for the process requirements. Consequently, laccase is a potential enzyme for degradation of lignin with promising applications that might improve efficiency and productivity with low investment cost [129]. Fungal enzymes biosynthesis for the depolymerization of lignin on industrial scale or chemical functionalization has been hindered by complications like culturing white rot fungi on an industrial scale as well as in lignin-degrading enzymes expression in other fungi [130,131].
5. Bonds Cleavage in Lignin
Variety of depolymerization protocols are employed to yield ‘green’ chemicals from lignin. The production of aromatic chemicals might be achieved through several processing routes using the lignin enriched fractions [132]. The regulated breaking of different linkages in lignin requires detailed information regarding the stability of the bonds under different conditions in addition to understand the lignin decomposition mechanism. In lignin, both ester and ether bonds are easily hydrolysable. Lignin can also be degraded by means of biological methods with micro-organisms, by chemical routes or via sun light (UV) [133].
Monomeric Lignin Molecules
Selective depolymerization involving C-O and C-C bond rupturing produce an excess of complex aromatics structures that are either difficult to generate via conventional petrochemical ways. These compounds are correlated to the fundamental building blocks of lignin and are highly desirable due to their production in a reasonable commercial amount. However, two barriers would have to be overcome. The first one is the advancement in technology for careful bond-scission to separate out the monomeric lignin structures, although this technology would be more difficult to develop than the other destructive processes that yield phenols or BTX. Secondly, applications and markets for lignin monomers are needed to be developed. For these reasons, this technology has long-term applications and currently their large-scale use is unknown [134].
6. Green Chemicals
By exploring the chemical worth of biomass, green chemical technologies developed to capture the resources and maximize the production of value added plus environmental friendly chemicals. In this integrating approach, high value chemicals co-produce which maximizes the use of all biomass components, waste streams and by-products virtually with keeping environmental footprint low [105]. Green chemicals obtained from the lignin are linked to the well-being of the environment with the potential production of renewable fuels, polymer building blocks and aromatic monomers such as phenol, vanillin, benzene, toluene, and xylene (BTX) [135].
6.1. Lignin for Production of Aromatic Chemicals
Lignin (the renewable raw material) is probably present in ample amounts for the synthesis of aromatic substances at industrial level. It seems easy to conclude that efficient and direct conversion of lignin into low molecular weight and distinct aromatic compounds is highly remarkable goal. But the synthesis of defined high-volume aromatic chemicals using diverse and physically intricate lignin is feasible and long-term opportunity, although it is a most challenging goal to achieve (Figure 10) [136].
Aromatics/Phenolics
Benzene, toluene and xylene (BTX) have large-scale of applications. Therefore, it has great potential in chemical industry. Lignin-based BTX is similar to BTX from petroleum and so can be used as a replacement for it. BTX represents 60% of all aromatics on market and 24% of the global petrochemical market [137].
Lignin can be depolymerized into various aromatic components. As these compounds are obtained from lignin, the first and the foremost duty is to eradicate the oxygen containing functional groups by decarboxylation, decarbonylation, dehydroxylation and demethoxylation [138]. Benzene is a resourceful petrochemical building block from which more than 250 products could be formed. Cyclohexane, ethyl benzene and cumene are the chief derivatives. The xylenes product well-known as mixed xylene contains four different isomers: ortho-xylene, para-xylene, meta-xylene and ethyl benzene. Toluene is gaining importance for the xylenes manufacturing through disproportionation of toluene and trans-alkylation with C-9 aromatics [139]. Aromatic complexes are found in several different configurations. However, most modern complexes of aromatics are considered to maximize the yield of para-xylene, benzene and sometimes ortho-xylene [140].
The main advantage of generating phenols from lignin is that nowadays phenol prices are quite high based on prices of oil. On the other hands, lignin as part of renewable source has relatively stable market value [137]. By focusing on phenol and its derivatives, the phenolic hydroxyl and the aromatic ring needs to be remain intact and thus less energy will be required to convert polyphenolic ligneous complex into useful compounds [141].
6.2. Lignin Valorization to Polymers
Advances in fractionation, catalyst development and purification technologies are necessary to obtain the required final depolymerized lignin by-product [142]. Shortly, lignin has the capability to substitute polymers such as polyacrylonitrile (PAN) to manufacture carbon fiber [143].
6.2.1. Carbon Fiber
Carbon fibers with properties like low density, high stiffness and extensive strength are highly valuable composite material [144]. They have wide range of applications that are increasing day by day. The prices of carbon fibers for automotive industry is relatively high these days. The precursor for carbon fiber is polyacrylonitrile (PAN) that makes 50% of all production costs. Lignin plays its role to lessen this production cost. Lignin, being a replacement for PAN makes the process much cost effective with potential usage involve manufacturing of sport goods and aircrafts, utilization in automobile industry as well as in civil engineering [137].
Kayacarbon developed the first lignin based carbon fibers, while Nippon Kayaku Co. in Tokyo, Japan made its commercial availability possible. Initially, lignin is melt-spun at high rates to generate economical lignin-derived carbon fiber that demands high purity lignin. The contaminations like polysaccharides, salts, water and further volatiles should be eliminated to obtain required results. Both Graf Tech International Holdings and Oak Ridge National Laboratory hold the record of generating elevated temperature thermal insulation prototypes by using lignin based carbon fibers [145].
6.2.2. Polymer Blends
Unmodified lignin does not have properties to be used as material. On the other hand, it can be blended with other synthetic or bio-based polymers. Lignin usually acts like UV degradation stabilizer or thermo-oxidation stabilizer. This functions lignin fulfils if it is blended with polyethylene, polystyrene, polypropylene or natural rubber [144].
6.2.3. Binders
Lignin do have major impact in the agrochemicals sector as well. Many applications including resins and foams sectors coupled with polymers and cement, which are tremendous dispersing agents and binders with dust controlling abilities are the results of lignosulfonates. Concrete industry is the main and the largest applications of lignosulfonates [146]. The strong dispersing agent properties of lignosulfonates allow less water consumption that provide the resulting concrete with better durability, elevated density and higher compressive strength etc. before utilizing as workable mixtures [147].
6.2.4. Polyurethane
Polyurethane, with an extensive variety of products in varied sectors, like paints, foam, adhesives, elastomer etc. is the most versatile polymers obtained by lignin valorization. Through its great insulation and mechanical properties, Rigid polyurethane (RPU having high mechanical and insulator capabilities) coupled with foams and elastomers is frequently used in freeze sectors, equipment manufacturing, automotive industry and construction in addition to nautical applications [148].
6.3. Certain New Products from Lignin Valorization
Several academic groups are working hard to attain a vast scope of new applications and useful products including fuel cells and high-performance materials, composites and batteries from lignin along with the classic aromatics and polymers [143,149,150]. Furthermore, by direct lignin fuel cells, lignin can also be used as fuel [151]. A novel N-doped fused carbon fibrous mat constructed via 9:1 combination of lignin:polyethylene oxide has been reported [152]. Since lignin possesses an aromatic character making it a remarkable preliminary material for graphite electrodes [153]. This can also prove to be an important and useful source for both fuel cells and lithium batteries as described presently [149].
7. Conclusions
More recent research work including the biological modifications of lignin indicates that lignin can be depolymerized into variety of useful chemicals of industrial importance. For the progress of an economical viable lignin valorization path to synthesize aromatic chemicals, advanced methods are required to assess the ideal conditions, appropriate hydrogen donors together with bio-refinery catalysts. There is also much need of the time to further develop this process for the commercial production of high purity lignin and lignin based byproducts.
In nature, lignocellulosic residues obtained via municipal solid wastes, agricultural source, grass, wood and forestry substances are available in bulk quantities and have an enormous bio-conversion potential. As a renewable resource, they are an important source of both biologically and chemically useful products. Lignin, when accumulated in sufficient amounts at places where agricultural residues reveal a discarding nuisance result in environmental decline coupled with valuable materials loss that can be helpful in paper and pulp industry as well as biomass fuel production and composting.
Varieties of innovative markets for lignocellulosic residues especially of lignin like Benzene, Toluene and Xylene (BTX) have been identified in recent times. Low cost bioremediation projects by utilizing fungi seem to be promising as they are the source of well-organized lignocellulose depolymerization enzyme machinery. However, additional consideration of the innumerable other enzymes coupled with organic acids for depolymerization reactions and its molecular features will be desired. The most remarkable task is to assimilate various enzymes roles and organic acids together with natural lignin degradation using a variety of microbes. Thus, lignin valorization by organic acids seems to be much more effective and safe to increase the product quantities and as well as to decrease costs compared to certain other costly manufacturing protocols.
Author Contributions
Conceptualization, This manuscript was concept of R.J. and A.S.; Writing-Original Draft Preparation was done by R.J.; Writing-Review & Editing was done by R.J. and M.F.; Visualization by A.S.; Supervision, N.S. and M.F.
Funding
The authors are thankful to worthy Vice Chancellor of University of the Punjab, Lahore, Pakistan for providing financial support for the accomplishment of this manuscript.
Acknowledgments
The author thanks to their potential institute (CAMB, PU, Lahore) and particularly to Higher Education Commission (HEC), Pakistan Grant No: 21-(1105/SRGP/R&D/HEC/2016) for providing the necessary facilities and funds to accomplish this work.
Conflicts of Interest
All authors declare no conflict of interest. The founding sponsors had no role in the writing of the manuscript and in the decision to publish it.
References
- Ferhan, M.; Tanguy, N.; Yan, N.; Sain, M. Comparison of Enzymatic, Alkaline, and UV/H2O2 Treatments for Extraction of Beetle-Infested Lodgepole Pine (BILP) and Aspen Bark Polyphenolic Extractives. ACS Sustain. Chem. Eng. 2013, 2, 165–172. [Google Scholar] [CrossRef]
- Savy, D.; Mazzei, P.; Roque, R.; Nuzzo, A.; Bowra, S.; Santos, R. Structural recognition of lignin isolated from bioenergy crops by subcritical water: Ethanol extraction. Fuel Process. Technol. 2015, 138, 637–644. [Google Scholar] [CrossRef]
- Long, H.; Li, X.; Wang, H.; Jia, J. Biomass resources and their bioenergy potential estimation: A review. Renew. Sustain. Energy Rev. 2013, 26, 344–352. [Google Scholar] [CrossRef]
- Perez, J.; Munoz-Dorado, J.; de la Rubia, T.D.; Martinez, J. Biodegradation and biological treatments of cellulose, hemicellulose and lignin: An overview. Int. Microbiol. 2002, 5, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.K.; Saini, R.; Tewari, L. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: Concepts and recent developments. 3 Biotech 2015, 5, 337–353. [Google Scholar] [CrossRef] [PubMed]
- Chundawat, S.P.; Beckham, G.T.; Himmel, M.E.; Dale, B.E. Deconstruction of lignocellulosic biomass to fuels and chemicals. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Laurichesse, S.; Averous, L. Chemical modification of lignins: Towards biobased polymers. Prog. Polym. Sci. 2014, 39, 1266–1290. [Google Scholar] [CrossRef]
- Abdelaziz, O.; Brink, D.P.; Prothmann, J.; Ravi, K.; Sun, M.; Garcia-Hidalgo, J.; Sandahl, M.; Hulteberg, C.P.; Turner, C.; Liden, G.; et al. Biological valorization of low molecular weight lignin. Biotechnol. Adv. 2016, 34, 1318–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mood, S.H.; Golfeshan, A.H.; Tabatabaei, M.; Jouzani, G.S.; Najafi, G.H.; Gholami, M.; Ardjmand, M. Lignocellulosic biomass to bioethanol, a comprehensive review with a focus on pretreatment. Renew. Sustain. Energy Rev. 2013, 27, 77–93. [Google Scholar] [CrossRef]
- Kumar, R.; Wyman, C.E. Effects of cellulase and xylanase enzymes on the deconstruction of solids from pretreatment of poplar by leading technologies. Biotechnol. Prog. 2009, 25, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, J.; Chang, X.; Chen, D.; Xue, Y.; Liu, P.; Lin, H.; Han, S. A review on the pretreatment of lignocellulose for high-value chemicals. Fuel Process. Technol. 2017, 160, 196–206. [Google Scholar] [CrossRef]
- Wan, C.; Li, Y. Fungal pretreatment of lignocellulosic biomass. Biotechnol. Adv. 2012, 30, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.P.; Hou, X.D.; Li, N.; Zong, M.H. Ionic liquids from renewable biomaterials: Synthesis, characterization and application in the pretreatment of biomass. Green Chem. 2012, 14, 304–307. [Google Scholar] [CrossRef]
- Travaini, R.; Otero, M.D.M.; Coca, M.; Da-Silva, R.; Bolado, S. Sugarcane bagasse ozonolysis pretreatment: Effect on enzymatic digestibility and inhibitory compound formation. Bioresour. Technol. 2013, 133, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Mirahmadi, K.; Mohseni Kabir, M.; Jeihanipour, A.; Karimi, K.; Taherzadeh, M. Alkaline pretreatment of spruce and birch to improve bioethanol and biogas production. BioResources 2010, 5, 928–938. [Google Scholar] [CrossRef]
- Mancini, G.; Papirio, S.; Lens, P.N.L.; Esposito, G. Solvent pretreatments of lignocellulosic materials to enhance biogas production: A review. Energy Fuels 2016, 30, 1892–1903. [Google Scholar] [CrossRef]
- Sathitsuksanoh, N.; Zhu, Z.G.; Zhang, Y.H.P. Cellulose solvent-based pretreatment for corn stover and Avicel: Concentrated phosphoric acid versus ionic liquid [BMIM]Cl. Cellulose 2012, 19, 1161–1172. [Google Scholar] [CrossRef]
- Mesa, L.; Gonzalez, E.; Cara, C.; Gonzalez, M.; Castro, E.; Mussatto, S.I. The effect of organosolv pretreatment variables on enzymatic hydrolysis of sugarcane bagasse. Chem. Eng. J. 2011, 168, 1157–1162. [Google Scholar] [CrossRef]
- Sharma, S.; Kumar, R.; Gaur, R.; Agrawal, R.; Gupta, R.P.; Tuli, D.K.; Das, B. Pilot scale study on steam explosion and mass balance for higher sugar recovery from rice straw. Bioresour. Technol. 2015, 175, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Perez, J.A.; Ballesteros, I.; Ballesteros, M.; Saez, F.; Negro, M.J.; Manzanares, P. Optimizing liquid hot water pretreatment conditions to enhance sugar recovery from wheat straw for fuel-ethanol production. Fuel 2008, 87, 3640–3647. [Google Scholar] [CrossRef]
- Lamsal, B.; Yoo, J.; Brijwani, K.; Alavi, S. Extrusion as a thermomechanical pre-treatment for lignocellulosic ethanol. Biomass Bioenergy 2010, 34, 1703–1710. [Google Scholar] [CrossRef]
- Balan, V.; Bals, B.; Chundawat, S.P.; Marshall, D.; Dale, B.E. Lignocellulosic biomass pretreatment using AFEX. In Biofuels: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2009; pp. 61–77. [Google Scholar]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, M.R.; Kim, S.; Lawrence, K.; Paton, R.S.; Chmely, S.C.; Nimlos, M.; Beckham, G.T. A mechanistic investigation of acid-catalyzed cleavage of aryl-ether linkages: Implications for lignin depolymerization in acidic environments. ACS Sustain. Chem. Eng. 2013, 2, 472–485. [Google Scholar] [CrossRef]
- Desnoyer, A.N.; Fartel, B.; MacLeod, K.C.; Patrick, B.O.; Smith, K.M. Ambient-Temperature Carbon–Oxygen Bond Cleavage of an α-Aryloxy Ketone with Cp2Ti (BTMSA) and Selective Protonolysis of the Resulting Ti–OR Bonds. Organometallics 2012, 31, 7625–7628. [Google Scholar] [CrossRef]
- Tobisu, M.; Chatani, N. Catalytic Hydrogenolysis of C-O Bonds in Aryl Ethers. ChemCatChem 2011, 3, 1410–1411. [Google Scholar] [CrossRef]
- Lu, F.C.; Ralph, J. Solution-state NMR of lignocellulosic biomass. J. Biobased Mater. Bioenergy 2011, 5, 169–180. [Google Scholar] [CrossRef]
- Mansfield, S.D.; Kim, H.; Lu, F.; Ralph, J. Whole plant cell wall characterization using solution-state 2D NMR. Nat. Protoc. 2012, 7, 1579–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, Y.Q.; Cao, S.L.; Ragauskas, A.J. Application of quantitative 31P NMR in biomass lignin and biofuel precursors characterization. Energy Environ. Sci. 2011, 4, 3154–3166. [Google Scholar] [CrossRef]
- Donohoe, B.S.; Decker, S.R.; Tucker, M.P.; Himmel, M.E.; Vinzant, T.B. Visualizing lignin coalescence and migration through maize cell walls following thermochemical pretreatment. Biotechnol. Bioeng. 2008, 101, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Carpita, N.C. Progress in the biological synthesis of the plant cell wall: New ideas for improving biomass for bioenergy. Curr. Opin. Biotechnol. 2012, 23, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Mielenz, J.R.; Xiao, X.; Ge, Y.; Hamilton, C.Y.; Rodriguez, M.; Chen, F.; Foston, M.; Ragauskas, A.; Bouton, J.; et al. Genetic manipulation of lignin reduces recalcitrance and improves ethanol production from switchgrass. Proc. Natl. Acad. Sci. USA 2011, 108, 3803–3808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziebell, A.; Gracom, K.; Katahira, R.; Chen, F.; Pu, Y.Q.; Ragauskas, A.; Dixon, R.A.; Davis, M. Increase in 4-coumaryl alcohol units during lignification in alfalfa (Medicago sativa) alters the extractability and molecular weight of lignin. J. Biol. Chem. 2010, 285, 38961–38968. [Google Scholar] [CrossRef] [PubMed]
- Fort, D.A.; Remsing, R.C.; Swatloski, R.P.; Moyna, P.; Moyna, G.; Rogers, R.D. Can ionic liquids dissolve wood? Processing and analysis of lignocellulosic materials with 1-n-butyl-3-methylimidazolium chloride. Green Chem. 2007, 9, 63–69. [Google Scholar] [CrossRef]
- Pu, Y.Q.; Jiang, N.; Ragauskas, A.J. Ionic liquid as a green solvent for lignin. J. Wood Chem. Technol. 2007, 27, 23–33. [Google Scholar] [CrossRef]
- Li, C.L.; Knierim, B.; Manisseri, C.; Arora, R.; Scheller, H.V.; Auer, M.; Vogel, K.P.; Simmons, B.A.; Singh, S. Comparison of dilute acid and ionic liquid pretreatment of switchgrass: Biomass recalcitrance, delignification and enzymatic saccharification. Bioresour. Technol. 2010, 101, 4900–4906. [Google Scholar] [CrossRef] [PubMed]
- Bozell, J.J.; Black, S.K.; Myers, M.; Cahill, D.; Miller, W.P.; Park, S. Solvent fractionation of renewable woody feedstocks: Organosolv generation of biorefinery process streams for the production of bio-based chemicals. Biomass Bioenergy 2011, 35, 4197–4208. [Google Scholar] [CrossRef]
- Bozell, J.J.; O’Lenick, C.J.; Warwick, S. Biomass fractionation for the biorefinery: Heteronuclear multiple quantum coherence—Nuclear magnetic resonance investigation of lignin isolated from solvent fractionation of switchgrass. J. Agric. Food Chem. 2011, 59, 9232–9242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.P.; Ding, S.Y.; Mielenz, J.R.; Cui, J.B.; Elander, R.T.; Laser, M.; Himmel, M.E.; McMillan, J.R.; Lynd, L.R. Fractionating recalcitrant lignocellulose at modest reaction conditions. Biotechnol. Bioeng. 2007, 97, 214–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hergert, H.L. Developments in Organosolv Pulping. An Overview; Wiley: New York, NY, USA, 1998; pp. 5–67. [Google Scholar]
- Sahin, H.T. Base-catalyzed organosolv pulping of jute. J. Chem. Technol. Biotechnol. 2003, 78, 1267–1273. [Google Scholar] [CrossRef]
- Sarkanen, K.V. Chemistry of Solvent Pulping. In Proceedings of the Tappi Pulping Conference, Toronto, ON, Canada, 14–17 October 1990; pp. 651–654. [Google Scholar]
- Nimz, H.H.; Casten, R. Chemical processing of lignocellulosics. Eur. J. Wood Wood Prod. 1986, 44, 207–212. [Google Scholar] [CrossRef]
- Young, R.A.; Davis, J.L.; Wiesmann, E.B. Organic Acid Pulping of Wood-Part II. Acetic Acid Pulping of Aspen. Holzforschung 1986, 40, 99–108. [Google Scholar] [CrossRef]
- Davis, J.L.; Young, R.A.; Deodhar, S.S. Organic acid pulping of wood, 3: Acetic acid pulping of spruce. J. Jpn. Wood Res. Soc. 1986. (In Japanese) [Google Scholar]
- Dapia, S.; Santos, V.; Parajo, J.C. Study of formic acid as an agent for biomass fractionation. Biomass Bioenergy 2002, 22, 213–221. [Google Scholar] [CrossRef]
- Jahan, M.S.; Chowdhury, D.N.; Islam, M.K. Atmospheric formic acid pulping and TCF bleaching of dhaincha (Sesbania aculeata), kash (Saccharum spontaneum) and banana stem (Musa Cavendish). Ind. Crops Prod. 2007, 26, 324–331. [Google Scholar] [CrossRef]
- Lam, H.Q.; Le Bigot, Y.; Delmas, M. Formic acid pulping of rice straw. Ind. Crops Prod. 2001, 14, 65–71. [Google Scholar] [CrossRef]
- Xu, J.; Thomsen, M.H.; Thomsen, A.B. Pretreatment on corn stover with low concentration of formic acid. J. Microbiol. Biotechnol. 2009, 19, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Ligero, P.; Villauerde, J.J.; de Vega, A.; Bao, M. Delignification of Eucalyptus globulus saplings in two organosolv systems (formic and acetic acid) preliminary analysis of dissolved lignins. Ind. Crops Prod. 2008, 27, 110–117. [Google Scholar] [CrossRef]
- Zhang, M.; Qi, W.; Liu, R.; Su, R.; Wu, S.; He, Z. Fractionating lignocellulose by formic acid: Characterization of major components. Biomass Bioenergy 2010, 34, 525–532. [Google Scholar] [CrossRef]
- Magnuson, J.K.; Lasure, L.L. Organic acid production by filamentous fungi. In Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine; Springer: New York, NY, USA, 2004; pp. 307–340. [Google Scholar]
- Plassard, C.; Fransson, P. Regulation of low-molecular weight organic acid production in fungi. Fungal Biol. Rev. 2009, 23, 30–39. [Google Scholar] [CrossRef]
- Dutton, M.V.; Evans, C.S. Oxalate production by fungi: Its role in pathogenicity and ecology in the soil environment. Can. J. Microbiol. 1996, 42, 881–895. [Google Scholar] [CrossRef]
- Jones, D.L. Organic acids in the rhizosphere—A critical review. Plant Soil 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Green, F.; Highley, T.L. Mechanism of brown-rot decay: Paradigm or paradox. Int. Biodeterior. Biodegrad. 1997, 39, 113–124. [Google Scholar] [CrossRef]
- Shimada, M.; Akamtsu, Y.; Tokimatsu, T.; Mii, K.; Hattori, T. Possible biochemical roles of oxalic acid as a low molecular weight compound involved in brown-rot and white-rot wood decays. J. Biotechnol. 1997, 53, 103–113. [Google Scholar] [CrossRef]
- Tanaka, N.; Akamatsu, Y.; Hattori, T.; Shimada, M. Effect of oxalic acid on the oxidative breakdown of cellulose by the Fenton reaction. Wood Res. Bull. Wood Res. Inst. Kyoto Univ. 1994, 81, 8–10. [Google Scholar]
- Ahonen-Jonnarth, U.; Van Hees, P.A.; Lundstrom, U.S.; Finlay, R.D. Organic acids produced by mycorrhizal Pinus sylvestris exposed to elevated aluminium and heavy metal concentrations. New Phytol. 2000, 146, 557–567. [Google Scholar] [CrossRef]
- Espejo, E.; Agosin, E. Production and degradation of oxalic acid by brown rot fungi. Appl. Environ. Microbiol. 1991, 57, 1980–1986. [Google Scholar] [PubMed]
- Hastrup, A.C.S.; Green, F.; Lebow, P.K.; Jensen, B. Enzymatic oxalic acid regulation correlated with wood degradation in four brown-rot fungi. Int. Biodeterior. Biodegrad. 2012, 75, 109–114. [Google Scholar] [CrossRef]
- Makela, M.; Galkin, S.; Hatakka, A.; Lundell, T. Production of organic acids and oxalate decarboxylase in lignin-degrading white rot fungi. Enzym. Microb. Technol. 2002, 30, 542–549. [Google Scholar] [CrossRef]
- Vanhees, P.; Jones, D.; Jentschke, G.; Godbold, D. Organic acid concentrations in soil solution: Effects of young coniferous trees and ectomycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 771–776. [Google Scholar] [CrossRef]
- Sauer, M.; Porro, D.; Mattanovich, D.; Branduardi, P. Microbial production of organic acids: Expanding the markets. Trends Biotechnol. 2008, 26, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Industrial Production of Organic Acids by Fungi: State of the Art and Opportunities. Available online: https://www.taylorfrancis.com/books/e/9781466585782/chapters/10.1201%2Fb15250-5 (accessed on 13 August 2018).
- Liaud, N.; Giniés, C.; Navarro, D.; Fabre, N.; Crapart, S.; Herpoël-Gimbert, I.; Sigoillot, J.C. Exploring fungal biodiversity: Organic acid production by 66 strains of filamentous fungi. Fungal Biol. Biotechnol. 2014, 1, 1. [Google Scholar] [CrossRef]
- Chesson, A. Mechanistic models of forage cell wall degradation. In Forage Cell Wall Structure and Digestibility, (Forage Cell Walls); American Society of Agronomy, Inc.: Madison, MI, USA, 1993; pp. 347–376. [Google Scholar]
- Kirk, T.K.; Farrell, R.L. Enzymatic ‘combustion’: The microbial degradation of lignin. Annu. Rev. Microbiol. 1987, 41, 465–501. [Google Scholar] [CrossRef] [PubMed]
- MaCarthy, A.J. Lignocellulose-degrading actinomycetes. FEMS Microbiol. Lett. 1987, 46, 145–163. [Google Scholar] [CrossRef]
- Hackett, W.F.; Connors, W.J.; Kirk, T.K.; Zeikus, J.G. Microbial decomposition of synthetic 14C-labeled lignins in nature: Lignin biodegradation in a variety of natural materials. Appl. Environ. Microbiol. 1977, 33, 43–51. [Google Scholar] [PubMed]
- Zeikus, J.G.; Wellstein, A.L.; Kirk, T.K. Molecular basis for the biodegradative recalcitrance of lignin in anaerobic environments. FEMS Microbiol. Lett. 1982, 15, 193–197. [Google Scholar] [CrossRef]
- Gaillard, B.D.; Richards, G.N. Presence of soluble lignin-carbohydrate complexes in the bovine rumen. Carbohydr. Res. 1975, 42, 135–145. [Google Scholar] [CrossRef]
- Chen, W.; Ohmiya, K.; Shimizu, S.; Kawakami, H. Anaerobic degradation of dehydrodiisoeugenol by rumen bacteria. J. Ferment. Technol. 1987, 65, 221–224. [Google Scholar] [CrossRef]
- Chen, W.; Ohmiya, K.; Shimizu, S.; Kawakami, H. Degradation of dehydrodivanillin by anaerobic bacteria from cow rumen fluid. Appl. Environ. Microbiol. 1985, 49, 211–216. [Google Scholar] [PubMed]
- Chen, W.; Supanwong, K.; Ohmiya, K.; Shimizu, S.; Kawakami, H. Anaerobic degradation of veratrylglycerol-beta-guaiacyl ether and guaiacoxyacetic acid by mixed rumen bacteria. Appl. Environ. Microbiol. 1985, 50, 1451–1456. [Google Scholar] [PubMed]
- Martin, M.M. Invertebrate-Microbial Interactions: Ingested Fungal Enzymes in Arthropod Biology; Cornell University Press: Ithaca, NY, USA, 1987. [Google Scholar]
- Lee, K.E.; Wood, T.G. Termites and Soils. 2001; Academic Press: London, UK, 1971. [Google Scholar]
- Martin, M.M. Cellulose digestion in insects. Comp. Biochem. Physiol. Part A Physiol. 1983, 75, 313–324. [Google Scholar] [CrossRef]
- Esenther, G.R.; Kirk, T.K. Catabolism of aspen sapwood in Reticulitermes flavipes (Isoptera: Rhinotermitidae). Annu. Entomol. Soc. Am. 1974, 67, 989–991. [Google Scholar] [CrossRef]
- Wood, T.G. Food and feeding habits of termites. In Production Ecology of Ants and Termites; Cambridge University Press: Cambridge, UK, 1978; pp. 55–80. [Google Scholar]
- Adler, E. Lignin chemistry—Past, present and future. Wood Sci. Technol. 1977, 11, 169–218. [Google Scholar] [CrossRef]
- Chakar, F.S.; Ragauskas, A.J. Review of current and future softwood Kraft lignin process chemistry. Ind. Crop. Prod. 2004, 20, 131–141. [Google Scholar] [CrossRef]
- Capanema, E.A.; Balakshin, M.Y.; Kadla, J.F. Quantitative characterization of a hardwood milled wood lignin by nuclear magnetic resonance spectroscopy. J. Agric. Food Chem. 2005, 53, 9639–9649. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, T. Lignin biochemistry: Biosynthesis and biodegradation. Wood Sci. Technol. 1990, 24, 23–63. [Google Scholar] [CrossRef]
- Lai, Y.Z.; Sarkanen, K.V. Isolation and Structural Studies. 1971, pp. 165–240. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/pol.1972.110100315 (accessed on 14 August 2018).
- Freudenberg, K.; Kraft, R.; Heimberger, W. Uber den Sinapinalkohol, den Coniferylalkohol und ihre Dehydrierungspolymerisate. Eur. J. Inorg. Chem. 1951, 84, 472–476. [Google Scholar] [CrossRef]
- Freudenberg, K. Lignin: Its constitution and formation from p-hydroxycinnamyl alcohols. Science 1965, 148, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, K.; Neish, A.C. Constitution and Biosynthesis of Lignin; Springer: Berlin, Germany, 1968; p. 129. [Google Scholar]
- Sarkanen, K.V. Precursors and their polymerization. In Lignins: Occurrence, Formation, Structure, and Reactions; Sarkanen, K.V., Ludwing, C.H., Eds.; Wiley: New York, NY, USA, 1971; pp. 95–163. [Google Scholar]
- Wood, T.G.; Thomas, R.J. The mutualistic association between Macrotermitinae and Termitomyces. In Insect-Fungus Interact; Academic Press: New York, NY, USA, 1989; pp. 69–92. [Google Scholar]
- Rohrmann, G.F.; Rossman, A.Y. Nutrient strategies of Macrotermes ukuzii (Isoptera: Termitidae). Pedobiologia 1980, 20, 61–73. [Google Scholar]
- Veivers, P.C.; Muhlemann, R.; Slaytor, M.; Leuthold, R.H.; Bignell, D.E. Digestion, diet and polyethism in two fungus-growing termites: Macrotermes subhyalinus Rambur and Macrotermes michaelseni Sjostedt. J. Insect Physiol. 1991, 37, 675–682. [Google Scholar] [CrossRef]
- Harwood, C.S.; Parales, R.E. The β-ketoadipate pathway and the biology of self-identity. Annu. Rev. Microbiol. 1996, 50, 553–590. [Google Scholar] [CrossRef] [PubMed]
- Hatakka, A. Lignin-modifying enzymes from selected white-rot fungi: Production and role from in lignin degradation. FEMS Microbiol. Rev. 1994, 13, 125–135. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, T.K. Lignin-degrading enzyme from the Hymenomycete Phanerochaete chrysosporium Burds. Science 1983, 221, 661–662. [Google Scholar] [CrossRef] [PubMed]
- Viswanath, B.; Rajesh, B.; Janardhan, A.; Kumar, A.P.; Narasimha, G. Fungal laccases and their applications in bioremediation. Enzym. Res. 2014, 2014, 163242. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.S.; Nityanand, C. Microbial laccases and their applications: A review. Asian J. Biotechnol. 2011, 3, 98–124. [Google Scholar] [CrossRef]
- Madhavi, V.; Lele, S.S. Laccase: Properties and applications. BioResources 2009, 4, 1694–1717. [Google Scholar]
- Saparrat, M.C.; Guillen, F.; Arambarri, A.M.; Martinez, A.T.; Martinez, M.J. Induction, isolation, and characterization of two laccases from the white rot basidiomycete Coriolopsis rigida. Appl. Environ. Microbiol. 2002, 68, 1534–1540. [Google Scholar] [CrossRef] [PubMed]
- Duran, N.; Rosa, M.A.; D’Annibale, A.; Gianfreda, L. Applications of laccases and tyrosinases (phenoloxidases) immobilized on different supports: A review. Enzym. Microbial. Technol. 2002, 31, 907–931. [Google Scholar] [CrossRef]
- Kunamneni, A.; Plou, F.J.; Ballesteros, A.; Alcalde, M. Laccases and their applications: A patent review. Recent Pat. Biotechnol. 2008, 2, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Gianfreda, L.; Xu, F.; Bollag, J.M. Laccases: A useful group of oxidoreductive enzymes. Bioremed. J. 1999, 3, 1–26. [Google Scholar] [CrossRef]
- Endo, K.; Hosono, K.; Beppu, T.; Ueda, K. A novel extracytoplasmic phenol oxidase of Streptomyces: Its possible involvement in the onset of morphogenesis. Microbiology 2002, 148, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Givaudan, A.; Effosse, A.; Faure, D.; Potier, P.; Bouillant, M.L.; Bally, R. Polyphenol oxidase in Azospirillum lipoferum isolated from rice rhizosphere: Evidence for laccase activity in non-motile strains of Azospirillum lipoferum. FEMS Microbiol. Lett. 1993, 108, 205–210. [Google Scholar] [CrossRef]
- Hullo, M.F.; Moszer, I.; Danchin, A.; Martin-Verstraete, I. Cot A of Bacillus subtilis is a copper-dependent laccase. J. Bacteriol. 2001, 183, 5426–5430. [Google Scholar] [CrossRef] [PubMed]
- Goodell, B.; Yamamoto, K.; Jellison, J.; Nakamura, M.; Fujii, T.; Takabe, K.; Hayashi, N. Laccase immunolabelling and microanalytical analysis of wood degraded by Lentinus edodes. Holzforschung 1998, 52, 345–350. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G.; Reid, I.D.; Lanthier, P.; Yaguchi, M. Lignin oxidation by laccase isozymes from Trametes versicolor and role of the mediator 2,2′-azinobis (3-ethylbenzthiazoline-6-sulfonate) in kraft lignin depolymerization. Appl. Environ. Microbiol. 1995, 61, 1876–1880. [Google Scholar] [PubMed]
- Kawai, S.; Umezawa, T.; Higuchi, T. Degradation mechanisms of phenolic β-1 lignin substructure model compounds by laccase of Coriolus versicolor. Arch. Biochem. Biophys. 1988, 262, 99–110. [Google Scholar] [CrossRef]
- Elegir, G.; Daina, S.; Zoia, L.; Bestetti, G.; Orlandi, M. Laccase mediator system: Oxidation of recalcitrant lignin model structures present in residual kraft lignin. Enzym. Microbial. Technol. 2005, 37, 340–346. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Shleev, S.V.; Yaropolov, Y.I. Laccase-mediator systems and their applications: A review. Appl. Biochem. Microbiol. 2007, 43, 523–535. [Google Scholar] [CrossRef]
- Rivera-Hoyos, C.M.; Morales-Álvarez, E.D.; Poutou-Piñales, R.A.; Pedroza-Rodríguez, A.M.; RodrÍguez-Vázquez, R.; Delgado-Boada, J.M. Fungal laccases. Fungal Biol. Rev. 2013, 27, 67–82. [Google Scholar] [CrossRef]
- Wong, D.W. Structure and action mechanism of ligninolytic enzymes. Appl. Biochem. Biotechnol. 2009, 157, 174–209. [Google Scholar] [CrossRef] [PubMed]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates. An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Jia, S.; Cox, B.J.; Guo, X.; Zhang, Z.C.; Ekerdt, J.G. Hydrolytic cleavage of β-O-4 ether bonds of lignin model compounds in an ionic liquid with metal chlorides. Ind. Eng. Chem. Res. 2010, 50, 849–855. [Google Scholar] [CrossRef]
- Zakzeski, J.; Bruijnincx, P.C.; Jongerius, A.L.; Weckhuysen, B.M. The catalytic valorization of lignin for the production of renewable chemicals. Chem. Rev. 2010, 110, 3552–3599. [Google Scholar] [CrossRef] [PubMed]
- Van Bloois, E.; Pazmino, D.E.T.; Winter, R.T.; Fraaije, M.W. A robust and extracellular heme-containing peroxidase from Thermobifida fusca as prototype of a bacterial peroxidase superfamily. Appl. Microbiol. Biotechnol. 2010, 86, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Dunford, H.B.; Stillman, J.S. On the function and mechanism of action of peroxidases. Coord. Chem. Rev. 1976, 19, 187–251. [Google Scholar] [CrossRef]
- Hammel, K.E.; Moen, M.A. Depolymerization of a synthetic lignin in vitro by lignin peroxidase. Enzym. Microb. Technol. 1991, 13, 15–18. [Google Scholar] [CrossRef]
- Perez, J.; Jeffries, T.W. Mineralization of 14C-ring-labeled synthetic lignin correlates with the production of lignin peroxidase, not of manganese peroxidase or laccase. Appl. Environ. Microbiol. 1990, 56, 1806–1812. [Google Scholar] [PubMed]
- Morgenstern, I.; Klopman, S.; Hibbett, D.S. Molecular evolution and diversity of lignin degrading heme peroxidases in the Agaricomycetes. J. Mol. Evol. 2008, 66, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Renganathan, V.; Gold, M.H. Spectral characterization of the oxidized states of lignin peroxidase, an extracellular heme enzyme from the white rot basidiomycete Phanerochaete chrysosporium. Biochemistry 1986, 25, 1626–1631. [Google Scholar] [CrossRef]
- Glenn, J.K.; Gold, M.H. Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-degrading basidiomycete, Phanerochaete chrysosporium. Arch. Biochem. Biophys. 1985, 242, 329–341. [Google Scholar] [CrossRef]
- Hofrichter, M. Lignin conversion by manganese peroxidase (MnP). Enzym. Microb. Technol. 2002, 30, 454–466. [Google Scholar] [CrossRef]
- Moreira, P.R.; Almeida-Vara, E.; Malcata, F.X.; Duarte, J.C. Lignin transformation by a versatile peroxidase from a novel Bjerkandera sp. strain. Int. Biodeterior. Biodegrad. 2007, 59, 234–238. [Google Scholar] [CrossRef]
- Camarero, S.; Sarkar, S.; Ruiz-Duenas, F.J.; Martinez, M.J.; Martinez, A.T. Description of a versatile peroxidase involved in the natural degradation of lignin that has both manganese peroxidase and lignin peroxidase substrate interaction sites. J. Biol. Chem. 1999, 274, 10324–10330. [Google Scholar] [CrossRef] [PubMed]
- Heinfling, A.; Ruiz-Duefias, F.J.; Martinez, M.J.; Bergbauer, M.; Szewzyk, U.; Martinez, A.T. A study on reducing substrates of manganese-oxidizing peroxidases from Pleurotus erygnii and Bjerkandera adusta. FEBS Lett. 1998, 428, 141–146. [Google Scholar] [CrossRef]
- Palma, C.; Lloret, L.; Sepúlveda, L.; Contreras, E. Production of versatile peroxidase from Pleurotus eryngii by solid-state fermentation using agricultural residues and evaluation of its catalytic properties. Prep. Biochem. Biotechnol. 2016, 46, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.; Hadar, Y.; Yarden, O. Transcript and activity levels of different Pleurotus ostreatus peroxidases are differentially affected by Mn2+. Environ. Microbiol. 2001, 3, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-degrading enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Zhao, X.; Wang, A.; Huber, G.W.; Zhang, T. Catalytic transformation of lignin for the production of chemicals and fuels. Chem. Rev. 2015, 115, 11559–11624. [Google Scholar] [CrossRef] [PubMed]
- Kamm, B.; Kamm, M. Principles of biorefineries. Appl. Microbiol. Biotechnol. 2004, 64, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Gosselink, R.J.A. Lignin as a Renewable Aromatic Resource for the Chemical Industry. 2011. Available online: https://www.wur.nl/upload_mm/d/7/f/35cbaa28-e1d6-4d6a-88f1-bfa1265b9af0_Mini-symp%206-12-2011%20Gosselink_Lignin%20as%20a%20renewable%20aromatic%20resource%20for%20the%20chem%20ind.pdfc (accessed on 13 August 2018).
- Gary, J.H.; Handwerk, G.E.; Kaiser, M.J. Petroleum Refining: Technology and Economics; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Sannigrahi, P.; Ragauskas, A.J.; Tuskan, G.A. Poplar as a feedstock for biofuels: A review of compositional characteristics. Biofuels Bioprod. Biorefin. 2010, 4, 209–226. [Google Scholar] [CrossRef]
- Holladay, J.E.; White, J.F.; Bozell, J.J.; Johnson, D. Top Value Added Chemicals from Biomass-Volume II, Results of Screening for Potential Candidates from Biorefinery Lignin (No. PNNL-16983); Pacific Northwest National Lab. (PNNL): Richland, WA, USA; National Renewable Energy Laboratory (NREL): Golden, CO, USA, 2007. [CrossRef]
- Smolarski, N. High-value opportunities for lignin: Unlocking its potential. Frost Sullivan 2012, 1, 15. [Google Scholar]
- Haveren, J.V.; Scott, E.L.; Sanders, J. Bulk chemicals from biomass. Biofuels Bioprod. Biorefin. 2008, 2, 41–57. [Google Scholar] [CrossRef]
- Allen, R.H.; Yats, L.D. Kinetics of three-compound equilibrations. V. Concurrent alkylation and isomerization. J. Am. Chem. Soc. 1961, 83, 2799–2805. [Google Scholar] [CrossRef]
- Gallezot, P. Catalytic routes from renewables to fine chemicals. Catal. Today 2007, 121, 76–91. [Google Scholar] [CrossRef]
- Song, Q.; Wang, F.; Cai, J.; Wang, Y.; Zhang, J.; Yu, W.; Xu, J. Lignin depolymerization (LDP) in alcohol over nickel-based catalysts via a fragmentation–hydrogenolysis process. Energy Environ. Sci. 2013, 6, 994–1007. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Perkins, J.H.; Vautard, F.; Meyer, H.M.; Messman, J.M.; Tolnai, B.; Naskar, A.K. Methanol fractionation of softwood kraft lignin: Impact on the lignin properties. ChemSusChem. 2014, 7, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Kadla, J.F.; Kubo, S.; Venditti, R.A.; Gilbert, R.D.; Compere, A.L.; Griffith, W. Lignin-based carbon fibers for composite fiber applications. Carbon 2002, 40, 2913–2920. [Google Scholar] [CrossRef]
- Lawoko, A.; Duval, D. A review on lignin-based polymeric, micro- and nanostructured materials. React. Funct. Polym. 2014, 85, 78–96. [Google Scholar] [CrossRef]
- Eberle, C.; Webb, D.C.; Albers, T.; Chen, C. Commercialization of New Carbon Fiber Materials Based on Sustainable Resources for Energy Applications; Oak Ridge National Laboratory (ORNL): Oak Ridge, TN, USA, 2013. [CrossRef]
- Stern, T.; Schwarzbauer, P. Wood-based lignosulfonate versus synthetic polycarboxylate in concrete admixture systems: The perspective of a traditional pulping by-product competing with an oil-based substitute in a business-to-business market in Central. Eur. For. Prod. J. 2008, 58, 81. [Google Scholar]
- Mailvaganam, N.P.; Rixom, M.R. Chemical Admixtures for Concrete; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Amaral, J.S.; Sepulveda, M.; Cateto, C.A.; Fernandes, I.P.; Rodrigues, A.E.; Belgacem, M.N.; Barreiro, M.F. Fungal degradation of lignin-based rigid polyurethane foams. Polym. Degrad. Stab. 2012, 97, 2069–2076. [Google Scholar] [CrossRef] [Green Version]
- Gnedenkov, S.V.; Opra, D.P.; Sinebryukhov, S.L.; Tsvetnikov, A.K.; Ustinov, A.Y.; Sergienko, V.I. Hydrolysis lignin-based organic electrode material for primary lithium batteries. J. Solid State Electrochem. 2013, 17, 2611–2621. [Google Scholar] [CrossRef]
- Zhang, R.; Xiao, X.; Tai, Q.; Huang, H.; Yang, J.; Hu, Y. Preparation of lignin-silica hybrids and its application in intumescent flame-retardant poly (lactic acid) system. High Perform. Polym. 2012, 24, 738–746. [Google Scholar] [CrossRef]
- Lima, R.B.; Raza, R.; Qin, H.; Li, J.; Lindstrom, M.E.; Zhu, B. Direct lignin fuel cell for power generation. RSC Adv. 2013, 3, 5083–5089. [Google Scholar] [CrossRef]
- Wang, S.X.; Yang, L.; Stubbs, L.P.; Li, X.; He, C. Lignin-derived fused electrospun carbon fibrous mats as high performance anode materials for lithium ion batteries. ACS Appl. Mater. Interfaces 2013, 5, 12275–12282. [Google Scholar] [CrossRef] [PubMed]
- Tenhaeff, W.E.; Rios, O.; More, K.; McGuire, M.A. Highly Robust Lithium Ion Battery Anodes from Lignin: An Abundant, Renewable, and Low Cost Material. Adv. Funct. Mater. 2014, 24, 86–94. [Google Scholar] [CrossRef]
Figure 1.
Lignin polymers can be terminated by a p-hydroxyl group or connected to additional lignin species, referred to as “phenolic” (X = OH) and “non-phenolic” (X = H or O-lignin) groups, respectively.
Figure 1.
Lignin polymers can be terminated by a p-hydroxyl group or connected to additional lignin species, referred to as “phenolic” (X = OH) and “non-phenolic” (X = H or O-lignin) groups, respectively.

Figure 2.
Ideal lignin molecule showing different linkages.

Figure 3.
Schematic representation of the lignin degradation steps and enzymes involved.

Figure 4.
Oxygen (O2) reduction into water (H2O) by laccase.

Figure 5.
Schematic diagram of laccase active site; containing four copper which belong to type-1, type-2 and type-3 binuclear copper site based on their electron paramagnetic resonance (EPR).
Figure 5.
Schematic diagram of laccase active site; containing four copper which belong to type-1, type-2 and type-3 binuclear copper site based on their electron paramagnetic resonance (EPR).

Figure 6.
Schematic representation of redox cycles for oxidation of substrates catalyzed by laccase.
Figure 6.
Schematic representation of redox cycles for oxidation of substrates catalyzed by laccase.

Figure 7.
Examples of laccases mediators. (a) 3-Hydroxyanthranilic acid (HAA); (b) 2,20-azino-bis-(3 ethylbenzothiazoline-6-sulphonic acid) (ABTS); (c) N-hydroxybenzotriazole (HBT); (d) N hydroxyphtaimide (HPI); (e) violuric acid (VLA); (f) N-hydroxyacetanilide (NHA); (g) methyl ester of 4 hydroxy-3,5-dimethoxy-benzoic acid (syringic acid); (h) 2,2,6,6-tetramethylpiperidine-1-yloxy (TEMPO).
Figure 7.
Examples of laccases mediators. (a) 3-Hydroxyanthranilic acid (HAA); (b) 2,20-azino-bis-(3 ethylbenzothiazoline-6-sulphonic acid) (ABTS); (c) N-hydroxybenzotriazole (HBT); (d) N hydroxyphtaimide (HPI); (e) violuric acid (VLA); (f) N-hydroxyacetanilide (NHA); (g) methyl ester of 4 hydroxy-3,5-dimethoxy-benzoic acid (syringic acid); (h) 2,2,6,6-tetramethylpiperidine-1-yloxy (TEMPO).

Figure 8.
Oxidation of ABTS catalyzed by laccase.

Figure 9.
Schematic representation of manganese peroxidase (MnP) catalyzed redox cycles for Mn2+.

Figure 10.
Commercial Lignin Transformations.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of various pretreatment methods.
| Pretreatment Methods | Feedstock | Sugar Yield | Advantages | Disadvantages | References |
|---|---|---|---|---|---|
| Biological | Softwood Wheat straw, rice straw | 20–50% Reducing sugar | Low energy consumption Cost effective Moderate reaction conditions | Required large sterile area Low hydrolysis rate | [12] |
| Ionic liquids (ILs) | Agricultural residuals: wheat straw, bagasse, corn stover, peanut and poplar sawdust | 60–85% Reducing sugars | Efficiently dissolution of cellulose | Great amounts of expensive ILs are needed Solutions viscous and difficult to handle | [13] |
| Ozonolysis | Agricultural residuals: wheat straw, bagasse, peanut and poplar sawdust | 45–90% Reducing sugars | Moderate reaction conditions Efficient lignin degradation | Costly protocol Requires large amount of ozone | [14] |
| Alkali pretreatment | Agricultural residuals: rice straw, wheat straw, woody material, sunflower stalk and corn stover | 65–85% Reducing sugars | Room temperature Destroy lignin | Less sugar degradation | [15] |
| Dilute acid | Agricultural residuals: wheat straw, and poplar sawdust | 45–80% Reducing sugars | Fast and do not need acid recycling | Formation of inhibitors Require high temperature and pressure | [16] |
| Concentrated acid | Agricultural residuals: wheat straw and bagasse | 60–90% Reducing sugars | High sugar conversion | Costly and need special reactors Highly corrosive and toxic | [17] |
| Organosolv | Agricultural residuals: wheat straw and sugarcane bagasse | Up to 60% of reducing sugars | Pure lignin removal as by-product Hydrolysis of lignin and hemicellulose | Costly process Requires recycling and drainage of solvents | [18] |
| Steam explosion | Agricultural residuals: wheat straw, corn stalk and sugarcane bagasse MSW Hardwood | 50–70% Reducing sugars | Cost effective Less hazardous process Lignin transformation Good sugar recovery | Incomplete destruction of lignin carbohydrate matrix Inhibitor compounds generation | [19] |
| Liquid hot water (LHW) | Agricultural residuals: wheat straw, corn stover, sunflower stalks and sugarcane bagasse | 80–94% Reducing sugars | Pure hemicellulose recovery No addition of catalysts High sugar recovery | High energy demand Dealing with left over solid mass | [20] |
| Extrusion | Agricultural residuals: wheat straw and rice straw | 50–75% Reducing sugars | Moderate temperature Good yield Less hazardous | Partially hemicellulose degradation Generation of inhibitors | [21] |
| Ammonia fiber explosion (AFEX) | Municipal solid waste Agricultural residuals: wheat straw, baggase and rice straw | Up to 80–90% of reducing sugars | Low formation of inhibitors Efficient lignin removal Moderate process conditions | Costly No efficiency with high lignin contents | [22] |
Table 2.
Acidophilic fungal strains [67].
Table 2.
Acidophilic fungal strains [67].
| Compounds | Fungal Strains |
|---|---|
| Ascomycota | |
| Itaconic acid | Aspergillus terreus |
| Fumaric acid | Aspergillus niger |
| Ascorbic acid | Aspergillus niger |
| Butyric acid | Aspergillus flavus |
| Isobutyric acid | Aspergillus niger |
| Malic acid | Aspergillus niger |
| Citric acid | Aspergillus niger |
| Succinic acid | Aspergillus flavipes |
| Lactic acid | Aspergillus niger |
| Oxalic acid | Aspergillus niger |
| Formic acid | Aspergillus flavipes |
| Acetic acid | Aspergillus niger |
| Propionic acid | Aspergillus niger |
| Gluconic acid | Aspergillus niger |
| Basidiomycota | |
| Gluconic acid | Pycnoporus coccineus |
| Oxalic acid | Ganoderma weberianum |
| Formic acid | Pycnoporus coccineus |
Table 3.
Common linkages in lignin [83].
Table 3.
Common linkages in lignin [83].
| Linkage Type | Share in Softwood Lignin (%) | Share in Hardwood Lignin (%) |
|---|---|---|
| β-O-4 | 45–50 | 60 |
| 5-5 | 10–27 | 3–9 |
| Β-5 | 9–12 | 6 |
| α-O-4 | 2–8 | 7 |
| β-β | 2–6 | 3–12 |
| β-1 | 7–10 | 1–7 |
| 4-O-5 | 4–8 | 7–9 |
| Dibenzodioxocin | 5–7 | 1–2 |
Table 4.
Functional groups in lignin [84].
Table 4.
Functional groups in lignin [84].
| Functional Groups | Abundance per 100 C9-units | |
|---|---|---|
| Softwood Lignin | Hardwood Lignin | |
| Methoxyl | 90–97 | 139–158 |
| Phenolic hydroxyl | 15–30 | 10–15 |
| Carbonyl | 10–20 | 17–24 |
| Aliphatic hydroxyl | 115–120 | 88–166 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Javaid, R.; Sabir, A.; Sheikh, N.; Ferhan, M. Recent Advances in Applications of Acidophilic Fungi to Produce Chemicals. Molecules 2019, 24, 786. https://doi.org/10.3390/molecules24040786
AMA Style
Javaid R, Sabir A, Sheikh N, Ferhan M. Recent Advances in Applications of Acidophilic Fungi to Produce Chemicals. Molecules. 2019; 24(4):786. https://doi.org/10.3390/molecules24040786
Chicago/Turabian StyleJavaid, Rehman, Aqsa Sabir, Nadeem Sheikh, and Muhammad Ferhan. 2019. "Recent Advances in Applications of Acidophilic Fungi to Produce Chemicals" Molecules 24, no. 4: 786. https://doi.org/10.3390/molecules24040786