Investigation on the Protective Effect of α-Mannan against the DNA Damage Induced by Aflatoxin B1 in Mouse Hepatocytes

 ,
,  ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Chemicals and animals
2.2. Genotoxicity/antigenotoxicity protocol
2.2.1. Experimental design
2.3. Unicellular alkaline electrophoresis (comet) assay
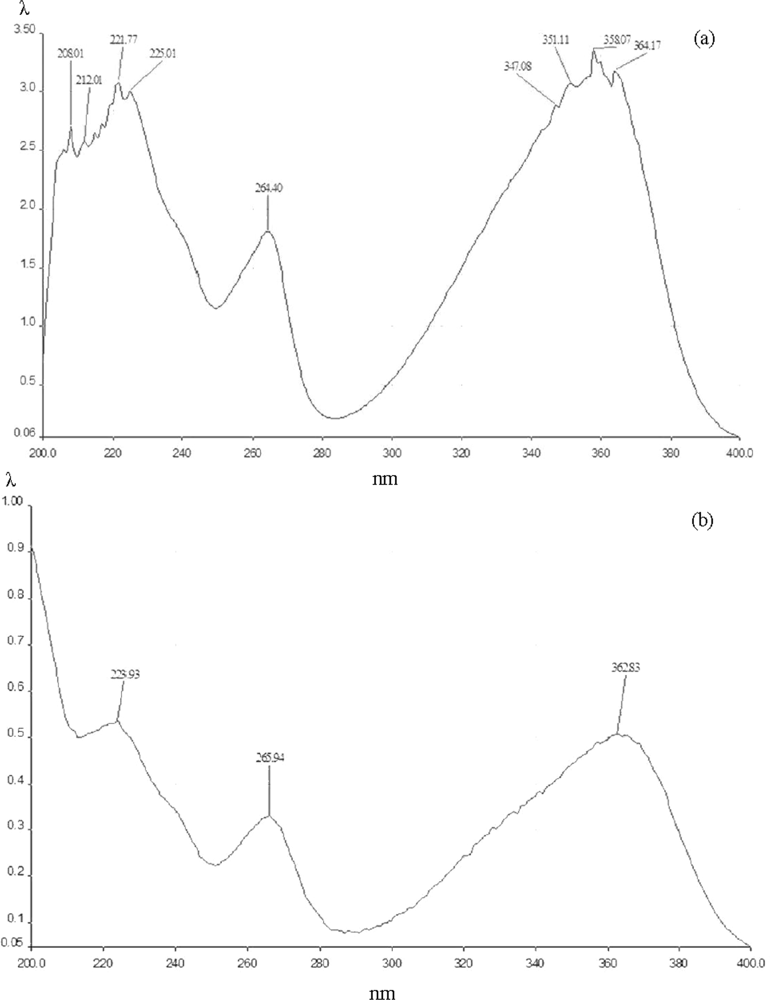
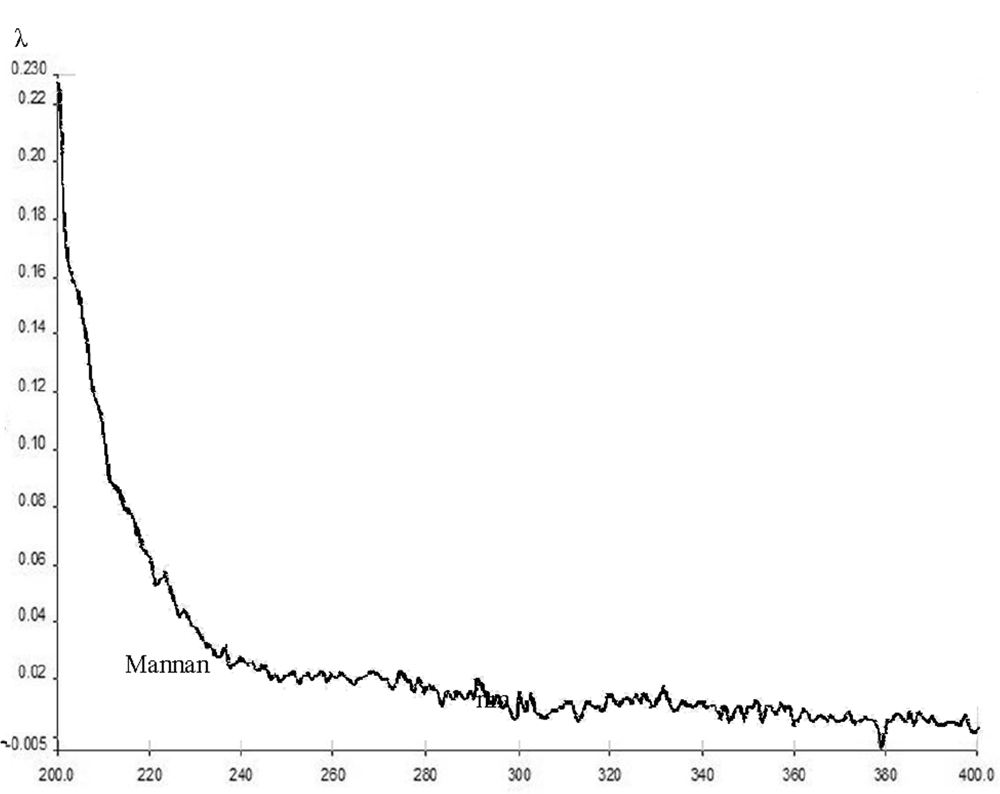
2.4. Crystallization, melting point determination, and spectroscopic analysis
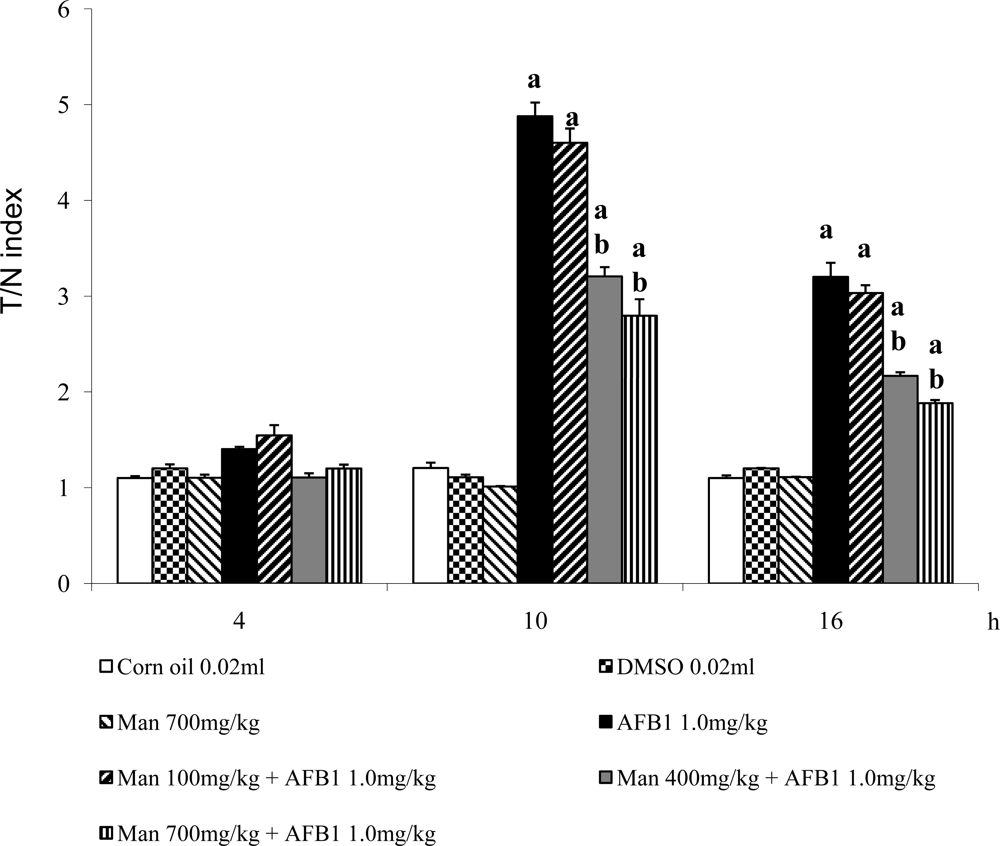
3. Results and Discussion
4. Conclusions
References
- Wang, JS; Groopman, JD. DNA Damage by Mycotoxins. Mutat. Res 1999, 424, 167–181. [Google Scholar]
- Witt, KL; Knapton, A; Wehr, CM; Hook, GJ; Mirsalis, J; Shelby, MD; MacGregor, JT. Micronucleated Erythrocyte Frequency in Peripheral Blood of B6C3F(1) Mice from Short-Term, Prechronic, and Chronic Studies of the NTP Carcinogenesis Bioassay Program. Environ. Mol. Mutagen 2000, 36, 163–194. [Google Scholar]
- Steyn, PS. Mycotoxins, General View, Chemistry and Structure. Toxicol Lett 1995, 82–83, 843–851. [Google Scholar]
- Cleveland, TE; Dowd, PE; Desjardins, AE; Bhatnagar, D; Coty, PJ. United States Department of Agriculture-Agricultural Research Service. Research on Preharvest Prevention of Mycotoxins and Mycotoxigenic Fungi in US Crops. Pest. Manag. Sci 2003, 59, 629–642. [Google Scholar]
- Kumagai, S; Nakajima, M; Tabata, S; Ishikuro, E; Tanaka, T; Norizuki, H; Itoh, Y; Aoyama, K; Fujita, K; Kai, S; Sato, T; Yoshiike, N; Sugita-Konishi, Y. Aflatoxin and ochratoxin. A Contamination of Retail Foods and Intake of These Mycotoxins in Japan. Food Add Contam 2008, 1–6. [Google Scholar]
- Wang, J; Liu, XM. Contamination of Aflatoxins in Different Kinds of Foods in China. Biomed. Environ. Sci 2007, 20, 483–487. [Google Scholar]
- Samarajeewa, U; Sen, AC; Cohen, MA; Wei, CI. Detoxification of Aflatoxins in Foods and Feeds by Physical and Chemical Methods. J Food Prot 1990, 53, 489–501. [Google Scholar]
- Robb, J. Mycotoxins: Contamination and Decontamination. Feed Mix 1993, 1, 18–23. [Google Scholar]
- Madrigal-Santillan, E; Madrigal-Bujaidar, E; Marquez-Marquez, R; Reyes, A. Antigenotoxic Effect of Saccharomyces cerevisiae on the Damage Produced in Mice Fed with Aflatoxin B1 Contaminated Corn. Food Chem. Toxicol 2006, 44, 2058–2063. [Google Scholar]
- Kogan, G; Pajtinka, M; Babincova, M; Miadoková, E; Rauko, P; Slamenova, D; Korolenko, TA. Yeast Cell Wall Polysaccharides as Antioxidants and Antimutagens: Can They Fight Cancer? Neoplasma 2008, 55, 387–393. [Google Scholar]
- Madrigal-Santillan, E; Alvarez-Gonzalez, I; Marquez-Marquez, R; Velazquez-Guadarrama, N; Madrigal-Bujaidar, E. Inhibitory Effect of Mannan on the Toxicity Produced in Mice Fed Aflatoxin B1 Contaminated Corn. Arch. Environ. Contam. Toxicol 2007, 53, 466–472. [Google Scholar]
- Križková, L; Ďuracková, Z; Šandula, J; Sasinková, V; Krajčovič, J. Antioxidative and Antimutagenic Activity of Yeast Cell Wall Mannans in Vitro. Mutat. Res 2001, 497, 213–222. [Google Scholar]
- Hartmann, A; Agurell, E; Beevers, C; Brendler-Schaab, S; Burlinson, B; Clay, P; Collins, A; Smith, A; Speit, G; Thybaud, V; Tice, RR. Recommendations for Conducting the in Vivo Alkaline Comet Assay. Mutagenesis 2003, 18, 45–51. [Google Scholar]
- Ralph, S; Petras, M. Comparison of Sensitivity to Methyl Methanesulphonate Among Tadpole Developmental Stages Using the Alkaline Single-Cell Gel Electrophoresis (Comet) Assay. Environ. Mol. Mutagen 1998, 31, 374–382. [Google Scholar]
- Castillo-Cadena, J; Tenorio-Vieyra, LE; Quintana-Carabia, AI; Garcia-Fabila, MM; Ramirez-San Juan, E; Madrigal-Bujaidar, E. Determination of DNA Damage in Floriculturists Exposed to Mixtures of Pesticides. J Biomed Biotechnol 2006, 2006(2), 1–12. [Google Scholar]
- Bergfors, TM. Protein Crystallization Technique Strategies and Tips A Laboratory Manual; International University Line: La Jolla, California, USA, 2001. [Google Scholar]
- Tice, RR. The single cell gel comet assay A microgel electrophoretic technique for the detection of DNA damage and repair in individual cells. In Environmental mutagenesis; Phillips, DH, Venitt, S, Eds.; Bios Scientific Publishers: Oxford, UK, 1995; pp. 315–339. [Google Scholar]
- Miadoková, E; Svidová, S; Ulčová, V; Dǜhová, V; Nad’ová, S; Rauko, P; Kogan, G. Diverse Biomodulatory Effects of Glucomannan from Candida utilis. Toxicol. In Vitro 2006, 20, 649–657. [Google Scholar]
- Friedman Angeli, JP; Ribeiro, LJ; Ferreira Bellini, M; Mantovani, MS. β-Glucan Extracted from the Medicinal Mushroom Agaricus blazei Prevents the Genotoxic Effects of Benzo(a)pyrene in the Human Hepatoma Cell Line HepG2. Arch Toxicol 2008. [Google Scholar]
- Tohamy, AA; El Ghor, AA; El Nahas, SM; Noshy, MM. β-Glucan Inhibits the Genotoxicity of Cyclophosphamide, Adriamycin and Cisplatin. Mutat. Res 2003, 541, 45–53. [Google Scholar]
- Toklu, HS; Öser Seirli, A; Velioğlu-Öğunc, A; Centinel, S; Sener, G. Acetaminophen-Induced Toxicity is Prevented by β-D-Glucan Treatment in Mice. Eur J Pharmacol 2006, 543, 133–140. [Google Scholar]
- Babincova, M; Bacova, Z; Machova, E; Kogan, G. Antioxidant Properties of Carboxymethyl Glucan: Comparative Analysis. J. Med. Food 2002, 5, 79–83. [Google Scholar]
- Križkova, L; Žitňanová, I; Mislovičová, D; Masárová, J; Sasinková, V; Ďuračková, Z; Krajčovič. Antioxidant and Antimutagenic Activity of Mannan Neoglycoconjugates: Mannan-Human Serum Albumine and Mannan-Penicillin G Acylase. Mutat Res 2006, 606, 72–79. [Google Scholar]
- Toklu, HZ; Şener, G; Jahovic, N; Uslu, B; Arbak, S; Yeğen, B. B-Glucan Protects Against Burn-Induced Oxidative Organ Damage in Rats. Intern. Immunopharmacol 2006, 6, 156–169. [Google Scholar]
- Weisburger, JH. Antimutagens, Anticarcinogens, and Effective Worldwide Cancer Prevention. J. Environ. Pathol. Toxicol. Oncol 1999, 18, 85–93. [Google Scholar]
- Weisburger, JH. Lifestyle, Health and Disease Prevention: The Underlying Mechanisms. Eur J Cancer Prev 2002, (Suppl 2), 51–57. [Google Scholar]
- Rao, CV; Chou, D; Simi, B; Ku, H; Reddy, BS. Prevention of Colonic Aberrant Crypt Foci and Modulation of Large Bowel Microbial Activity by Dietary Coffee Fiber, Inulin and Pectin. Carcinogenesis 1998, 19, 1815–1819. [Google Scholar]
- Glei, M; Hofman, T; Küster, K; Hollmann, J; Linhahuer, MG; Pool-Zobel, BL. Both Weath (Triticum aestivum) Bran Arabinoxylans and Gut Flora-Mediated Fermentation Products Protect Human Colon Cells from Genotoxic Activities of 4-Hydroxynonenal and Hydrogen Peroxide. J. Agric. Food Chem 2006, 54, 2088–2095. [Google Scholar]
- Miranda, DDC; Arçari, DP; Ladeira, MSP; Calori-Dominguez, MA; Romero, AC; Salvadori, DMF; Gloria, EM; Pedrazzoli, J, Jr; Ribeiro, ML. Analysis of DNA Damage Induced by Aflatoxin B1 in Dunkin-Hartley Guinea Pigs. Mycopathologia 2007, 163, 275–280. [Google Scholar]
- Anderson, D; Yu, T-W; Hambly, RJ; Mendy, M; Wild, CP. Aflatoxin Exposure and DNA Damage in the Comet Assay in Individuals from the Gambia, West Africa. Teratog, Carcinog, Mutagen 1999, 19, 147–155. [Google Scholar]
- Steed, JW; Turner, DR; Wallace, KJ. Core Concepts in Supramolecular Chemistry and Nanochemistry; John Wiley & Sons, Ltd: Chichester, UK, 2007; pp. 5–25. [Google Scholar]
- Reyes-Arellano, A; Rose, R; Steller, I; Sustmann, R. Crystal engineering: Supramolecular Structures by Cocrystallization of meso-1,2-diphenyl-1,2-ethanediol and bisimines. Simple Cases of Moleular Recognition. J. Struct. Chem 1995, 6, 391–396. [Google Scholar]
- Reyes, A; Nájera, J; Mendoza, R; Molins, I; Grau, E; Espinosa, E; Salgado, H; Tamariz, J; Soriano, M; Paneerselvam, K. Bisimines in Supramolecular Synthesis. Supramol Chem 1999, 11, 17–24. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Agent/dose | Time (h) | Grades of damage (%) | |||
|---|---|---|---|---|---|
| G0 | G1 | G2 | G3 | ||
| Corn oil
0.02 mL | 4 | 84 | 13 | 2 | 1 |
| 10 | 87 | 11 | 2 | 0 | |
| 16 | 92 | 4 | 3 | 1 | |
| DMSO
0.02 mL | 4 | 77 | 17 | 4 | 2 |
| 10 | 87 | 5 | 4 | 4 | |
| 16 | 86 | 8 | 4 | 2 | |
| Man
700 mg/kg | 4 | 91 | 7 | 2 | 0 |
| 10 | 85 | 10 | 3 | 2 | |
| 16 | 89 | 6 | 3 | 2 | |
| AFB1 1.0 mg/kg | 4 | 72 | 12 | 8 | 8 |
| 10 | 2 | 9 | 24 | 65 | |
| 16 | 18 | 18 | 27 | 37 | |
| Man + AFB1 100 + 1.0 mg/kg | 4 | 67 | 8 | 10 | 15 |
| 10 | 4 | 10 | 29 | 57 | |
| 16 | 16 | 20 | 30 | 34 | |
| Man + AFB1 400 + 1.0 mg/kg | 4 | 75 | 12 | 8 | 5 |
| 10 | 23 | 20 | 23 | 35 | |
| 16 | 32 | 15 | 25 | 28 | |
| Man + AFB1 700 + 1.0 mg/kg | 4 | 80 | 14 | 3 | 3 |
| 10 | 42 | 18 | 17 | 23 | |
| 16 | 36 | 28 | 19 | 17 | |
| Chemical | Melting point (°C) |
|---|---|
| AFB1/Man | 182.5 |
| AFB1 | 132.5 |
| Man | 246.0 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Madrigal-Santillán, E.; Morales-González, J.A.; Sánchez-Gutiérrez, M.; Reyes-Arellano, A.; Madrigal-Bujaidar, E. Investigation on the Protective Effect of α-Mannan against the DNA Damage Induced by Aflatoxin B1 in Mouse Hepatocytes. Int. J. Mol. Sci. 2009, 10, 395-406. https://doi.org/10.3390/ijms10020395
Madrigal-Santillán E, Morales-González JA, Sánchez-Gutiérrez M, Reyes-Arellano A, Madrigal-Bujaidar E. Investigation on the Protective Effect of α-Mannan against the DNA Damage Induced by Aflatoxin B1 in Mouse Hepatocytes. International Journal of Molecular Sciences. 2009; 10(2):395-406. https://doi.org/10.3390/ijms10020395
Chicago/Turabian StyleMadrigal-Santillán, Eduardo, José Antonio Morales-González, Manuel Sánchez-Gutiérrez, Alicia Reyes-Arellano, and Eduardo Madrigal-Bujaidar. 2009. "Investigation on the Protective Effect of α-Mannan against the DNA Damage Induced by Aflatoxin B1 in Mouse Hepatocytes" International Journal of Molecular Sciences 10, no. 2: 395-406. https://doi.org/10.3390/ijms10020395