The Importance of Brain Banks for Molecular Neuropathological Research: The New South Wales Tissue Resource Centre Experience

{kind=link}
Abstract
:1. Introduction
2. The New South Wales Tissue Resource Centre (TRC)
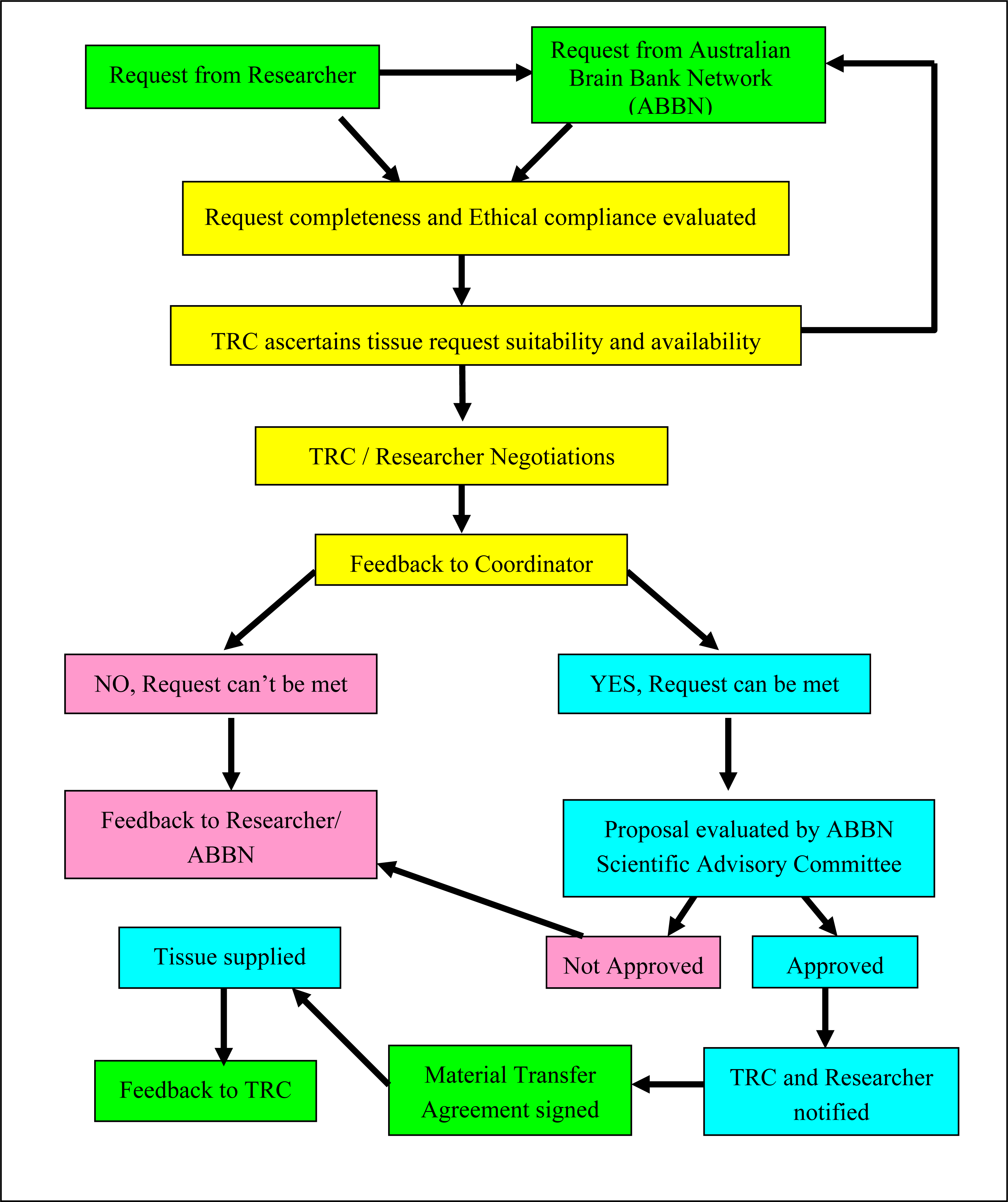
3. Changing nature of tissue requests and operational protocols at the TRC
4. Tissue integrity and clinical information quality assurance
4.1. Tissue integrity domain
4.2. Clinical domain
5. Evolution of the TRC
6. Australian Brain Bank Network
7. Specific examples of molecular neuropathological studies using TRC tissues
7.1. Schizophrenia
7.2. Alcohol use disorders
7.3. Movement disorders and dementias
8. Conclusions
Acknowledgments
References
- Boguski, M; Jones, A. Neurogenomics: At the intersection of neurobiology and genome sciences. Nat. Neurosci 2004, 7, 429–433. [Google Scholar]
- World Health Organization. Disease incidence, prevalence and disability. In World Health Organisation- Global Burden of Disease - 2004 update; 2008; pp. 28–37. [Google Scholar]
- Bell, JE; Alafuzoff, I; Al-Sarraj, S; Arzberger, T; Bogdanovic, N; Budka, H; Dexter, DT; Falkai, P; Ferrer, I; Gelpi, E; Gentleman, SM; Giaccone, G; Huitinga, I; Ironside, JW; Klioueva, N; Kovacs, GG; Meyronet, D; Palkovits, M; Parchi, P; Patsouris, E; Reynolds, R; Riederer, P; Roggendorf, W; Seilhean, D; Schmitt, A; Schmitz, P; Streichenberger, N; Schwalber, A; Kretzschmar, H. Management of a twenty-first century brain bank: experience in the BrainNet Europe consortium. Acta Neuropathol 2008, 115, 497–507. [Google Scholar]
- Schmitt, A; Bauer, M; Heinsen, H; Feiden, W; Falkai, P; Alafuzoff, I; Arzberger, T; Al-Sarraj, S; Bell, JE; Bogdanovic, N; Bruck, W; Budka, H; Ferrer, I; Giaccone, G; Kovacs, GG; Meyronet, D; Palkovits, M; Parchi, P; Patsouris, E; Ravid, R; Reynolds, R; Riederer, P; Roggendorf, W; Schwalber, A; Seilhean, D; Kretzschmar, H. How a neuropsychiatric brain bank should be run: A consensus paper of Brainnet Europe II. J. Neural. Transm 2007, 114, 527–537. [Google Scholar]
- Schmitt, A; Parlapani, E; Bauer, M; Heinsen, H; Falkai, P. Is brain banking of psychiatric cases valuable for neurobiological research? Clinics 2008, 63, 255–266. [Google Scholar]
- Kretzschmar, H. Brain banking: Opportunities, challenges and meaning for the future. Nat. Rev. Neurosci 2009, 10, 70–78. [Google Scholar]
- Sheedy, D; Garrick, T; Dedova, I; Hunt, C; Miller, R; Sundqvist, N; Harper, C. An Australian Brain Bank: A critical investment with a high return. Cell Tissue Bank 2008, 9, 205–216. [Google Scholar]
- Chevyreva, I; Faull, RL; Green, CR; Nicholson, LF. Assessing RNA quality in postmortem human brain tissue. Exp. Mol. Pathol 2008, 84, 71–77. [Google Scholar]
- Lipska, BK; Deep-Soboslay, A; Weickert, CS; Hyde, TM; Martin, CE; Herman, MM; Kleinman, JE. Critical factors in gene expression in postmortem human brain: Focus on studies in schizophrenia. Biol. Psychiatry 2006, 60, 650–658. [Google Scholar]
- Stan, AD; Ghose, S; Gao, XM; Roberts, RC; Lewis-Amezcua, K; Hatanpaa, KJ; Tamminga, CA. Human postmortem tissue: what quality markers matter? Brain. Res 2006, 1123, 1–11. [Google Scholar]
- Tomita, H; Vawter, MP; Walsh, DM; Evans, SJ; Choudary, PV; Li, J; Overman, KM; Atz, ME; Myers, RM; Jones, EG; Watson, SJ; Akil, H; Bunney, WE, Jr. Effect of agonal and postmortem factors on gene expression profile: quality control in microarray analyses of postmortem human brain. Biol. Psychiatry 2004, 55, 346–352. [Google Scholar]
- Webster, MJ. Tissue preparation and banking. Prog. Brain. Res 2006, 158, 3–14. [Google Scholar]
- Harrison, PJ; Heath, PR; Eastwood, SL; Burnet, PW; McDonald, B; Pearson, RC. The relative importance of premortem acidosis and postmortem interval for human brain gene expression studies: selective mRNA vulnerability and comparison with their encoded proteins. Neurosci. Lett 1995, 200, 151–154. [Google Scholar]
- Harper, C; Garrick, T; Matsumoto, I; Pfefferbaum, A; Adalsteinsson, E; Sullivan, E; Dodd, P; Lewohl, J; Butterworth, R. How important are brain banks for alcohol research? Alcohol. Clin. Exp. Res 2003, 27, 310–323. [Google Scholar]
- Hill, C; Roberts, S; Keks, N; Dean, B; Mackinnon, A; Copolov, D. Diagnostic Instrument for Brain Studies-Revised; Mental Health Research Institute: Victoria, Australia, 2005. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 4th Ed ed; American Psychiatric Association: Washington, DC, USA, 1994. [Google Scholar]
- Garrick, T; Glaw, X; Harper, CG; North, A. The diagnosis is critical for psychiatric brain banks: An interrater reliability study. Aust. N. Z. J. Psychiatry 2006, 40, 606–607. [Google Scholar]
- Sundqvist, N; Garrick, T; Bishop, I; Harper, C. Reliability of post-mortem psychiatric diagnosis for neuroscience research. Aust. N. Z. J. Psychiatry 2008, 42, 221–227. [Google Scholar]
- Flatscher-Bader, T; Matsumoto, I; Wilce, P. Smoking and alcoholism target genes associated with plasticity and glutamate transmission in the human VTA. Hum. Mol. Genet 2008, 17, 38–51. [Google Scholar]
- Flatscher-Bader, T; Wilce, PA. Chronic smoking and alcoholism change expression of selective genes in the human prefrontal cortex. Alcohol. Clin. Exp. Res 2006, 30, 908–915. [Google Scholar]
- Grinberg, LT; Ferretti, RE; Farfel, JM; Leite, R; Pasqualucci, CA; Rosemberg, S; Nitrini, R; Saldiva, PH; Filho, WJ. Brain bank of the Brazilian aging brain study group - a milestone reached and more than 1,600 collected brains. Cell Tissue Bank 2007, 8, 151–162. [Google Scholar]
- Vonsattel, JP; Del Amaya, MP; Keller, CE. Twenty-first century brain banking. Processing brains for research: The Columbia University methods. Acta Neuropathol 2008, 115, 509–532. [Google Scholar]
- Waldvogel, HJ; Bullock, JY; Synek, BJ; Curtis, MA; van Roon-Mom, WM; Faull, RL. The collection and processing of human brain tissue for research. Cell Tissue Bank 2008, 9, 169–179. [Google Scholar]
- Clark, DDI; Cordwell, S; Matsumoto, I. A proteome analysis of the anterior cingulate cortex gray matter in schizophrenia. Mol. Psychiatry 2006, 11, 459–470. [Google Scholar]
- Clark, D; Dedova, I; Cordwell, S; Matsumoto, I. Altered proteins of the anterior cingulate cortex white matter proteome in schizophrenia. Proteomics. Clin. Appl 2007, 1, 157–166. [Google Scholar]
- Sivagnanasundaram, S; Crossett, B; Dedova, I; Cordwell, S; Matsumoto, I. Abnormal pathways on the genu of the corpus callosum in schizophrenia pathogenesis: a proteome study. Proteomics. Clin. Appl 2007, 1, 1291–1305. [Google Scholar]
- Alexander-Kaufman, K; Dedova, I; Harper, C; Matsumoto, I. Proteome analysis of the dorsolateral prefrontal region from healthy individuals. Neurochem. Int 2007, 5, 433–439. [Google Scholar]
- Zavitsanou, K. M1 receptor agonism, a possible treatment for cognitive deficits in schizophrenia. Neuropsychopharmacology 2004, 29, 1585–1586. [Google Scholar]
- Zavitsanou, K; Garrick, T; Huang, X-F. Selective antagonist [3H]SR141716A binding to cannabinoid CB1 receptors is increased in the anterior cingulate cortex in schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2004, 28, 355–360. [Google Scholar]
- Zavitsanou, K; Katsifis, A; Mattner, F; Huang, X-F. Investigation of m1/m4 muscarinic receptors in the anterior cingulate cortex in schizophrenia, bipolar disorder, and major depression disorder. Neuropsychopharmacology 2004, 29, 619–625. [Google Scholar]
- Zavitsanou, K; Katsifis, A; Yu, Y; Huang, X-F. M2/M4 muscarinic receptor binding in the anterior cingulate cortex in schizophrenia and mood disorders. Brain. Res. Bull 2005, 65, 397–403. [Google Scholar]
- Zavitsanou, K; Ward, PB; Huang, X-F. Selective alterations in ionotropic glutamate receptors in the anterior cingulate cortex in schizophrenia. Neuropsychopharmacology 2002, 27, 826–833. [Google Scholar]
- Zavitsanou, K; Huang, X-F. Decreased [3H] spiperone binding in the anterior cingulate cortex of schizophrenia patients: An autoradiographic study. Neuroscience 2002, 109, 709–716. [Google Scholar]
- Ito, M; Depaz, I; Wilce, P; Suzuki, T; Niwa, S; Matsumoto, I. Expression of human neuronal protein 22, a novel cytoskeleton-associated protein, was decreased in the anterior cingulate cortex of schizophrenia. Neurosci. Lett 2005, 378, 125–130. [Google Scholar]
- Wheeler, DG; Harper, CG. Localised reductions in gyrification in the posterior cingulate: Schizophrenia and controls. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 319–327. [Google Scholar]
- Newell, KA; Deng, C; Huang, XF. Increased cannabinoid receptor density in the posterior cingulate cortex in schizophrenia. Exp. Brain. Res 2006, 172, 556–560. [Google Scholar]
- Newell, KA; Zavitsanou, K; Kum-Jew, S; Huang, X-F. Alterations of muscarinic and GABA receptor binding in the posterior cingulate cortex in schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 225–233. [Google Scholar]
- Newell, KA; Zavitsanou, K; Huang, X-F. Ionotropic glutamate receptor binding in the posterior cingulate cortex in schizophrenia patients. Neuroreport 2005, 16, 1363–1367. [Google Scholar]
- Tooney, PA; Au, GG; Chahl, LA. Tachykinin NK1 and NK3 receptors in the prefrontal cortex of the human brain. Clin. Exp. Pharmacol. Physiol 2000, 27, 947–949. [Google Scholar]
- Tooney, PA; Au, GG; Chahl, LA. Localisation of tachykinin NK1 and NK3 receptors in the human prefrontal and visual cortex. Neurosci. Lett 2000, 283, 185–188. [Google Scholar]
- Tooney, PA; Crawter, VC; Chahl, LA. Increased tachykinin NK(1) receptor immunoreactivity in the prefrontal cortex in schizophrenia. Biol. Psychiatry 2001, 49, 523–527. [Google Scholar]
- Ide, M; Ohnishi, T; Murayama, M; Matsumoto, I; Yamada, K; Iwayama, Y; Dedova, I; Toyota, T; Asada, T; Takashima, A; Yoshikawa, T. Failure to support a genetic contribution of AKT1 polymorphisms and altered AKT signaling in schizophrenia. J. Neurochem 2006, 99, 277–287. [Google Scholar]
- Tooney, PA; Chahl, LA. Neurons expressing calcium-binding proteins in the prefrontal cortex in schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2004, 28, 273–278. [Google Scholar]
- Wheeler, DG; Dixon, G; Harper, CG. No differences in calcium-binding protein immunoreactivity in the posterior cingulate and visual cortex: schizophrenia and controls. Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 630–639. [Google Scholar]
- Deng, C; Huang, X-F. Decreased density of muscarinic receptors in the superior temporal gyrusin schizophrenia. J. Neurosci. Res 2005, 81, 883–890. [Google Scholar]
- Deng, C; Huang, X-F. Increased density of GABAA receptors in the superior temporal gyrus in schizophrenia. Exp. Brain. Res 2006, 168, 587–590. [Google Scholar]
- Bowden, N; Scott, R; Tooney, P. Altered gene expression in the superior temporal gyrus in schizophrenia. BMC Genomics 2008, 9, 199. [Google Scholar]
- Bowden, NA; Scott, RJ; Tooney, PA. Altered expression of regulator of G-protein signalling 4 (RGS4) mRNA in the superior temporal gyrus in schizophrenia. Schizophr. Res 2007, 89, 165–168. [Google Scholar]
- Beveridge, N; Tooney, P; Carroll, A; Gardiner, E; Bowden, N; Scott, R; Tran, N; Dedova, I; Cairns, M. Dysregulation of miRNA 181b in the temporal cortex in schizophrenia. Hum. Mol. Genet 2008, 17, 1156–1168. [Google Scholar]
- Kapoor, R; Lim, K; Cheng, A; Garrick, T; Kapoor, V. Preliminary evidence for a link between schizophrenia and NMDA-glycine site receptor ligand metabolic enzymes, D-amino acid oxidase (DAAO) and kyurenine aminotransferase-1 (KAT-1). Brain. Res 2006, 1106, 205–210. [Google Scholar]
- Weidenhofer, J; Bowden, NA; Scott, RJ; Tooney, PA. Altered gene expression in the amygdala in schizophrenia: up-regulation of genes located in the cytomatrix active zone. Mol. Cell. Neurosci 2006, 31, 243–250. [Google Scholar]
- Weidenhofer, J; Yip, J; Zavitsanou, K; Huang, X-F; Chahl, LA; Tooney, PA. Immunohistochemical localisation of the NK1 receptor in the human amygdala: preliminary investigation in schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 1313–1321. [Google Scholar]
- Fan, L; Bellinger, F; Ge, Y-L; Wilce, P. Genetic study of alcoholism and novel gene expression in the alcoholic brain. Addict Biol 2004, 9, 11–18. [Google Scholar]
- Liu, J; Lewohl, JM; Dodd, PR; Randall, PK; Harris, RA; Mayfield, RD. Gene expression profiling of individual cases reveals consistent transcriptional changes in alcoholic human brain. J. Neurochem 2004, 90, 1050–1058. [Google Scholar]
- Lewohl, JM; Wang, L; Miles, MF; Zhang, L; Dodd, PR; Harris, RA. Gene expression in human alcoholism: microarray analysis of frontal cortex. Alcohol. Clin. Exp. Res 2000, 24, 1873–1882. [Google Scholar]
- Lewohl, JM; Dodd, PR; Mayfield, RD; Harris, RA. Application of DNA microarrays to study human alcoholism. J. Biomed. Sci 2001, 8, 28–36. [Google Scholar]
- Mayfield, RD; Lewohl, JM; Dodd, PR; Herlihy, A; Liu, J; Harris, RA. Patterns of gene expression are altered in the frontal and motor cortices of human alcoholics. J. Neurochem 2002, 81, 802–813. [Google Scholar]
- Liu, J; Lewohl, J; Harris, RA; Iyer, V; Dodd, P; Randall, PK; Mayfield, RD. Patterns of gene expression in the frontal cortex discriminate alcoholic from nonalcoholic individuals. Neuropsychopharmacology 2006, 31, 1574–1582. [Google Scholar]
- Johansson, S; Ekstrom, T; Marinova, Z; Okvist, A; Sheedy, D; Garrick, T; Harper, C; Kuzmin, A; Yakovleva, T; Bakalkin, G. Dysregulation of cell death machinery in the prefrontal cortex of human alcoholics. Int. J. Neuropsychopharmacol 2008, 12, 1–7. [Google Scholar]
- Harper, C; Dixon, G; Sheedy, D; Garrick, T. Neuropathological alterations in alcoholic brains. Studies arising from the New South Wales Tissue Resource Centre. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 951–961. [Google Scholar]
- Kril, JJ; Halliday, GM; Svoboda, MD; Cartwright, H. The cerebral cortex is damaged in chronic alcoholics. Neuroscience 1997, 79, 983–998. [Google Scholar]
- Flatscher-Bader, T; van der Brug, M; Hwang, JW; Gochee, PA; Matsumoto, I; Niwa, S; Wilce, PA. Alcohol-responsive genes in the frontal cortex and nucleus accumbens of human alcoholics. J. Neurochem 2005, 93, 359–370. [Google Scholar]
- Flatscher-Bader, T; van der Brug, MP; Landis, N; Hwang, JW; Harrison, E; Wilce, PA. Comparative gene expression in brain regions of human alcoholics. Genes. Brain. Behav 2006, 5, 78–84. [Google Scholar]
- Johansson, S; Fuchs, A; Okvist, A; Karimi, M; Harper, C; Garrick, T; Sheedy, D; Hurd, Y; Bakalkin, G; Ekstrom, T. Validation of endogenous controls for quantitative gene expression analysis: application on brain cortices of human chronic alcoholics. Brain. Res 2007, 1132, 20–28. [Google Scholar]
- Mayfield, RD; Liu, J; Randall, PK; Lewohl, JM; Dodd, PR; Harris, RA. Methods for the identification of differentially expressed genes in human post-mortem brain. Methods 2003, 31, 301–305. [Google Scholar]
- Liu, J; Lewohl, JM; Harris, RA; Dodd, PR; Mayfield, RD. Altered gene expression profiles in the frontal cortex of cirrhotic alcoholics. Alcohol. Clin. Exp. Res 2007, 31, 1460–1466. [Google Scholar]
- Okvist, A; Johansson, S; Kuzmin, A; Bazov, I; Merino-Martinez, R; Ponomarev, I; Mayfield, RD; Harris, RA; Sheedy, D; Garrick, T; Harper, C; Hurd, Y; Terenius, L; Ekstrom, T; Bakalkin, G; Yakovleva, T. Neuroadaptions in Human Chronic Alcoholics: Dysregulation of the NF-KB System. PLoS ONE 2007, 2, e930. [Google Scholar]
- Henriksson, R; Kuzmin, A; Okvist, A; Harper, C; Sheedy, D; Garrick, T; Yakovleva, T; Bakalkin, G. Elevated Synaptophysin I in the Prefrontal Cortex of Human Chronic Alcoholics. Synapse 2008, 62, 829–833. [Google Scholar]
- Lewohl, JM; Van Dyk, DD; Craft, GE; Innes, DJ; Mayfield, RD; Cobon, G; Harris, RA; Dodd, PR. The application of proteomics to the human alcoholic brain. Ann. N. Y. Acad. Sci 2004, 1025, 14–26. [Google Scholar]
- Alexander-Kaufman, K; Cordwell, S; Harper, C; Matsumoto, I. A proteome analysis of the dorsolateral prefrontal cortex in human alcoholic patients. Proteomics. Clin. Appl 2007, 1, 62–72. [Google Scholar]
- Alexander-Kaufman, K; James, G; Sheedy, D; Harper, C; Matsumoto, I. Differential protein expression in the prefrontal white matter of human alcoholics: a proteomics study. Mol. Psychiatry 2006, 11, 56–65. [Google Scholar]
- Alexander-Kaufman, K; Harper, C. Transketolase: Observations in alcohol-related brain damage research. Biochem. Cell. Biol. 2008. doi:10.1016/j.biocel.2008.04.005. [Google Scholar]
- Depaz, I; Ito, M; Matsumoto, I; Niwa, S; Kroon, P; Wilce, PA. Expression of hNP22 is altered in the frontal cortex and hippocampus of the alcoholic human brain. Alcohol. Clin. Exp. Res 2003, 27, 1481–1488. [Google Scholar]
- Kashem, M; James, G; Harper, C; Wilce, P; Matsumoto, I. Differential protein expression in the corpus callosum (splenium) of human alcoholics: A proteomics study. Neurochem. Int 2007, 50, 450–459. [Google Scholar]
- Kashem, MA; Harper, C; Matsumoto, I. Differential protein expression in the corpus callosum (Genu) of human alcoholics. Neurochem. Int 2008, 53, 1–11. [Google Scholar]
- Matsuda-Matsumoto, H; Iwazaki, T; Kashem, MA; Harper, C; Matsumoto, I. Differential protein expression profiles in the hippocampus of human alcoholics. Neurochem. Int 2007, 50, 450–459. [Google Scholar]
- Pamphlett, R; Kum-Jew, S; King, NJC. Flaviviruses in motor neuron disease. Muscle. Nerve 2005, 32, 108–109. [Google Scholar]
- Morahan, J; Yu, B; Trent, R; Pamphlett, R. Are metallothionein genes silenced in ALS? Toxicol. Lett 2006, 168, 83–87. [Google Scholar]
- Oates, N; Pamphlett, P. An epigenetic analysis of SOD1 and VEGF in ALS. Amyotroph. Lateral. Scler 2007, 8, 83–86. [Google Scholar]
- Pamphlett, R; Kum Jew, S. TDP-43 inclusions do not protect motor neurons from sporadic ALS. Acta. Neuropathol 2008, 116, 221–222. [Google Scholar]
- Saunderson, R; Yu, B; Trent, R; Pamphlett, R. A comparison of the lengths of androgen receptor triplet repeats in brain and blood in motor neuron diseases. J. Neurol. Sci 2008, 267, 125–128. [Google Scholar]
- Ruff, M; Pamphlett, R. A simple method for comparing microarray genotype data between brain and other tissues. J. Neurosci. Methods 2008, 173, 315–317. [Google Scholar]
- Fedorow, H; Halliday, GM; Rickert, CH; Gerlach, M; Riederer, P; Double, KL. Evidence for specific phases in the development of human neuromelanin. Neurobiol. Aging 2006, 27, 506–512. [Google Scholar]
- Halliday, G; Fedorow, H; Rickert, CH; Gerlach, M; Reiderer, P; Double, KL. Evidence for specific phases in the development of human neuromelanin. Neutral. Transm 2006, 113, 721–728. [Google Scholar]
- Cullen, KM. Perivascular astrocytes within Alzheimer’s disease plaques. Neuroreport 1997, 8, 1961–1966. [Google Scholar]
- Cullen, KM; Halliday, GM. Neurofibrillary degeneration and cell loss in the nucleus basalis n comparison to cortical Alzheimer pathology. Neurobiol. Aging 1998, 19, 297–306. [Google Scholar]
- Laws, SM; Hone, E; Taddei, K; Harper, C; Dean, B; McClean, C; Masters, C; Lautenschlager, N; Gandy, SE; Martins, RN. Variation at the APOE -491 promoter locus is associated with altered brain levels of apolipoprotein E. Mol. Psychiatry 2002, 7, 886–890. [Google Scholar]
- Kwok, J; Loy, C; Hamilton, G; Lau, E; Hallupp, M; Williams, J; Owen, M; Broe, A; Tang, N; Lam, L; Powell, J; Lovestone, S; Schofield, P. Glycogen synthase kinase-3β and Tau Genes Interact in Alzheimer’s disease. Ann. Neurol 2008, 64, 446–454. [Google Scholar]
- Sokolova, A; Hill, M; Rahimi, F; Warden, L; Halliday, G; Shepherd, C. Monocyte Chemoattractant Protein-1 plays a dominant role in the chronic inflammation observed in Alzheimer’s disease. Brain. Pathol. 2008, 1–7. [Google Scholar]
- Sutherland, GT; Nowak, G; Halliday, GM; Kril, J. Tau isoform expression in frontotemporal dementia without tau deposition. J. Clin. Neurosci 2007, 14, 1182–1185. [Google Scholar]

© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dedova, I.; Harding, A.; Sheedy, D.; Garrick, T.; Sundqvist, N.; Hunt, C.; Gillies, J.; Harper, C.G. The Importance of Brain Banks for Molecular Neuropathological Research: The New South Wales Tissue Resource Centre Experience. Int. J. Mol. Sci. 2009, 10, 366-384. https://doi.org/10.3390/ijms10010366
Dedova I, Harding A, Sheedy D, Garrick T, Sundqvist N, Hunt C, Gillies J, Harper CG. The Importance of Brain Banks for Molecular Neuropathological Research: The New South Wales Tissue Resource Centre Experience. International Journal of Molecular Sciences. 2009; 10(1):366-384. https://doi.org/10.3390/ijms10010366
Chicago/Turabian StyleDedova, Irina, Antony Harding, Donna Sheedy, Therese Garrick, Nina Sundqvist, Clare Hunt, Juliette Gillies, and Clive G. Harper. 2009. "The Importance of Brain Banks for Molecular Neuropathological Research: The New South Wales Tissue Resource Centre Experience" International Journal of Molecular Sciences 10, no. 1: 366-384. https://doi.org/10.3390/ijms10010366
APA StyleDedova, I., Harding, A., Sheedy, D., Garrick, T., Sundqvist, N., Hunt, C., Gillies, J., & Harper, C. G. (2009). The Importance of Brain Banks for Molecular Neuropathological Research: The New South Wales Tissue Resource Centre Experience. International Journal of Molecular Sciences, 10(1), 366-384. https://doi.org/10.3390/ijms10010366