Localization of D-β-Aspartyl Residue-Containing Proteins in Various Tissues

Abstract
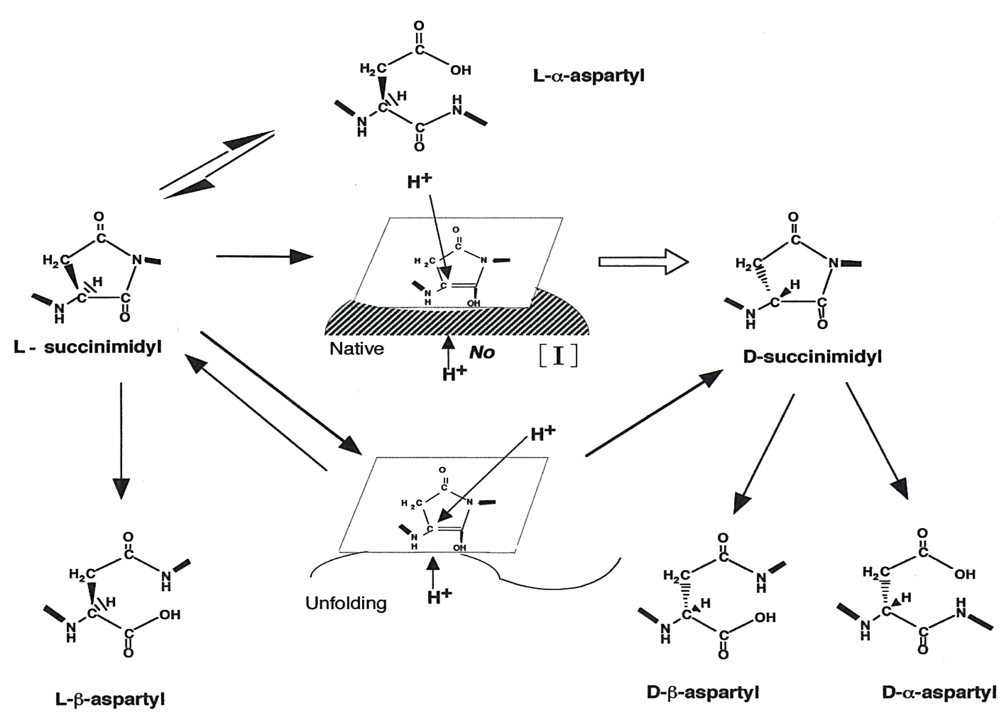
:1. Introduction
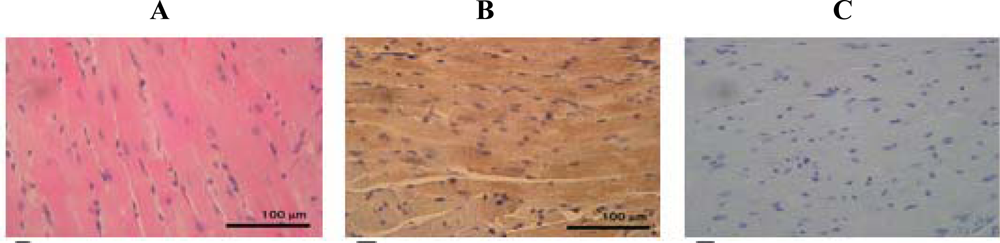
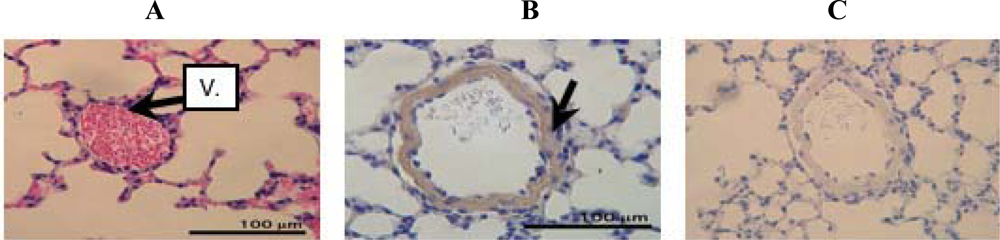
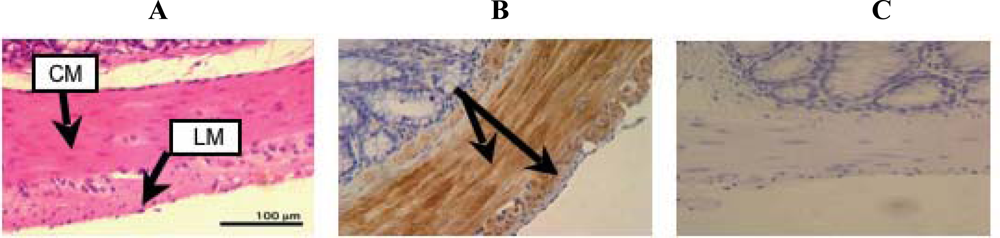
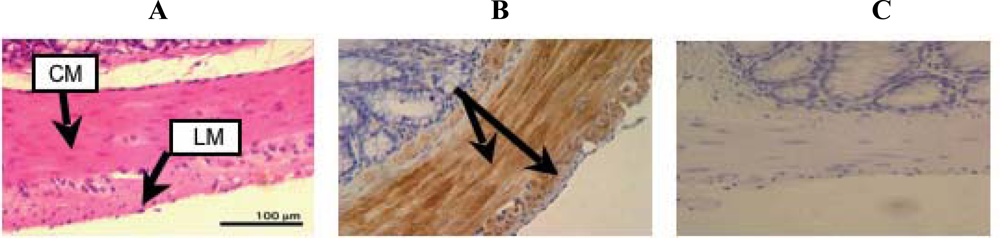
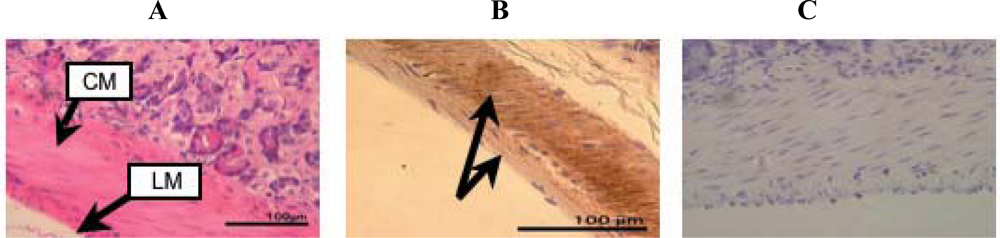
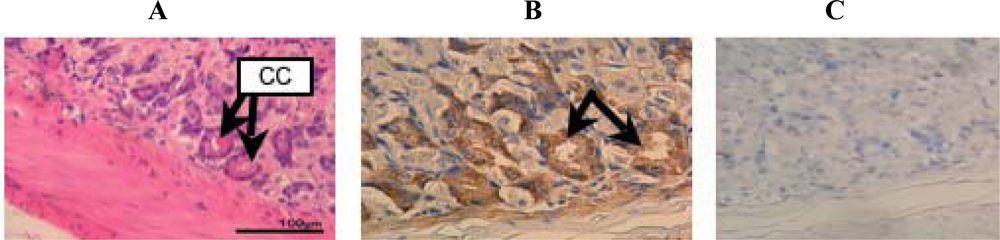
2. Results and Discussion
3. Experimental Section
3.1. Animals
3.2. Materials
3.3. Tissue preparation
3.4. Hematoxylin-Eosin staining
3.5. Antibody against d-β-Asp-containing peptide
3.6. Immunohistochemistry
4. Conclusions
Acknowledgments
References and Notes
- Masters, PM. Stereochemically altered noncollagenous protein from human dentin. Calcif. Tissue Int 1983, 35, 43–47. [Google Scholar]
- Ritz, S; Turzynski, A; Schutz, HW; Hollmann, A; Rochholz, G. Identification of osteocalcin as a permanent aging constituent of the bone matrix: basis for an accurate age at death determination. Forensic Sci. Int 1996, 77, 13–26. [Google Scholar]
- Powell, JT; Vine, N; Crossman, M. On the accumulation of D-aspartate in elastin and other proteins of the ageing aorta. Atherosclerosis 1992, 97, 201–208. [Google Scholar]
- Fisher, GH; Garcia, NM; Payan, IL; Cadilla-Perezrios, R; Sheremata, WAH; Man, EH. D-aspartic acid in purified myelin and myelin basic protein. Biochem. Biophys. Res. Commun 1986, 135, 683–687. [Google Scholar]
- Shapira, R; Chou, CH. Differential racemization of aspartate and serine in human myelin basic protein. Biochem. Biophys. Res. Commun 1987, 146, 1342–1349. [Google Scholar]
- Masters, PM; Bada, JL; Zigler, JS, Jr. Aspartic acid racemisation in the human lens during ageing and in cataract formation. Nature 1977, 268, 71–73. [Google Scholar]
- Fujii, N. D-amino acids in living higher organisms. Orig. Life Evol. Biosph 2002, 32, 103–127. [Google Scholar]
- Fujii, N; Satoh, K; Harada, K; Ishibashi, Y. Simultaneous stereoinversion and isomerization at specific aspartic acid residues in alpha A-crystallin from aged human lens. J. Biochem 1994, 116, 663–669. [Google Scholar]
- Fujii, N; Ishibashi, Y; Satoh, K; Fujino, M; Harada, K. Simultaneous racemization and isomerization at specific aspartic acid residues in alpha B-crystallin from the aged human lens. Biochim. Biophys. Acta 1994, 1204, 157–163. [Google Scholar]
- Roher, AE; Lowenson, JD; Clarke, S; Wolkow, C; Wang, R; Cotter, RJ; Reardon, IM; Zurcher-Neely, HA; Heinrikson, RL; Ball, MJ; Greenberg, BD. Structural alterations in the peptide backbone of beta-amyloid core protein may account for its deposition and stability in Alzheimer's disease. J. Biol. Chem 1993, 268, 3072–3083. [Google Scholar]
- Young, GW; Hoofring, SA; Mamula, MJ; Doyle, HA; Bunick, GJ; Hu, Y; Aswad, DW. Protein L-isoaspartyl methyltransferase catalyzes in vivo racemization of Aspartate-25 in mammalian histone H2B. J. Biol. Chem 2005, 280, 26094–26098. [Google Scholar]
- Fujii, N; Harada, K; Momose, Y; Ishii, N; Akaboshi, M. D-amino acid formation induced by a chiral field within a human lens protein during aging. Biochem. Biophys. Res. Commun 1999, 263, 322–326. [Google Scholar]
- Fujii, N; Takemoto, LJ; Momose, Y; Matsumoto, S; Hiroki, K; Akaboshi, M. Formation of four isomers at Asp-151 residue of aged human alpha A-crystallin by natural aging. Biochem. Biophys. Res. Commun 1999, 265, 746–751. [Google Scholar]
- Fujii, N; Momose, Y; Ishibashi, Y; Uemura, T; Takita, M; Takehana, M. Specific racemization and isomerization of the aspartyl residue of alpha A-crystallin due to UV-B irradiation. Exp. Eye Res 1997, 65, 99–104. [Google Scholar]
- Fujii, N; Shimo-Oka, T; Ogiso, M; Momose, Y; Kodama, T; Kodama, M; Akaboshi, M. Localization of biologically uncommon D-beta-aspartate-containing alphaA-crystallin in human eye lens. Mol. Vis 2000, 6, 1–5. [Google Scholar]
- Fujii, N; Tajima, S; Tanaka, N; Fujimoto, N; Takata, T; Shimo-Oka, T. The presence of D-beta-aspartic acid-containing peptides in elastic fibers of sun-damaged skin: a potent marker for ultraviolet-induced skin aging. Biochem. Biophys. Res. Commun 2002, 294, 1047–1051. [Google Scholar]
- Miura, Y; Fujimoto, N; Komatsu, T; Tajima, S; Kawada, A; Saito, T; Fujii, N. Immunohistochemical study of chronological and photo-induced aging skins using the antibody raised against -aspartyl residue-containing peptide. J. Cutan. Pathol 2004, 31, 51–56. [Google Scholar]
- Takata, T; Shimo-Oka, T; Miki, K; Fujii, N. Characterization of new D-beta-aspartate-containing proteins in a lens-derived cell line. Biochem. Biophys. Res. Commun 2005, 334, 1022–1031. [Google Scholar]
- Kaji, Y; Oshika, T; Takazawa, Y; Fukayama, M; Takata, T; Fujii, N. Localization of D-beta-aspartic acid containing proteins in human eyes. Invest. Ophthalmol. Vis. Sci 2007, 48, 3923–3927. [Google Scholar]
- McCudden, CR; Kraus, VB. Biochemistry of amino acid racemization and clinical application to musculoskeletal disease. Clin. Biochem 2006, 39, 1112–1130. [Google Scholar]
- Gineyts, E; Cloos, PA; Borel, O; Grimaud, L; Delmas, PD; Garnero, P. Racemization and isomerization of type I collagen C-telopeptides in human bone and soft tissues: assessment of tissue turnover. Biochem. J 2000, 345, 481–485. [Google Scholar]
- Geiger, T; Clarke, S. Deamidation, isomerization, and racemization at asparaginyl and aspartyl residues in peptides. Succinimide-linked reactions that contribute to protein degradation. J. Biol. Chem 1987, 262, 785–794. [Google Scholar]
- Stephenson, RC; Clarke, S. Succinimide formation from aspartyl and asparaginyl peptides as a model for the spontaneous degradation of proteins. J. Biol. Chem 1989, 264, 6164–6170. [Google Scholar]
- Tyler-Cross, R; Schirch, V. Effects of amino acid sequence, buffers, and ionic strength on the rate and mechanism of deamidation of asparagine residues in small peptides. J. Biol. Chem 1991, 266, 22549–22556. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | age | sex | brain | heart | lung | stomach | small intestine | long intestine | liver | kidney | spleen | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cardiac muscle | Vessel | Chief cell | Long. muscle | Circ. muscle | Long. muscle | Circ. muscle | Long. muscle | Circ. muscle | |||||||
| 1 | 6 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 2 | 6 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 3 | 6 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 4 | 24 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 5 | 24 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 6 | 54 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 7 | 54 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 8 | 92 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
| 9 | 92 | F | − | + | + | + | + | + | + | + | + | + | − | − | − |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Motoie, R.; Fujii, N.; Tsunoda, S.; Nagata, K.; Shimo-oka, T.; Kinouchi, T.; Fujii, N.; Saito, T.; Ono, K. Localization of D-β-Aspartyl Residue-Containing Proteins in Various Tissues. Int. J. Mol. Sci. 2009, 10, 1999-2009. https://doi.org/10.3390/ijms10051999
Motoie R, Fujii N, Tsunoda S, Nagata K, Shimo-oka T, Kinouchi T, Fujii N, Saito T, Ono K. Localization of D-β-Aspartyl Residue-Containing Proteins in Various Tissues. International Journal of Molecular Sciences. 2009; 10(5):1999-2009. https://doi.org/10.3390/ijms10051999
Chicago/Turabian StyleMotoie, Ryota, Noriko Fujii, Shigeru Tsunoda, Kenji Nagata, Tadashi Shimo-oka, Tadatoshi Kinouchi, Norihiko Fujii, Takeshi Saito, and Koji Ono. 2009. "Localization of D-β-Aspartyl Residue-Containing Proteins in Various Tissues" International Journal of Molecular Sciences 10, no. 5: 1999-2009. https://doi.org/10.3390/ijms10051999