Purification and Characterization of Organic Solvent and Detergent Tolerant Lipase from Thermotolerant Bacillus sp. RN2

Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Characterization of Bacillus sp. RN 2 Lipase
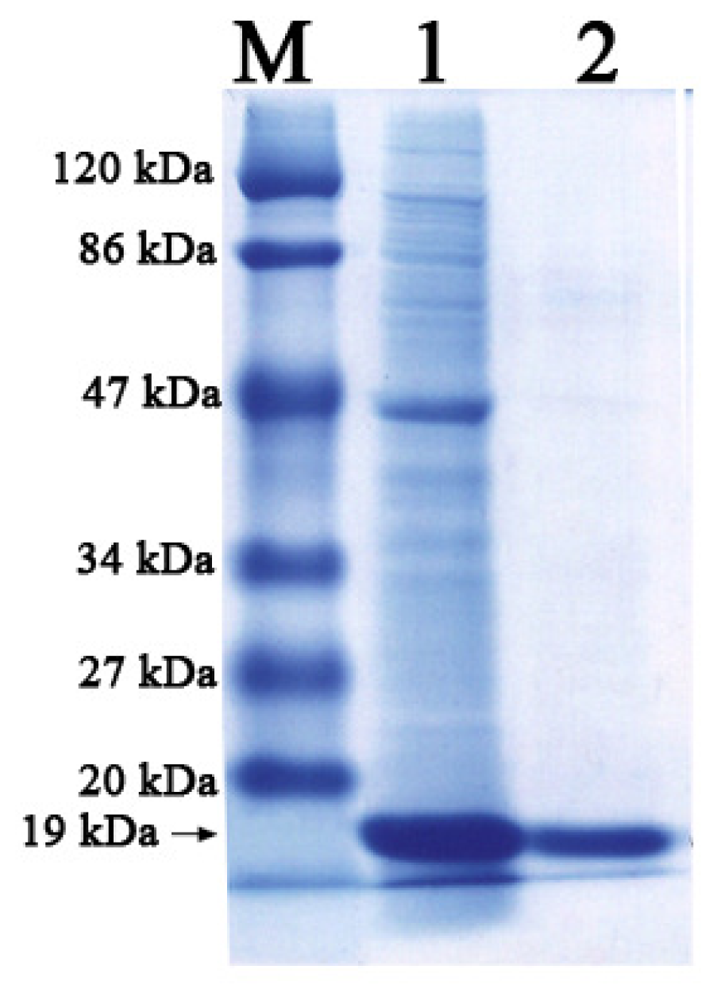
2.2. Expression and Purification of Lip-SBRN2
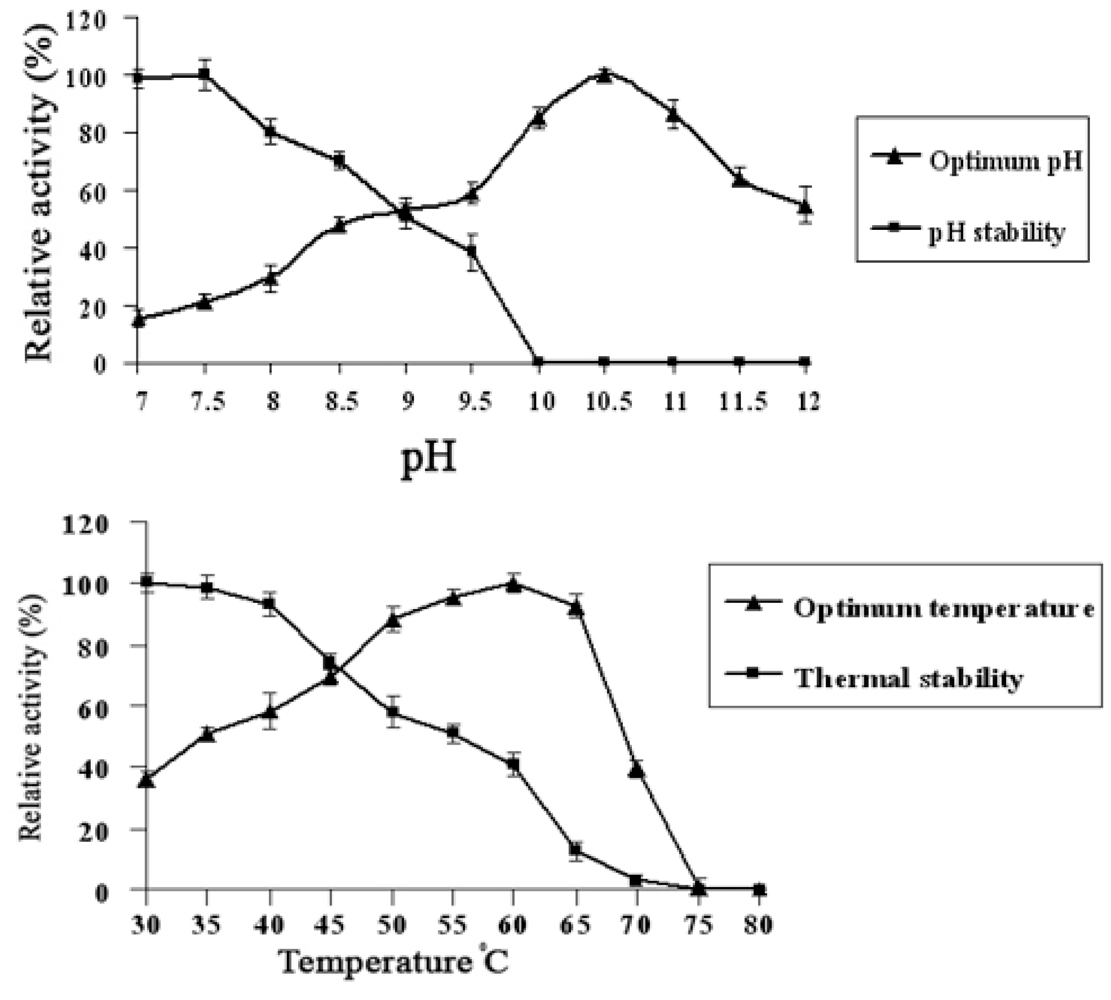
2.3. Optimum pH and Temperature for Lip-SBRN2 Activity
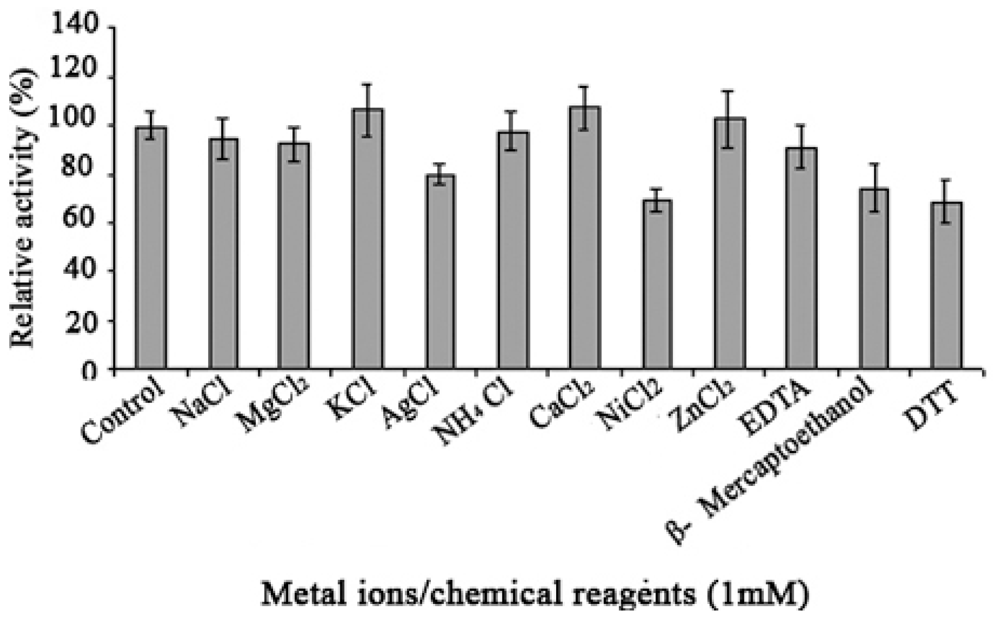
2.4. Effect of Metal Ions and Chemical Reagent on Lip-SBRN2 Activity
2.5. Substrate Specificity
2.6. Effect of Organic Solvents and Detergent on Lip-SBRN2 Activity
3. Experimental Section
3.1. Bacterial Strains Collection
3.2. PCR Amplification of Thermotolerant Bacillus sp. RN2 Lipase Gene (Lip-SBRN2 Gene)
3.3. Over-Expression of the Lip-SBRN2 Gene
3.4. Purification of Lip-SBRN2
3.5. Lipase Assay
4. Conclusion
Acknowledgements
References
- Cousin, X; Hotelier, T; Giles, K; Lievin, P; Toutant, JP; Chatonnet, A. The alpha/beta fold family of proteins database and the cholinesterase gene server esther. Nucleic Acids Res 1997, 25, 143–146. [Google Scholar]
- Jaeger, KE; Dijkstra, BW; Reetz, MT. Bacterial biocatalysts: Molecular biology, three-dimensional structures, and biotechnological applications of lipases. Annu. Rev. Microbiol 1999, 53, 315–351. [Google Scholar]
- Ollis, DL; Cheah, E; Cygler, M; Dijkstra, B; Frolow, F; Franken, SM; Harel, M; Remington, SJ; Silman, I; Schrag, J. The alpha/beta hydrolase fold. Protein Eng 1992, 5, 197–211. [Google Scholar]
- Chapus, C; Rovery, M; Sarda, L; Verger, R. Minireview on pancreatic lipase and colipase. Biochimie 1988, 70, 1223–1234. [Google Scholar]
- Wang, CS; Hartsuck, J. A Bile salt-activated lipase. A multiple function lipolytic enzyme. Biochim. Biophys. Acta 1993, 1166, 1–19. [Google Scholar]
- Schmidt-Dannert, C. Recombinant microbial lipases for biotechnological applications. Bioorg. Med. Chem 1999, 7, 2123–2130. [Google Scholar]
- Nthangeni, MB; Patterton, HG; Van, TA; Vergeer, WP; Litthauer, D. Over-expression and properties of a purified recombinant Bacillus licheniformis lipase: A comparative report on Bacillus lipases. Enzyme Microb. Technol 2001, 28, 705–712. [Google Scholar]
- Schmidt-Dannert, C; Rua, ML; Atomi, H; Schmid, RD. Thermoalkalophilic lipase of Bacillus thermocatenulatus. I. Molecular cloning, nucleotide sequence, purification and some properties. Biochim. Biophys. Acta 1996, 1303, 105–114. [Google Scholar]
- Lee, DW; Koh, YS; Kim, KJ; Kim, BC; Choi, HJ; Kim, DS; Suhartono, MT; Pyun, YR. Isolation and characterisation of a thermophilic lipase from Bacillus thermoleovorans ID-1. FEMS Microbiol. Lett 1999, 179, 393–400. [Google Scholar]
- Kim, HK; Park, SY; Lee, JK; Oh, TK. Gene cloning and characterization of thermostable lipase from Bacillus stearothermophilus L1. Biosci. Biotechnol. Biochem 1998, 62, 66–71. [Google Scholar]
- Zock, J; Cantwell, C; Swartling, J; Hodges, R; Pohl, T; Sutton, K; Rosteck, P; McGilvray, D; Queener, S. The Bacillus subtilis pnbA gene encoding p-nitrobenzyl esterase: Cloning, sequence and high-level expression in Escherichia coli. Gene 1994, 151, 37–43. [Google Scholar]
- Grangemard, I; Wallach, J; Maget-Dana, R; Peypoux, F. Lichenysin: A more efficient cation chelator than surfactin. Appl. Biochem. Biotechnol 2001, 90, 199–210. [Google Scholar]
- Jenny, K; Kappeli, O; Fiechter, A. Biosurfactants from Bacillus licheniformis: Structural analysis and characterization. Appl. Microbiol. Biotechnol 1991, 36, 5–13. [Google Scholar]
- Yakimov, MM; Timmis, KN; Wray, V; Fredrickson, HL. Characterization of a new lipopeptide surfactant produced by thermotolerant and halotolerant subsurface Bacillus licheniformis BAS50. Appl. Environ. Microbiol 1995, 61, 1706–1713. [Google Scholar]
- Pohlenz, HD; Boidol, W; Schuttke, I; Streber, W. Purification and properties of an Arthrobacter oxydans P52 carbamate hydrolase specific for the herbicide phenmedipham and nucleotide sequence of the corresponding gene. J. Bacteriol 1992, 174, 6600–6607. [Google Scholar]
- Lesuisse, E; Schanck, K; Colson, C. Purification and preliminary characterization of the extracellular lipase of Bacillus subtilis 168, an extremely basic pH-tolerant enzyme. Eur. J. Biochem 1993, 216, 155–160. [Google Scholar]
- Sugihara, A; Ueshima, M; Shimada, Y; Tsunasawa, S; Tominaga, Y. Purification and characterization of a novel thermostable lipase from Pseudomonas cepacia. J. Biochem. (Tokyo) 1992, 112, 598–603. [Google Scholar]
- Iacazio, G; Perissol, C; Faure, B. A new tannase substrate for spectrophotometric assay. J. Microbiol. Methods 2000, 42, 209–214. [Google Scholar]
- Kwon, DY; Rhee, JS. A Simple and Rapid colorimetric method for determination of free fatty acids for lipase assay. J. Am. Oil. Chem. Soc 1986, 63, 89–92. [Google Scholar]
- Pencreac’h, G; Baratti, JC. Hydrolysis of p-nitrophenyl palmitate in n-heptane by the Pseudomonas cepacia lipase: A simple test for the determination of lipase activity in organic media. Enzyme Microb. Technol 1996, 18, 417–422. [Google Scholar]
- Goto, K; Omura, T; Hara, Y; Sadaie, Y. Application of the partial 16S rDNA sequence as an index for rapid identification of species in the gen Bacillus. J. Gen. Appl. Microbiol 2000, 46, 1–8. [Google Scholar]
- Bradford, MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Laemmli, UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Steps | Volume (mL) | Total activity (Units) | Total activity/mL (Units/mL) | Total protein (mg) | Specific activity (Units/mg) | Purification factor (fold) | Recovery yield (%) |
|---|---|---|---|---|---|---|---|
| Crude enzyme | 5 | 281 | 56.2 | 174 | 1.6 | 1 | 100 |
| Ni-NTA column | 2 | 144 | 72 | 1.3 | 118 | 74 | 51 |
| Substrates | Relative activity (%) |
|---|---|
| p-nitrophenylester | |
| pNP-butyrate (C4) | 40 ± 6 |
| pNP-caprylate (C8) | 92 ± 8 |
| pNP-caprate (C10) | 68 ± 9 |
| pNP-laurate (C12) | 100 ± 15 |
| pNP-myristate (C14) | 59 ± 7 |
| pNP-palmitate (C16) | 54 ± 10 |
| pNP-stearate (C18) | 35 ± 4 |
| Natural oils | |
| Coconut oil (12:0) | 727.3 ± 16 |
| Palm oil (16:0) | 32.0 ± 13 |
| Olive oil (18:1) | 100.0 ± 9 |
| Grape seed oil (18:2) | 32.4 ± 7 |
| Soybean oil (18:3) | 42.2 ± 7 |
| Organic solvents/detergents (25%) | Relative activity of Bacillus sp. RN2 lipase | |
|---|---|---|
| 30 min incubation Relative activity (%) | 2 h incubation Relative activity (%) | |
| None | 100.0 ± 7 | 100.0 ± 4 |
| Organic solvents | ||
| Benzene | 108.0 ± 11 | 104.4 ± 3.4 |
| Diethylether | 102.4 ± 5.2 | 103.6 ± 0.5 |
| Acetone | 100.8 ± 0.8 | 82.2 ± 11.3 |
| n-hexane | 94.0 ± 4 | 94.5 ± 3.5 |
| Dimethylsulfoxide | 85.2 ±.8.4 | 96.4 ± 11.3 |
| Cyclohexane | 84.4 ± 11.2 | 85.1 ± 5.6 |
| N,N-dimethylformamide | 80.4 ± 5.5 | 86.9 ± 3.4 |
| Chloroform | 80.0 ± 11 | 84.4 ± 10.5 |
| Acetonitrile | 69.6 ± 9.5 | 82.5 ± 11 |
| Toluene | 68.4 ± 12.3 | 80.4 ± 13.4 |
| 2-propanol | 60.8 ± 8.7 | 66.5 ± 6.6 |
| Detergents | ||
| 25%Tween-20 | 60.8 ± 11.5 | 96.4 ± 7.3 |
| 25%Triton X-100 | 41.6 ± 10.6 | 86.5 ± 8.7 |
| 1% SDS | 112.5 ± 11.3 | 485.7 ± 11.5 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kanjanavas, P.; Khuchareontaworn, S.; Khawsak, P.; Pakpitcharoen, A.; Pothivejkul, K.; Santiwatanakul, S.; Matsui, K.; Kajiwara, T.; Chansiri, K. Purification and Characterization of Organic Solvent and Detergent Tolerant Lipase from Thermotolerant Bacillus sp. RN2. Int. J. Mol. Sci. 2010, 11, 3783-3792. https://doi.org/10.3390/ijms11103783
Kanjanavas P, Khuchareontaworn S, Khawsak P, Pakpitcharoen A, Pothivejkul K, Santiwatanakul S, Matsui K, Kajiwara T, Chansiri K. Purification and Characterization of Organic Solvent and Detergent Tolerant Lipase from Thermotolerant Bacillus sp. RN2. International Journal of Molecular Sciences. 2010; 11(10):3783-3792. https://doi.org/10.3390/ijms11103783
Chicago/Turabian StyleKanjanavas, Pornpimon, Sintawee Khuchareontaworn, Paisarn Khawsak, Arda Pakpitcharoen, Khajeenart Pothivejkul, Somchai Santiwatanakul, Kenji Matsui, Tadahiko Kajiwara, and Kosum Chansiri. 2010. "Purification and Characterization of Organic Solvent and Detergent Tolerant Lipase from Thermotolerant Bacillus sp. RN2" International Journal of Molecular Sciences 11, no. 10: 3783-3792. https://doi.org/10.3390/ijms11103783