Effect of Different Light Intensities on Total Phenolics and Flavonoids Synthesis and Anti-oxidant Activities in Young Ginger Varieties (Zingiber officinale Roscoe)

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolics (TP) Content
2.2. Total Flavonoids (TF) Content
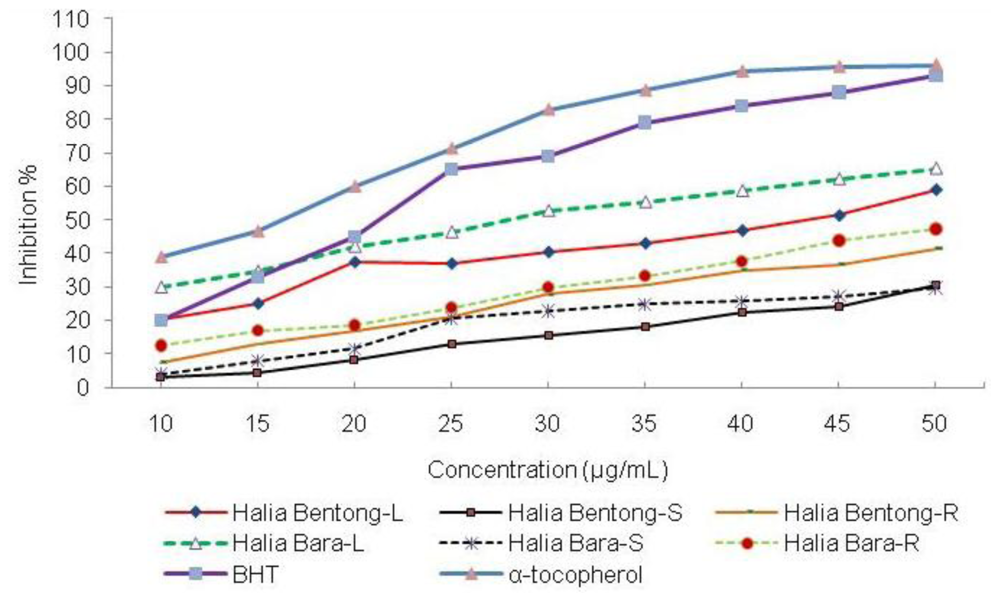
2.3. Antioxidant Activity
2.3.1. 1,1-Diphenyl-2-picryl-hydrazyl (DPPH) Assay
2.3.2. Reducing Ability, Ferric Reducing Antioxidant Potential (FRAP)
3. Experimental Section
3.1. Plant Material and Maintenance
3.2. Extract Preparation
3.3. Determination of Total Phenolic Contents (TP)
3.4. Determination of Total Flavonoid Contents (TF)
3.5. Determination of Antioxidant Activities
3.5.1. DPPH Radical Scavenging Assay
3.5.2. Reducing Ability (FRAP Assay)
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Larsen, K; Ibrahim, H; Khaw, SH; Saw, LG. Gingers of Peninsular Malaysia and Singapore; Natural History Publications (Borneo): Kota Kinabalu, Sabah, Malaysia, 1999. [Google Scholar]
- Kikuzaki, H; Nakatani, N. Antioxidant effect of some ginger constituents. Food Sci 1993, 578, 1407–1410. [Google Scholar]
- Kim, D-O; Jeond, SW; Lee, CY. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem 2003, 81, 321–326. [Google Scholar]
- Atoui, K; Mansouri, A; Bosku, G; Kefalas, P. Tea and herbal infusions: their antioxidant activity and phenolic profile. Food Chem 2005, 89, 27–36. [Google Scholar]
- Cook, NC; Samman, S. Review: flavonoids chemistry, methabolysm, cardioprotective effects and dietary sources. Nutr. Biochem 1996, 7, 66–76. [Google Scholar]
- Arts, IC; Jacobs, DRJ; Gross, M; Harnack, LJ; Folsom, AR. Dietary Catechins and cancer incidence among postmenopausal women: the Iowa Women’s Health Study (United States). Cancer Causes Control 2002, 13, 373–382. [Google Scholar]
- Davis, W; Lamson, MS; Matthew, S; Brignall, ND. Antioxidants and Cancer III: Quercetin. Altern. Med. Rev 2000, 5, 196–208. [Google Scholar]
- Shukla, Y; Prasad, S; Tripathi, C; Singh, M; George, J; Kalra, N. In vitro and in vivo modulation of testosterone mediated alterations in apoptosis related proteins by [6]-gingerol. Mol. Nutr. Food Res 2007, 51, 1492–1502. [Google Scholar]
- Feng, XP; Cha, ZY; Wang, HS. Study on the Extractive Technology of Total Flavones from Foliage of Gingko. Chem. Ind. Corros. Control Sichuan 2002, 2, 138. [Google Scholar]
- Liao, HB; He, ZE. Research and Prospect of kudzu root. Nat. Food Assoc 2003, 24, 81–83. [Google Scholar]
- Chen, HG; Yu, YG; Zeng, OX. Study on extraction of flavonoids and Alkaloids from lotus leaf. Food Sci 2002, 23, 69–71. [Google Scholar]
- Ghasemzadeh, A; Jaafar, HZE; Rahmat, A. Antioxidant Activities, Total Phenolics and Flavonoids Content in Two Varieties of Malaysia Young Ginger (Zingiber officinale Roscoe). Molecules 2010, 15, 4324–4333. [Google Scholar]
- Rozanida, AR; Nurul Izza, N; Mohd Helme, MH; Zanariah, H. Xanwhite TM–A cosmeceutical product from species in the family Zingiberaceae. Forest Res. Inst. Malaysia Selangor 2005, 14, 31–36. [Google Scholar]
- Tang, Y; Zhao, A. Study on the Flavone Content of Buckwheat. J. Sichuan Agric. Univ 2001, 19, 352–354. [Google Scholar]
- Hemm, MR; Rider, SD; Ogas, J; Murry, DJ; Chapple, C. Light induces phenylpropanoid metabolism in Arabidopsis roots. Plant J 2004, 38, 765–778. [Google Scholar]
- Liu, CZ; Guo, C; Wang, YC; Ouyang, F. Effect of light irradiation on hairy root growth and artemisinin biosynthesis of Artemisia annua. Process Biochem 2002, 38, 581–585. [Google Scholar]
- Xie, BD; Wang, HT. Effects of light spectrum and photoperiod on contents of flavonoid and terpene in leaves of Ginkgo biloba L. Nanjing Forestry Univ 2006, 30, 51–54. [Google Scholar]
- Graham, TL. Flavonoid and flavonol glycoside metabolism in Arabidopsis. Plant Physiol. Biochem 1998, 36, 135–144. [Google Scholar]
- Briskin, DP; Gawienowski, MC. Differential effects of light and nitrogen on production of hypericins and leaf glands in Hypericum perforatum. Plant Physiol 2001, 39, 1075–1081. [Google Scholar]
- Kurata, H; Matsumura, S; Furusaki, S. Light irradiation causes physiological andmetabolic changes for purine alkaloid production by a Coffea Arabica cell suspension culture. Plant Sci 1997, 123, 197–203. [Google Scholar]
- Ferreira, ICFR; Baptista, P; Vilas-Boas, M; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: individual cap and stipe activity. Food Chem 2007, 100, 1511–1516. [Google Scholar]
- Rodrigo, R; Bosco, C. Oxidative stress and protective effect of polyphenols: comparative studies in human and rodent kidney. Comp. Biochem. Physiol 2006, 142, 317–327. [Google Scholar]
- Mori, K; Goto-Yamamoto, N; Kitayama, M; Hashizume, K. Loss of anthocyanins in red-wine grape under high temperatura. Exp. Bot 2007, 58, 1935–1945. [Google Scholar]
- Mori, K; Sugaya, S; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic 2005, 105, 319–330. [Google Scholar]
- Yamane, T; Jeong, ST; Goto-Yamamoto, N; Koshita, Y; Kobayashi, S. Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Viticult 2006, 57, 54–59. [Google Scholar]
- Katsube, T; Tabata, H; Ohta, Y; Yamasaki, Y; Anuurad, E; Shiwaku, K. Screening for antioxidant activity in edible plant products: Comparison of low-density lipoprotein oxidation assay, DPPH radical scavenging assay and Folin–Ciocalteu assay. Agric. Food Chem 2004, 52, 2391–2396. [Google Scholar]
- Chan, EWC; Lim, YY; Wong, FL; Lianto, FS. Antioxidant and tyrosinase inhibition properties of leaves and rhizomes in ginger species. Food Chem 2008, 109, 477–483. [Google Scholar]
- Ravindran, PN; Nirmal Babu, K. Ginger the genus Zingiber; C.H.I.P.S: Weimar, Texas, TX, USA, 2005. [Google Scholar]
- Diaz, J; Bernal, A; Pomar, F; Merino, F. Induction of shikimate dehydrogenase and peroxidase in pepper (Capsicumannum L.) seedlings in response to copper stress and its relation to lignification. Plant Sci 2001, 161, 161–179. [Google Scholar]
- Sergio, LC; Claudio, FP; Carlos, SA. Inhibitory effects of shikmic acid on pep carboxylase activity. Plant Cell Physiol 1996, 37, 870–872. [Google Scholar]
- Vinay, RP; Prakash, RP; Sushil, SK. Antioxidant Activity of Some Selected Medicinal Plants in Western Region of India. Adv. Biol. Res 2010, 4, 23–26. [Google Scholar]
- Fonseca, JM; Rushing, JW; Rajapakse, NC. Potential implications of medicinal plants production in controlled environments: the case of feverfew (Tanacetum parthenium). Horticult. Sci 2006, 41, 531–535. [Google Scholar]
- Mosaleeyanon, K; Zobayed, SMA; Afreen, F. Relationship between net photosynthesis rate and secondary metabolite content in Strawberry. Plant Sci 2005, 169, 523–553. [Google Scholar]
- Michel, H; Klaus, K. The protective functions of carotenoids and flavonoids pigments against excess visible radiation at chilling temperature investigated in Arabidopsis. Planta 2001, 213, 953–966. [Google Scholar]
- Bergquist, S; Gertsson, U; Nordmark, LY; Olsson, ME. Effects of shade nettings, sowing time and storage on baby spinach flavonoids. J. Sci. Food Agric 2007, 87, 2464–2471. [Google Scholar]
- Ramawat, KG (Ed.) Herbal Drugs: Ethnomedicine to Modern Medicine; Springer-Verlag: Berlin, Heidelberg, Germany, 2009; Volume 22, p. 32.
- Miean, KH; Mohamed, S. Flavonoid (Myricetin, Quercetin, Kaempferol, Luteolin, and Apigenin) Content of Edible Tropical Plants. J. Agric. Food Chem 2001, 49, 3106–3112. [Google Scholar]
- Shui-Yuan, C; Feng, X; Yan, W. Advances in the study of flavonoids in Ginkgo biloba leaves. J. Med. Plants Res 2009, 3, 1248–1252. [Google Scholar]
- Ruiz, JM; Rivero, RM; Lopez-Cantarero, I; Romero, L. Role of Ca2+ in metabolism of phenolic compounds in tobacco leaves (Nicotiana tabacum L.). Plant Growth Regul 2003, 41, 173–177. [Google Scholar]
- Sakihama, Y; Yamasaki, H. Lipid peroxidation induces by phenolics in cinjunction with aluminium ions. J. Biol. Plant 2002, 45, 249–254. [Google Scholar]
- Larson, RA. The antioxidants of higher plants. Phytochemistry 1988, 27, 969–978. [Google Scholar]
- Miliauskas, G; Venskutonis, PR; van Beek, TA. Screening of radical scavenging activity of some medicinal and aromatic plant extracts. Food Chem 2004, 85, 231–237. [Google Scholar]
- Agnaniet, H; Menut, C; Bessie, JM. Aromatic plants of tropical central Africa. Part XLIX: Chemical composition of essential oils of the leaf and rhizomes of Aframomum giganteum from Gabon. Flavour Fragrance 2004, 19, 205–209. [Google Scholar]
- Elzaawely, AA; Xuan, TD; Tawata, S. Essential oils, kava pyrones and phenolic compounds from leaves and rhizomess of Alpinia zerumbet and their antioxidant activity. Food Chem 2007, 103, 486–494. [Google Scholar]
- Mpalantinos, MA; de Moura, RS; Parente, JP; Kuster, RM. Biologically active flavonoids and kava pyrones from the aqueous extract of Alpinia zerumbet. Phytother. Res 1998, 12, 442–444. [Google Scholar]
- Habsah, M; Amran, M; Mackeen, MM; Lajis, NH. Screening of Zingiberaceae extracts for antimicrobial and antioxidant activities. J. Ethnopharmacol 2000, 72, 403–410. [Google Scholar]
- Hasna, O; Afidah, A. Antioxidant activity and phenolic content of Paederia foetida and Syzygium aqueum. Molecules 2009, 14, 970–978. [Google Scholar]
- Ghasemzadeh, A; Jaafar, HZE; Rahmat, A. Identification and Concentration of Some Flavonoid Components in Malaysian Young Ginger (Zingiber officinale Roscoe) Varieties by a High Performance Liquid Chromatography Method. Molecules 2010, 15, 6231–6243. [Google Scholar]
- Oktay, M; Gulcin, I; Kufrevioglu, OI. Determination of in vitro antioxidant activity of fennel (Foeniculum vulgare) seed extracts. Lebensm. Wiss. Techol 2003, 36, 263–271. [Google Scholar]
- Bushra, S; Farooq, A; Muhammad, A. Effect of Extraction Solvent/Technique on the Antioxidant Activity of Selected Medicinal Plant Extracts. Molecules 2009, 14, 2167–2180. [Google Scholar]
- Mensor, LL; Menezes, FS; Leitao, GG; Reis, AS; dos Santos, TS; Coube, CS; Leitão, SG. Screening of Brazilian plant extracts for antioxidant activity by the use of DPPH free radical method. Phytother. Res 2001, 15, 127–130. [Google Scholar]
- Benzie, IFF; Strain, JJ. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: the FRAP assay. Anal. Biochem 1996, 239, 70–76. [Google Scholar]


{kind=link}
| Light Intensities (μmol m−2s−1) | Plant Part | TF (mg Quercetin/g dry weight) | TP (mg Gallic acid/g dry weight) | ||
|---|---|---|---|---|---|
| Halia Bentong | Halia Bara | Halia Bentong | Halia Bara | ||
| 310 | Leaves | 5.95 + 0.2c | 8.45 ± 0.38a | 27.43 ± 2.34e | 31.73 ± 2.10cd |
| Stems | 1.83 + 0.22hi | 1.96 ± 0.28h | 6.38 ± 1.25h | 7.11 ± 1.58gh | |
| Rhizomes | 3.91 + 0.083efg | 4.34 ± 0.08e | 8.9 ± 0.23fgh | 9.48 ± 0.21fgh | |
| 460 | Leaves | 5.04 + 0.27d | 5.7 ± 0.09cd | 28.96 ± 1.55de | 34.16 ± 4.8bc |
| Stems | 1.27 + 0.2i | 1.47 ± 0.21hi | 7.33 ± 1.13fgh | 8.432 ± 1.19fgh | |
| Rhizomes | 3.47 + 0.14fg | 4.03 ± 0.061efg | 9.69 ± 0.38fgh | 11.22 ± 0.16fg | |
| 630 | Leaves | 4.14 + 0.18ef | 6.12 ± 0.015c | 31.1 ± 0.98cde | 37.33 ± 4.45ab |
| Stems | 1.3 + 0.24hi | 1.55 ± 0.33hi | 7.47 ± 1.37fgh | 8.83 ± 1.82fgh | |
| Rhizomes | 3.37 + 0.079g | 3.97 ± 0.28efg | 9.81 ± 0.21fgh | 11.05 ± 0.77fg | |
| 790* | Leaves | 5.71 + 0.54cd | 7.05 ± 1.67b | 33 ± 1.13cd | 39.06 ± 9.23a |
| Stems | 1.26 + 0.12hi | 1.5 ± 0.14hi | 7.8 ± 0.68fgh | 8.56 ± 0.81fgh | |
| Rhizomes | 3.66 + 0.125fg | 4.14 ± 0.13ef | 10.22 ± 0.33fgh | 11.53 ± 0.36f | |
| Light Intensities (μmol m−2s−1) | Extraction Source | Halia Bentong | Halia Bara |
|---|---|---|---|
| 310 | Leaves | 59.02 ± 0.87b | 65.26 ± 0.9a |
| Stems | 30.31 ± 1.84hi | 29.59 ± 0.59i | |
| Rhizomes | 41.36 ± 0.63f | 47.26 ± 0.92e | |
| 790* | Leaves | 51.12 ± 1.65d | 56.36 ± 0.97c |
| Stems | 32.85 ± 0.57g | 31.45 ± 1.49gh | |
| Rhizomes | 51.41 ± 0.51d | 58.22 ± 1.19b | |
| Light Intensities (μmol m−2s−1) | Extraction Source | Halia Bentong | Halia Bara |
|---|---|---|---|
| 310 | Leaves | 552.24 ± 32.4f | 587.31 ± 25.6e |
| Stems | 378.4 ± 48.2h | 372.33 ± 32.33hi | |
| Rhizomes | 692.71 ± 16.48c | 788.57 ± 22.6a | |
| 790 | Leaves | 541.55 ± 34.1g | 574.9 ± 58.14e |
| Stems | 381.11 ± 48.7h | 363.1 ± 21.43i | |
| Rhizomes | 677.2 ± 18.38d | 770.4 ± 43.11b | |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ghasemzadeh, A.; Jaafar, H.Z.E.; Rahmat, A.; Wahab, P.E.M.; Halim, M.R.A. Effect of Different Light Intensities on Total Phenolics and Flavonoids Synthesis and Anti-oxidant Activities in Young Ginger Varieties (Zingiber officinale Roscoe). Int. J. Mol. Sci. 2010, 11, 3885-3897. https://doi.org/10.3390/ijms11103885
Ghasemzadeh A, Jaafar HZE, Rahmat A, Wahab PEM, Halim MRA. Effect of Different Light Intensities on Total Phenolics and Flavonoids Synthesis and Anti-oxidant Activities in Young Ginger Varieties (Zingiber officinale Roscoe). International Journal of Molecular Sciences. 2010; 11(10):3885-3897. https://doi.org/10.3390/ijms11103885
Chicago/Turabian StyleGhasemzadeh, Ali, Hawa Z. E. Jaafar, Asmah Rahmat, Puteri Edaroyati Megat Wahab, and Mohd Ridzwan Abd Halim. 2010. "Effect of Different Light Intensities on Total Phenolics and Flavonoids Synthesis and Anti-oxidant Activities in Young Ginger Varieties (Zingiber officinale Roscoe)" International Journal of Molecular Sciences 11, no. 10: 3885-3897. https://doi.org/10.3390/ijms11103885