Time-Course Expression Profiles of Hair Cycle-Associated Genes in Male Mini Rats after Depilation of Telogen-Phase Hairs

Abstract
:1. Introduction
2. Results and Discussion
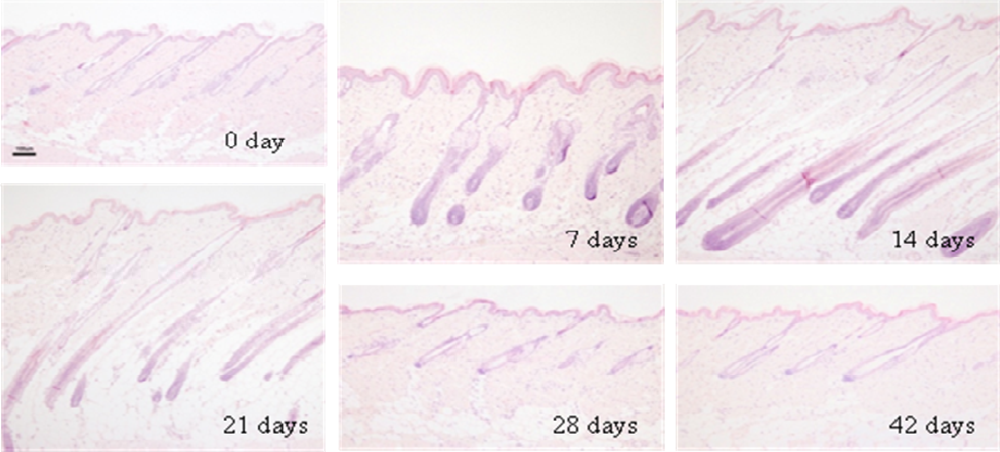
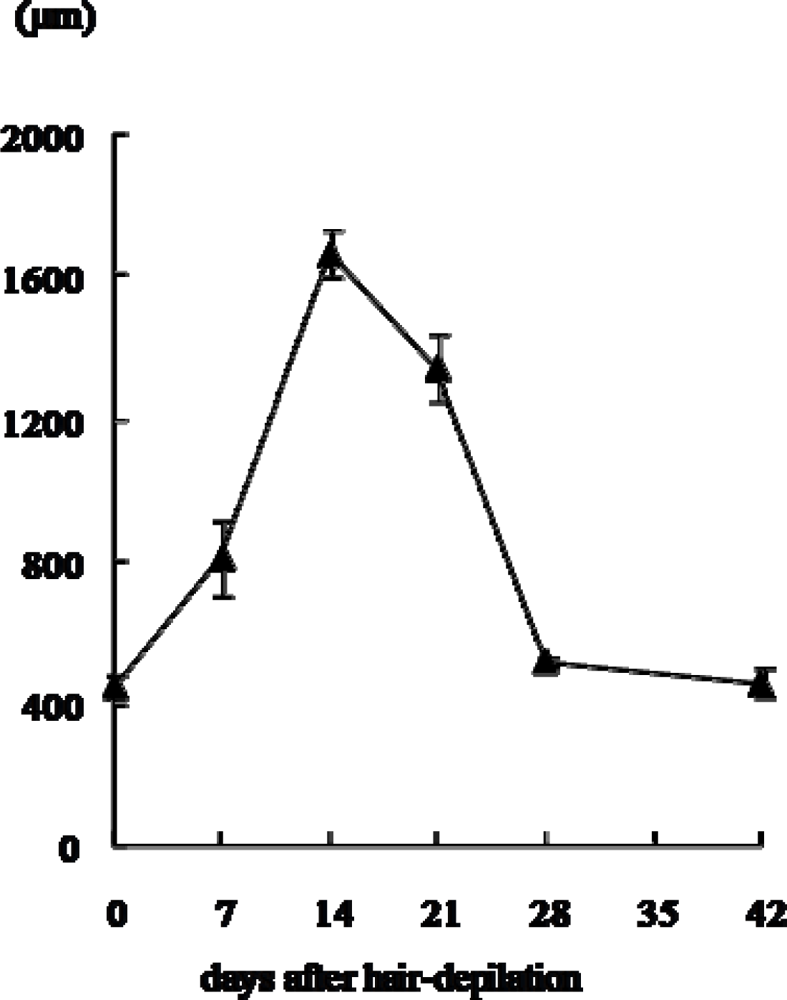
2.1. Changes in histology and depth (length) of hair follicles
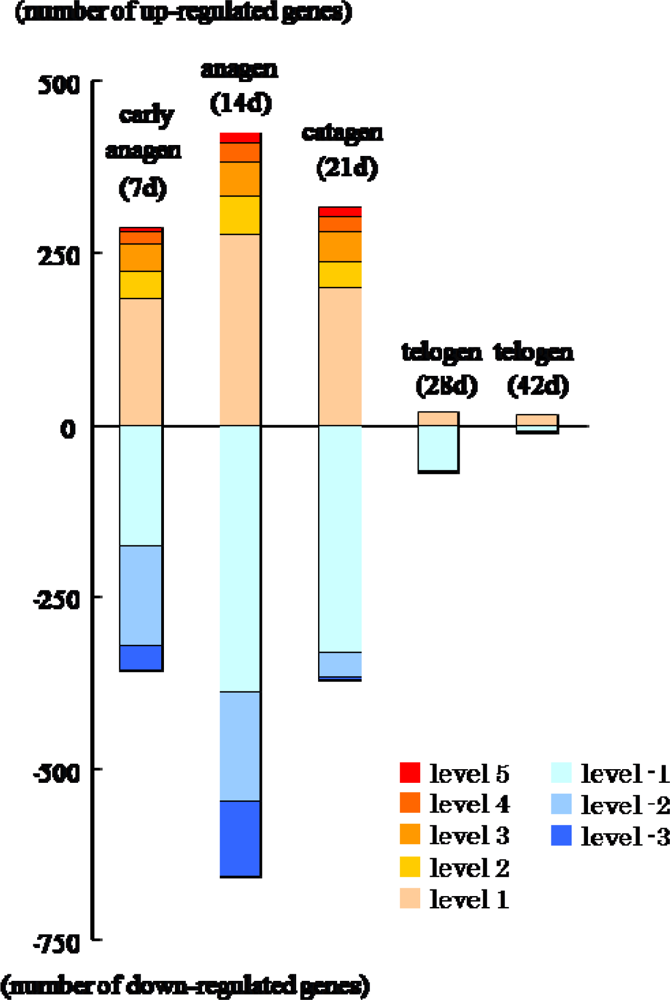
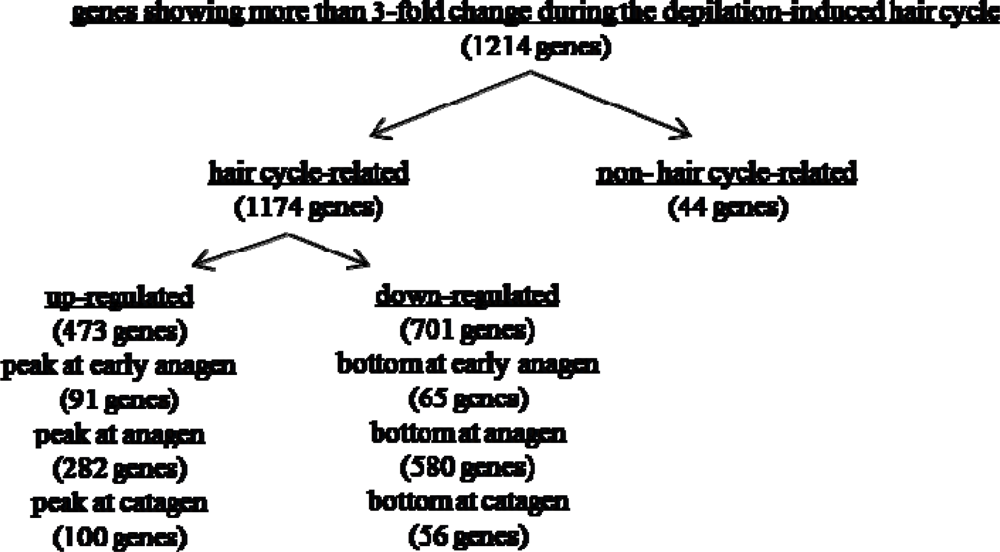
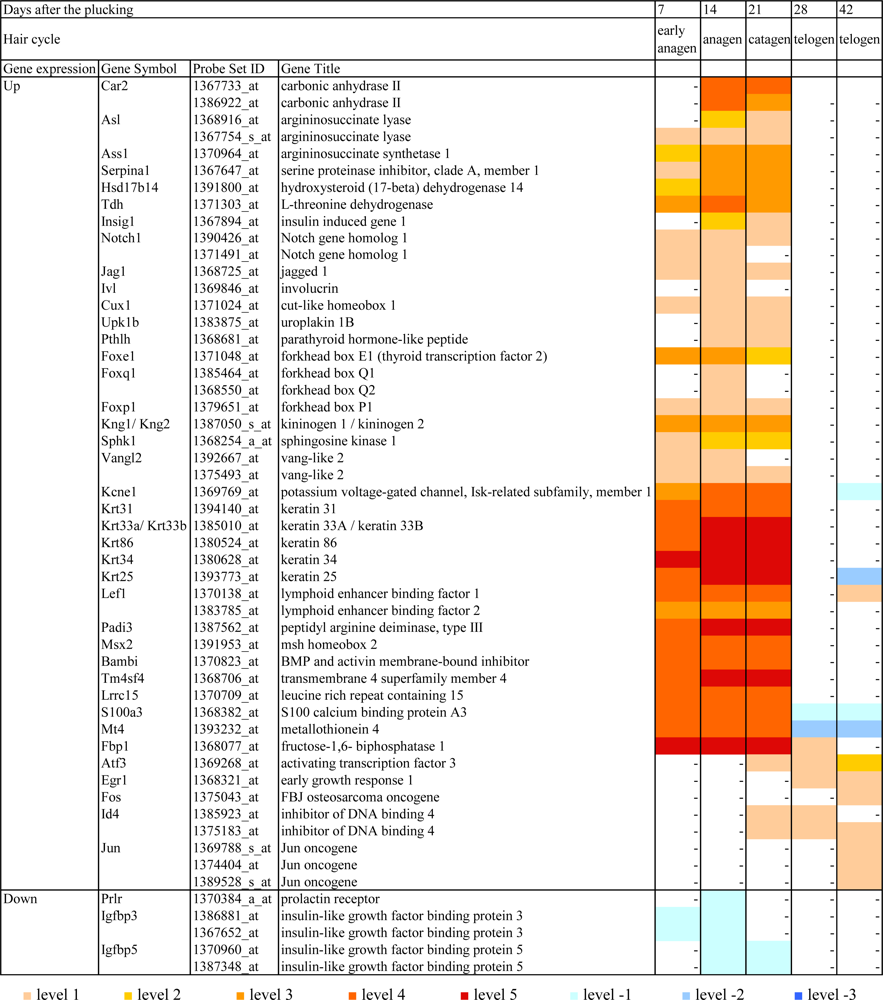
2.2. Gene expression profiles
3. Materials and Methods
3.1. Animals
3.2. Treatments
3.3. Histology
3.4. Microarray analysis
3.5. Microarray data analysis
4. Conclusions
Acknowledgments
References
- Matsumoto, K; Kakidani, H; Anzai, M; Nakagata, N; Takahashi, A; Takahashi, Y; Miyata, K. Evaluation of an antisense RNA transgene for inhibiting growth hormone gene expression in transgenic rats. Dev. Genet 1995, 16, 273–277. [Google Scholar]
- Matsumoto, K; Kakidani, H; Takahashi, A; Nakagata, N; Anzai, M; Matsuzaki, Y; Takahashi, Y; Miyata, K; Utsumi, K; Iritani, A. Growth retardation in rats whose growth hormone gene expression was suppressed by antsense RNA transgene. Mol. Reprod. Dev 1993, 36, 53–58. [Google Scholar]
- Ikawa, A; Ishii, Y; Suzuki, K; Yasoshima, A; Suzuki, N; Nakayama, H; Takahashi, S; Doi, K. Age-related changes in the dorsal skin histology in Mini and Wistar rats. Histol. Histopathol 2002, 17, 419–426. [Google Scholar]
- Muller-Rover, S; Handjiski, B; Van Der Veen, C; Eichmuller, S; Foitzik, K; McKay, IA; Stenn, KS; Paus, R. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair stages. J. Invest. Dermatol 2001, 117, 3–15. [Google Scholar]
- Umeda-Ikawa, A; Ishii, Y; Doi, K. Hair cycle induction by hair plucking in Mini rats. Exp. Mol. Pathol 2006, 80, 306–307. [Google Scholar]
- Umeda-Ikawa, A; Ishii, Y; Suzuki, K; Uetsuka, K; Nakayama, H; Doi, K. Dorsal skin responses to topical application with hydrogen peroxide in Mini and Wistar rats. Exp. Tox. Pathol 2002, 54, 239–244. [Google Scholar]
- Lin, KK; Chudova, D; Hatfield, GW; Smyth, P; Anderson, B. Identification of hair cycle-associated genes from time-course gene expression profile data by using replicate variance. Proc. Natl. Acad. Sci. USA 2004, 101, 15955–15960. [Google Scholar]
- Schlake, T; Beibel, M; Weger, N; Boehm, T. Major shift in genomic activity accompany progression through different atages of the hair cycle. Gene Expr. Patterns 2004, 4, 141–152. [Google Scholar]
- Ishimatsu-Tsuji, Y; Moro, O; Kishimoto, J. Expression profiling and cellular localization of genes associated with hair cycle induced by wax depilation. J. Invest. Dermatol 2005, 125, 410–420. [Google Scholar]
- Priestley, GC. Rates and duration of hair growth in the albino rat. J. Anat 1966, 100, 147–157. [Google Scholar]
- Stenn, KS; Paus, R. Controls of hair follicle cycling. Physiol. Rev 2001, 81, 449–494. [Google Scholar]
- Paus, R; Foitzik, K. In search of the “hair clock”: a guide tour. Differentiation 2004, 72, 489–511. [Google Scholar]
- Kizawa, K; Tsuchimoto, S; Hashimoto, K; Uchiwa, H. Gene expression of mouse S100A3, a cysteine-rich calcium-binding protein, in developing hair follicle. J. Invest. Dermatol 1998, 111, 879–886. [Google Scholar]
- Nachat, R; Mechin, MC; Charveron, M; Serre, G; Constans, J; Simon, M. Peptidylargiine deiminase isoforms are differentially expressed in the anagen hair follicles and other human skin appendages. J. Invest. Dermatol 2005, 125, 34–41. [Google Scholar]
- Weger, N; Schlake, T. IGF-1 signaling controls the hair growth cycle and the differentiation of hair shafts. J. Invest. Dermatol 2005, 125, 873–882. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Gene symbol |
|---|---|
| Genes up-regulated during the depilation-induced hair cycle | |
| Cell cycle/ tissue regeneration | 3 cyclins, 4 cell division cycles, 3 tubulins, 3 kinesin families, Tpd52l1, Pttg1, Nuf2, Tm4sf4, Tm4sf1 |
| Proliferation/differentiation/development | Cml5, Hspa2, Kng1, Sphk1, Mycn, Pthlh, Upk1b, Cux1, Tyrp1, Vangl2, Dlx3, Stmn1, Marcksl1, Pdlim7 |
| Keratin/ keratinocyte differentiation | 6 keratins, Sprr1al, Ivl, Notch1 |
| Translation | 5 eukaryotic translation initiation factors, Cars, Yars, Rpl7 |
| Regulation of transcription | Foxe1, Foxq1, Foxp1, Sox4, Csdc2, Top2a, Myo5a, Mafb, Trps1, Cited4, Dnmt1, E2f5, Fhl2, Hmgb2, Pcgf6 |
| Wnt/TGFβ signaling pathway | Lef1, Bambi, Msx1, Msx2, Bmp4, Csnk1e, Gpc3, Fzd1, Stra6, Mitf, Smad7, Sostdc1, Wnt4, Tcf7 |
| Transport | 11 solute carrier families, Abcg2, Fxyd4, Kcne1, Nup210, Selenbp1, Scg5, Ucp2, 2 ATPases, Bspry |
| Signal transduction | Itsn1, Gprc5a, Gpsm2, Ednrb, P2ry5, Ptger4, Gpr56, Nradd, Farp1, Mfhas1 |
| Amino acid metabolic process | Ggt1, Acy3, Gnmt, Cbs, Gss, Gclc, Cth, Otub2 |
| Carbohydrate metabolic process | Fbp1, 2 protein phosphatases, Hexb |
| Steroid hormone related | Hsd17b14, Tdh, Serpina1, Car2, Ass1, Asl, Arg1, Insig1, Adipor2, Fads3 |
| Cell Adhesion | 3 collagens, Dsc2, Gpnmb, Spon1 |
| Proteolysis | 3 cathepsins, Cpm, Capn8, Lap3, Pitrm1, Metap2, Prep, St14, Atg4b |
| Cell communication | Lrrc15, Gjb2, Gjb6, Jag1 |
| Protein modification process | Padi1, Padi3, Padi4 |
| Calcium ion binding | S100a3, S100a7a, Tesc, Tchh, Mt4 |
| Genes down-regulated during the depilation-induced hair cycle | |
| Cell cycle arrest | Ak1, Cgref1, Pmp22, Nbl1, Dst |
| Anti-apoptosis/regulation of apoptosis | Eef1a2, Sod2, Cryab, Tsc22d3, Vnn1, Cabc1, Cidea, Nol3, Cck |
| Negative regulation of cell growth | 4 insulin-like growth factor binding proteins, Dab2, Gas6 |
| Immune response | 5 chemokine ligands, 4 CD antigens, 4 Fc receptors, 4 RT1-class II, 7 complements, Il1b, Hla-dma |
| Regulation of transcription | Abra, Ldb3, Six1, Satb1, Cebpd, Dbp, Synpo2, Rxrg, Tcea3, Myf6, Tbx15, Lass5, Aebp1, Rora, Lrrfip1, Tnxa |
| Wnt/TGFβ signaling pathway | Cav1, Cav2, Cpz, Grem1, Grem2, Sfrp4 |
| Transport | 4 ATPases, 2 calcium channels, 3 sodium channels, 6 solute carrier families, 4 transmembrane proteins, Mup5, Fxyd1, Cp, Serpinh1, Ap1s2, Rbp7, Clic2 |
| Signal transduction | Hspb6, Agtr1a, Mfap4, Lpar1, Rgs10, Cxcr7, Plcb4, Obscn |
| Carbohydrate metabolic process | 5 protein phosphatases, 2 pyruvate dehydrogenase kinases, Pgm1, Phka1, Pygm, Gpd1, Gfpt2, Agl |
| Fatty acid biosynthetic/metabolic process | Ankrd23, Cpt1b, Fabp3, Acadm, Crot, Ptgs1, Phyh |
| Steroid biosynthetic/metabolic process | Ebp, Soat1, Hmgcr, Prlr, Prkaa2, Sult1a1, Vldlr |
| Cell adhesion | 6 collagens, Lox, Emb, Ddr2, Itgbl1, Epdr1, Hrc |
| Proteolysis | Mmp2, Ctsk, Cpa3, Mcpt1, Pcolce, Ctss, Dpp7, Pgcp |
| Muscle contraction | 9 myosin polypeptides, 6 troponins, Mybpc2, Mylk2, Myom1, Tmod1, Tpm1 |
| Genes up-regulated with aging | |
| Regulation of transcription | Jun, Fos, Atf3, Id4, Egr1 |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Umeda-Ikawa, A.; Shimokawa, I.; Doi, K. Time-Course Expression Profiles of Hair Cycle-Associated Genes in Male Mini Rats after Depilation of Telogen-Phase Hairs. Int. J. Mol. Sci. 2009, 10, 1967-1977. https://doi.org/10.3390/ijms10051967
Umeda-Ikawa A, Shimokawa I, Doi K. Time-Course Expression Profiles of Hair Cycle-Associated Genes in Male Mini Rats after Depilation of Telogen-Phase Hairs. International Journal of Molecular Sciences. 2009; 10(5):1967-1977. https://doi.org/10.3390/ijms10051967
Chicago/Turabian StyleUmeda-Ikawa, Aya, Isao Shimokawa, and Kunio Doi. 2009. "Time-Course Expression Profiles of Hair Cycle-Associated Genes in Male Mini Rats after Depilation of Telogen-Phase Hairs" International Journal of Molecular Sciences 10, no. 5: 1967-1977. https://doi.org/10.3390/ijms10051967