Heterogeneity of Mitochondria and Mitochondrial Function within Cells as Another Level of Mitochondrial Complexity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Overview of Existing Results and Their Discussion
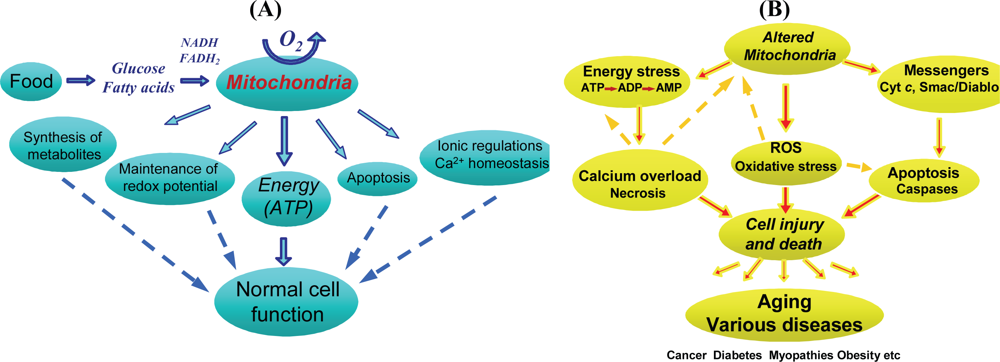
2.1. Multiple functions of mitochondria in the cell
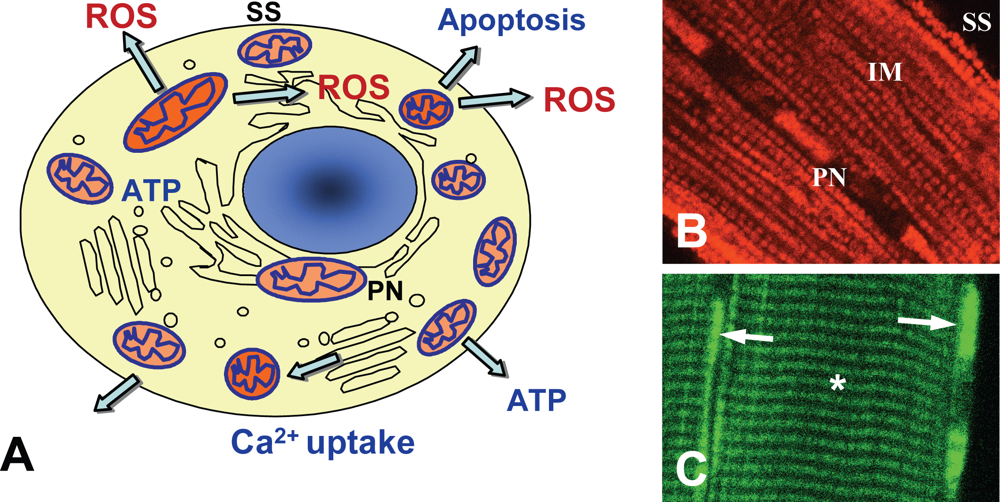
2.2. Heterogeneity of mitochondrial morphology and organization
2.3. The connectivity of mitochondria
2.4. Functional heterogeneity of mitochondria
2.5. Possible physiological roles of distinct mitochondrial subpopulations
2.6. Heterogeneity of mitochondrial ROS
2.7. Heterogeneity of mitochondria in pathology
2.8. Heterogeneity of mitochondria in apoptosis
2.9. Possible cellular mechanisms causing mitochondrial heterogeneity
2.10. Various levels of mitochondrial heterogeneity
3. Conclusions
Acknowledgments
References
- Anmann, T; Eimre, M; Kuznetsov, AV; Andrienko, T; Kaambre, T; Sikk, P; Seppet, E; Tiivel, T; Vendelin, M; Seppet, E; Saks, VA. Calcium-Induced contraction of sarcomeres changes the regulation of mitochondrial respiration in permeabilized cardiac cells. FEBS J 2005, 272, 3145–3161. [Google Scholar]
- Appaix, F; Kuznetsov, AV; Usson, Y; Kay, L; Andrienko, T; Olivares, J; Kaambre, T; Sikk, P; Margreiter, R; Saks, V. Possible role of cytoskeleton in intracellular arrangement and regulation of mitochondria. Exp. Physiol 2003, 88, 175–190. [Google Scholar]
- Collins, TJ; Berridge, MJ; Lipp, P; Bootman, MD. Mitochondria are morphologically and functionally heterogeneous within cells. EMBO J 2002, 21, 1616–1627. [Google Scholar]
- Huang, S; Heikal, AA; Webb, WW. Two-photon fluorescence spectroscopy and microscopy of NAD(P)H and flavoprotein. Biophys. J 2002, 82, 2811–2825. [Google Scholar]
- Kuznetsov, AV; Mayboroda, O; Kunz, D; Winkler, K; Schubert, W; Kunz, WS. Functional imaging of mitochondria in saponin-permeabilized mice muscle fibers. J. Cell Biol 1998, 140, 1091–1099. [Google Scholar]
- Kuznetsov, AV; Troppmair, J; Sucher, R; Hermann, M; Saks, V; Margreiter, R. Mitochondrial subpopulations and heterogeneity revealed by confocal imaging: Possible physiological role? Biochim. Biophys. Acta 2006, 1757, 686–691. [Google Scholar]
- Egner, A; Jakobs, S; Hell, SW. Fast 100-nm resolution three-dimensional microscope reveals structural plasticity of mitochondria in live yeast. Proc. Natl. Acad. Sci. USA 2002, 99, 3370–3375. [Google Scholar]
- Plecita-Hlavata, L; Lessard, M; Santorova, J; Bewersdorf, J; Jezek, P. Mitochondrial oxidative phosphorylation and energetic status are reflected by morphology of mitochondrial network in INS-1E and HEP-G2 cells viewed by 4Pi microscopy. Biochim. Biophys. Acta 2008, 1777, 834–846. [Google Scholar]
- Vendelin, M; Beraud, N; Guerrero, K; Andrienko, T; Kuznetsov, AV; Olivares, J; Kay, L; Saks, VA. Mitochondrial regular arrangement in muscle cells: A “crystal-like” pattern. Am. J. Physiol. Cell Physiol 2005, 288, C757–C767. [Google Scholar]
- Romashko, DN; Marban, E; O’rourke, B. subcellular metabolic transients and mitochondrial redox waves in heart cells. Proc. Natl. Acad. Sci. USA 1998, 95, 1618–1623. [Google Scholar]
- Bruce, JI; Giovannucci, DR; Blinder, G; Shuttleworth, TJ; Yule, DI. Modulation of [Ca2+]i signaling dynamics and metabolism by perinuclear mitochondria in mouse parotid acinar cells. J. Biol. Chem 2004, 279, 12909–12917. [Google Scholar]
- Collins, TJ; Lipp, P; Berridge, MJ; Bootman, MD. Mitochondria are morphologically and functionally heterogeneous within single cells. J. Physiol. (London) 2002, 539, 98P–99P. [Google Scholar]
- Kuznetsov, AV; Usson, Y; Leverve, X; Margreiter, R. Subcellular heterogeneity of mitochondrial function and dysfunction: evidence obtained by confocal imaging. Mol Cell Biochem 2004, 256–257, 359–365. [Google Scholar]
- Park, MK; Ashby, MC; Erdemli, G; Petersen, OH; Tepikin, AV. Perinuclear, perigranular and sub-plasmalemmal mitochondria have distinct functions in the regulation of cellular calcium transport. EMBO J 2001, 20, 1863–1874. [Google Scholar]
- Hollenbeck, PJ; Saxton, WM. The axonal transport of mitochondria. J. Cell Sci 2005, 118, 5411–5419. [Google Scholar]
- Beraud, N; Pelloux, S; Usson, Y; Kuznetsov, AV; Ronot, X; Tourneur, Y; Saks, V. Mitochondrial dynamics in heart cells: Very low amplitude high frequency fluctuations in adult cardiomyocytes and flow motion in HL-1 cells. J. Bioenerg. Biomem 2009. [Google Scholar]
- Anesti, V; Scorrano, L. The relationship between mitochondrial shape and function and the cytoskeleton. Biochim. Biophys. Acta 2006, 1757, 692–699. [Google Scholar]
- Capetanaki, Y. Desmin cytoskeleton: a potential regulator of muscle mitochondrial behavior and function. Trends Cardiovasc. Med 2002, 12, 339–348. [Google Scholar]
- Rizzuto, R; Pinton, P; Carrington, W; Fay, FS; Fogarty, KE; Lifshitz, LM; Tuft, RA; Pozzan, T. Close contacts with the endoplasmic reticulum as determinants of mitochondrial Ca2+ responses. Science 1998, 280, 1763–1766. [Google Scholar]
- Saks, V; Dzeja, P; Schlattner, U; Vendelin, M; Terzic, A; Wallimann, T. Cardiac system bioenergetics: metabolic basis of the frank-starling law. J. Physiol. (London) 2006, 571, 253–273. [Google Scholar]
- Saks, VA; Kaambre, T; Sikk, P; Eimre, M; Orlova, E; Paju, K; Piirsoo, A; Appaix, F; Kay, L; Regitz-Zagrosek, V; Fleck, E; Seppet, E. Intracellular energetic units in red muscle cells. Biochem. J 2001, 356, 643–657. [Google Scholar]
- Seppet, EK; Kaambre, T; Sikk, P; Tiivel, T; Vija, H; Tonkonogi, M; Sahlin, K; Kay, L; Appaix, F; Braun, U; Eimre, M; Saks, VA. Functional complexes of mitochondria with Ca, MgATPases of myofibrils and sarcoplasmic reticulum in Muscle Cells. Biochim. Biophys. Acta 2001, 1504, 379–395. [Google Scholar]
- Kay, L; Li, Z; Mericskay, M; Olivares, J; Tranqui, L; Fontaine, E; Tiivel, T; Sikk, P; Kaambre, T; Samuel, JL; Rappaport, L; Usson, Y; Leverve, X; Paulin, D; Saks, VA. Study of regulation of mitochondrial respiration in vivo. An analysis of influence of ADP diffusion and possible role of cytoskeleton. Biochim. Biophys. Acta 1997, 1322, 41–59. [Google Scholar]
- Milner, DJ; Mavroidis, M; Weisleder, N; Capetanaki, Y. Desmin cytoskeleton linked to muscle mitochondrial distribution and respiratory function. J. Cell Biol 2000, 150, 1283–1298. [Google Scholar]
- Rostovtseva, TK; Sheldon, KL; Hassanzadeh, E; Monge, C; Saks, V; Bezrukov, SM; Sackett, DL. Tubulin binding blocks mitochondrial voltage-dependent anion channel and regulates respiration. Proc. Natl. Acad. Sci. USA 2008, 105, 18746–18751. [Google Scholar]
- Saks, V; Kuznetsov, A; Andrienko, T; Usson, Y; Appaix, F; Guerrero, K; Kaambre, T; Sikk, P; Lemba, M; Vendelin, M. Heterogeneity of ADP diffusion and regulation of respiration in cardiac cells. Biophys. J 2003, 84, 3436–3456. [Google Scholar]
- Andrienko, T; Kuznetsov, AV; Kaambre, T; Usson, Y; Orosco, A; Appaix, F; Tiivel, T; Sikk, P; Vendelin, M; Margreiter, R; Saks, VA. Metabolic consequences of functional complexes of mitochondria, myofibrils and sarcoplasmic reticulum in muscle cells. J. Exp. Biol 2003, 206, 2059–2072. [Google Scholar]
- McBride, HM; Neuspiel, M; Wasiak, S. Mitochondria: More than just a powerhouse. Curr. Biol 2006, 16, R551–R560. [Google Scholar]
- Kroemer, G; Reed, JC. Mitochondrial control of cell death. Nat. Med 2000, 6, 513–519. [Google Scholar]
- Newmeyer, DD; Ferguson-Miller, S. Mitochondria: Releasing power for life and unleashing the machineries of death. Cell 2003, 112, 873. [Google Scholar]
- Karbowski, M; Norris, KL; Cleland, MM; Jeong, SY; Youle, RJ. Role of bax and bak in mitochondrial morphogenesis. Nature 2006, 443, 658–662. [Google Scholar]
- Kuznetsov, AV; Janakiraman, M; Margreiter, R; Troppmair, J. Regulating cell survival by controlling cellular energy production: Novel functions for ancient signaling pathways? FEBS Lett 2004, 577, 1–4. [Google Scholar]
- Kuznetsov, AV; Smigelskaite, J; Doblander, C; Janakiraman, M; Hermann, M; Wurm, M; Scheidl, SF; Sucher, R; Deutschmann, A; Troppmair, J. Survival signaling by C-RAF: mitochondrial reactive oxygen species and Ca2+ are critical targets. Mol. Cell Biol 2008, 28, 2304–2313. [Google Scholar]
- Riedl, SJ; Salvesen, GS. The apoptosome: signalling platform of cell death. Nat. Rev. Mol. Cell Biol 2007, 8, 405–413. [Google Scholar]
- Youle, RJ. Cellular demolition and the rules of engagement. Science 2007, 315, 776–777. [Google Scholar]
- Pelloux, S; Robillard, J; Ferrera, R; Bilbaut, A; Ojeda, C; Saks, V; Ovize, M; Tourneur, Y. Non-beating HL-1 cells for confocal microscopy: application to mitochondrial functions during cardiac preconditioning. Progr. Biophys. Mol. Biol 2006, 90, 270–298. [Google Scholar]
- Dzeja, PP; Bortolon, R; Perez-Terzic, C; Holmuhamedov, EL; Terzic, A. Energetic communication between mitochondria and nucleus directed by catalyzed phosphotransfer. Proc. Natl. Acad. Sci. USA 2002, 99, 10156–10161. [Google Scholar]
- Ball, EH; Singer, SJ. Mitochondria are associated with microtubules and not with intermediate filaments in cultured fibroblasts. Proc. Natl. Acad. Sci. USA 1982, 79, 123–126. [Google Scholar]
- Mose-Larsen, P; Bravo, R; Fey, SJ; Small, JV; Celis, JE. Putative association of mitochondria with a subpopulation of intermediate-sized filaments in cultured human skin fibroblasts. Cell 1982, 31, 681–692. [Google Scholar]
- Heggeness, MH; Simon, M; Singer, SJ. Association of mitochondria with microtubules in cultured cells. Proc. Natl. Acad. Sci. USA 1978, 75, 3863–3866. [Google Scholar]
- Belikova, YO; Kuznetsov, AV; Saks, VA. Specific limitations for intracellular diffusion of adp in cardiomyocytes. Biochemistry (Moscow) 1990, 55, 1450–1460. [Google Scholar]
- Tang, HL; Lung, HL; Wu, KC; Le, AHP; Tang, HM; Fung, MC. Vimentin Supports Mitochondrial Morphology and Organization. Biochem. J 2008, 410, 141–146. [Google Scholar]
- Winter, L; Abrahamsberg, C; Wiche, G. Plectin isoform 1b mediates mitochondrion-intermediate lament network linkage and controls organelle shape. J. Cell Biol 2008, 181, 903–911. [Google Scholar]
- Kaasik, A; Veksler, V; Boehm, E; Novotova, M; Minajeva, A; Ventura-Clapier, R. Energetic crosstalk between organelles: Architectural integration of energy production and utilization. Circ. Res 2001, 89, 153–159. [Google Scholar]
- Collins, TJ; Bootman, MD. Mitochondria are morphologically heterogeneous within cells. J. Exp. Biol 2003, 206, 1993–2000. [Google Scholar]
- Zorov, DB; Filburn, CR; Klotz, LO; Zweier, JL; Sollott, SJ. Reactive oxygen species (ros)-induced ros release: A new phenomenon accompanying induction of the mitochondrial permeability transition in cardiac myocytes. J. Exp. Med 2000, 192, 1001–1014. [Google Scholar]
- De Giorgi, F; Lartigue, L; Ichas, F. Electrical coupling and plasticity of the mitochondrial network. Cell Calc 2000, 28, 365–370. [Google Scholar]
- Amchenkova, AA; Bakeeva, LE; Chentsov, YS; Skulachev, VP; Zorov, DB. Coupling membranes as energy-transmitting cables. I. Filamentous mitochondria in fibroblasts and mitochondrial clusters in cardiomyocytes. J. Cell Biol 1988, 107, 481–495. [Google Scholar]
- Cortassa, S; Aon, MA; Winslow, RL; O’Rourke, B. A mitochondrial oscillator dependent on reactive oxygen species. Biophys. J 2004, 87, 2060–2073. [Google Scholar]
- Knight, MM; Roberts, SR; Lee, DA; Bader, DL. Live cell imaging using confocal microscopy induces intracellular calcium transients and cell death. Am. J. Physiol. Cell Physiol 2003, 284, C1083–C1089. [Google Scholar]
- Wang, W; Fang, H; Groom, L; Cheng, A; Zhang, W; Liu, J; Wang, X; Li, K; Han, P; Zheng, M; Yin, J; Wang, W; Mattson, MP; Kao, JP; Lakatta, EG; Sheu, SS; Ouyang, K; Chen, J; Dirksen, RT; Cheng, H. Superoxide flashes in single mitochondria. Cell 2008, 134, 279–290. [Google Scholar]
- Palmer, JW; Tandler, B; Hoppel, CL. Biochemical differences between subsarcolemmal and interfibrillar mitochondria from rat cardiac muscle: Effects of procedural manipulations. Arch. Biochem. Biophys 1985, 236, 691–702. [Google Scholar]
- Lesnefsky, EJ; Tandler, B; Ye, J; Slabe, TJ; Turkaly, J; Hoppel, CL. Myocardial ischemia decreases oxidative phosphorylation through cytochrome oxidase in subsarcolemmal mitochondria. Am. J. Physiol 1997, 273, H1544–H1554. [Google Scholar]
- Jimenez, M; Yvon, C; Lehr, L; Leger, B; Keller, P; Russell, A; Kuhne, F; Flandin, P; Giacobino, JP; Muzzin, P. Expression of uncoupling protein-3 in subsarcolemmal and intermyofibrillar mitochondria of various mouse muscle types and its modulation by fasting. Eur. J. Biochem 2002, 269, 2878–2884. [Google Scholar]
- Kuznetsov, AV; Schneeberger, S; Renz, O; Meusburger, H; Saks, V; Usson, Y; Margreiter, R. Functional heterogeneity of mitochondria after cardiac cold ischemia and reperfusion revealed by confocal imaging. Transplantation 2004, 77, 754–756. [Google Scholar]
- Kuznetsov, AV; Schneeberger, S; Leverve, X; Margreiter, R. Functional heterogeneity of mitochondria in normal cells and after ischemia/reperfusion revealed by confocal imaging. Shock 2005, 23, 36–38. [Google Scholar]
- Salvioli, S; Dobrucki, J; Moretti, L; Troiano, L; Fernandez, MG; Pinti, M; Pedrazzi, J; Franceschi, C; Cossarizza, A. Mitochondrial heterogeneity during staurosporine-induced apoptosis in HL60 cells: Analysis at the single cell and single organelle level. Cytometry 2000, 40, 189–197. [Google Scholar]
- Benard, G; Rossignol, R. Ultrastructure of the mitochondrion and its bearing on function and bioenergetics. Antioxid. Redox Signal 2008, 10, 1313–1342. [Google Scholar]
- Bowser, DN; Minamikawa, T; Nagley, P; Williams, DA. Role of mitochondria in calcium regulation of spontaneously contracting cardiac muscle cells. Biophys. J 1998, 75, 2004–2014. [Google Scholar]
- Kristian, T; Weatherby, TM; Bates, TE; Fiskum, G. Heterogeneity of the calcium-induced permeability transition in isolated non-synaptic brain mitochondria. J. Neurochem 2002, 83, 1297–1308. [Google Scholar]
- Chen, H; Chomyn, A; Chan, DC. Disruption of fusion results in mitochondrial heterogeneity and dysfunction. J. Biol. Chem 2005, 280, 26185–26192. [Google Scholar]
- Lawrie, AM; Rizzuto, R; Pozzan, T; Simpson, AW. A role for calcium influx in the regulation of mitochondrial calcium in endothelial cells. J. Biol. Chem 1996, 271, 10753–10759. [Google Scholar]
- Echtay, KS; Roussel, D; St Pierre, J; Jekabsons, MB; Cadenas, S; Stuart, JA; Harper, JA; Roebuck, SJ; Morrison, A; Pickering, S; Clapham, JC; Brand, MD. Superoxide activates mitochondrial uncoupling proteins. Nature 2002, 415, 96–99. [Google Scholar]
- Miwa, S; Brand, MD. Mitochondrial matrix reactive oxygen species production is very sensitive to mild uncoupling. Biochem. Soc. Trans 2003, 31, 1300–1301. [Google Scholar]
- Turrens, JF. Mitochondrial formation of reactive oxygen species. J. Physiol 2003, 552, 335–344. [Google Scholar]
- Mollica, MP; Lionetti, L; Crescenzo, R; D’Andrea, E; Ferraro, M; Liverini, G; Iossa, S. Heterogeneous bioenergetic behaviour of subsarcolemmal and intermyofibrillar mitochondria in fed and fasted rats. Cell Mol. Life Sci 2006, 63, 358–366. [Google Scholar]
- Iossa, S; Lionetti, L; Mollica, MP; Crescenzo, R; Botta, M; Samec, S; Dulloo, AG; Liverini, G. Differences in proton leak kinetics, but not in UCP3 protein content, in subsarcolemmal and intermyofibrillar skeletal muscle mitochondria from fed and fasted rats. FEBS Lett 2001, 505, 53–56. [Google Scholar]
- Balaban, RS; Nemoto, S; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar]
- Kudin, AP; Bimpong-Buta, NY; Vielhaber, S; Elger, CE; Kunz, WS. Characterization of superoxide-producing sites in isolated brain mitochondria. J. Biol. Chem 2004, 279, 4127–4135. [Google Scholar]
- Li, C; Jackson, RM. Reactive species mechanisms of cellular hypoxia-reoxygenation injury. Am. J. Physiol. Cell Physiol 2002, 282, C227–C241. [Google Scholar]
- Akar, FG; Aon, MA; Tomaselli, GF; O’Rourke, B. The mitochondrial origin of postischemic arrhythmias. J. Clin. Invest 2005, 115, 3527–3535. [Google Scholar]
- Otani, H. Reactive oxygen species as mediators of signal transduction in ischemic preconditioning. Antioxid. Redox Signal 2004, 6, 449–469. [Google Scholar]
- Pinton, P; Rimessi, A; Marchi, S; Orsini, F; Migliaccio, E; Giorgio, M; Contursi, C; Minucci, S; Mantovani, F; Wieckowski, MR; Del Sal, G; Pelicci, PG; Rizzuto, R. Protein kinase C beta and prolyl isomerase 1 regulate mitochondrial effects of the life-span determinant p66shc. Science 2007, 315, 659–663. [Google Scholar]
- Wall, JA; Wei, J; Ly, M; Belmont, P; Martindale, JJ; Tran, D; Sun, J; Chen, WJ; Yu, W; Oeller, P; Briggs, S; Gustafsson, AB; Sayen, MR; Gottlieb, RA; Glembotski, CC. Alterations in oxidative phosphorylation complex proteins in the hearts of transgenic mice that overexpress the P38 MAP kinase activator, MAP kinase kinase 6. Am. J. Physiol. Heart Circ. Physiol 2006, 291, H2462–H2472. [Google Scholar]
- Bensaad, K; Tsuruta, A; Selak, MA; Vidal, MN; Nakano, K; Bartrons, R; Gottlieb, E; Vousden, KH. TIGAR, a P53-inducible regulator of glycolysis and apoptosis. Cell 2006, 126, 107–120. [Google Scholar]
- Hajek, P; Villani, G; Attardi, G. Rate-limiting step preceding cytochrome c release in cells primed for fas-mediated apoptosis revealed by analysis of cellular mosaicism of respiratory changes. J. Biol. Chem 2001, 276, 606–615. [Google Scholar]
- Kuznetsov, AV; Schneeberger, S; Seiler, R; Brandacher, G; Mark, W; Steurer, W; Saks, V; Usson, Y; Margreiter, R; Gnaiger, E. Mitochondrial defects and heterogeneous cytochrome c release after cardiac cold ischemia and reperfusion. Am. J. Physiol. Heart Circ. Physiol 2004, 286, H1633–H1641. [Google Scholar]
- Shoubridge, EA. Mitochondrial DNA diseases: Histological and cellular studies. J. Bioenerg. Biomem 1994, 26, 301–310. [Google Scholar]
- Hoppel, CL; Moghaddas, S; Lesnefsky, EJ. Interfibrillar cardiac mitochondrial comples III defects in the aging rat heart. Biogerontology 2002, 3, 41–44. [Google Scholar]
- Leist, M; Single, B; Castoldi, AF; Kuhnle, S; Nicotera, P. Intracellular adenosine triphosphate (ATP) concentration: a switch in the decision between apoptosis and necrosis. J. Exp. Med 1997, 185, 1481–1486. [Google Scholar]
- Wikstrom, JD; Katzman, SM; Mohamed, H; Twig, G; Graf, SA; Heart, E; Molina, AJA; Corkey, BE; de Vargas, LM; Danial, NN; Collins, S; Shirihai, OS. Beta-cell mitochondria exhibit membrane potential heterogeneity that can be altered by stimulatory or toxic fuel levels. Diabetes 2007, 56, 2569–2578. [Google Scholar]
- Hermann, M; Kuznetsov, A; Maglione, M; Smigelskaite, J; Margreiter, R; Troppmair, J. Cytoplasmic signaling in the control of mitochondrial uproar? Cell Commun. Signal 2008, 6, 4. [Google Scholar]
- Mironov, SL; Symonchuk, N. ER vesicles and mitochondria move and communicate at synapses. J. Cell Sci 2006, 119, 4926–4934. [Google Scholar]
- Mironov, SL. ADP regulates movements of mitochondria in neurons. Biophys. J 2007, 92, 2944–2952. [Google Scholar]




© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kuznetsov, A.V.; Margreiter, R. Heterogeneity of Mitochondria and Mitochondrial Function within Cells as Another Level of Mitochondrial Complexity. Int. J. Mol. Sci. 2009, 10, 1911-1929. https://doi.org/10.3390/ijms10041911
Kuznetsov AV, Margreiter R. Heterogeneity of Mitochondria and Mitochondrial Function within Cells as Another Level of Mitochondrial Complexity. International Journal of Molecular Sciences. 2009; 10(4):1911-1929. https://doi.org/10.3390/ijms10041911
Chicago/Turabian StyleKuznetsov, Andrey V., and Raimund Margreiter. 2009. "Heterogeneity of Mitochondria and Mitochondrial Function within Cells as Another Level of Mitochondrial Complexity" International Journal of Molecular Sciences 10, no. 4: 1911-1929. https://doi.org/10.3390/ijms10041911