Anti-Photoaging Effects of Soy Isoflavone Extract (Aglycone and Acetylglucoside Form) from Soybean Cake


Abstract
:1. Introduction
2. Results and Discussion
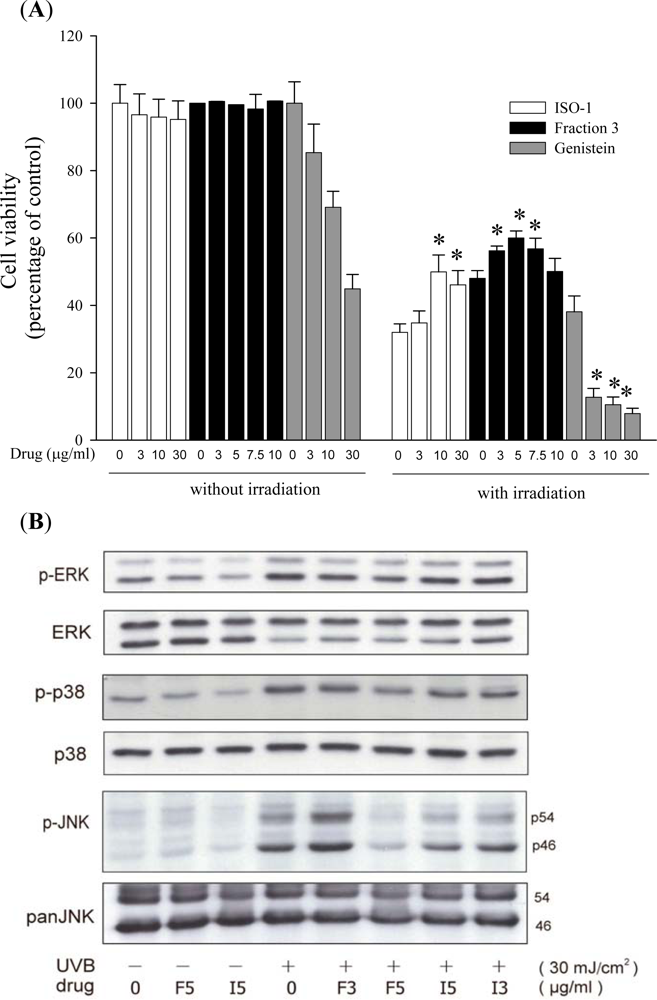
2.1. Soy Isoflavone Extract Fraction 3 Inhibits UVB-Induced Keratinocyte Death
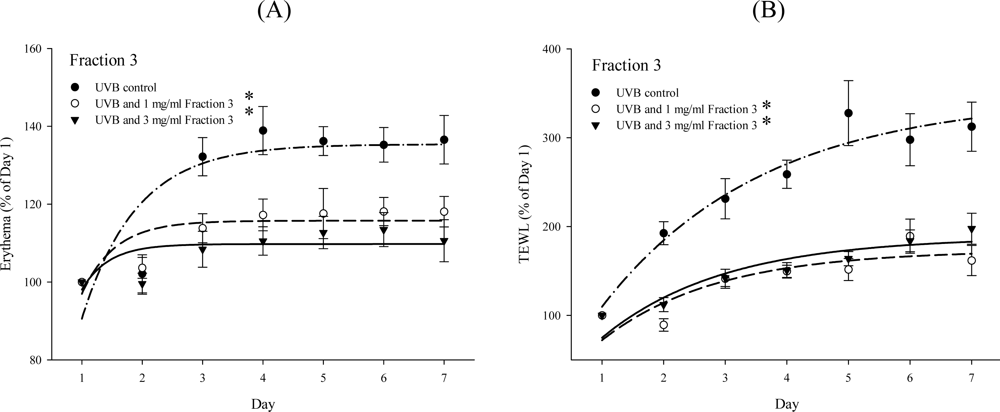
2.2. Soy Isoflavone Extract Fraction 3 Attenuates the Level of Erythema and TEWL after UVB Exposure
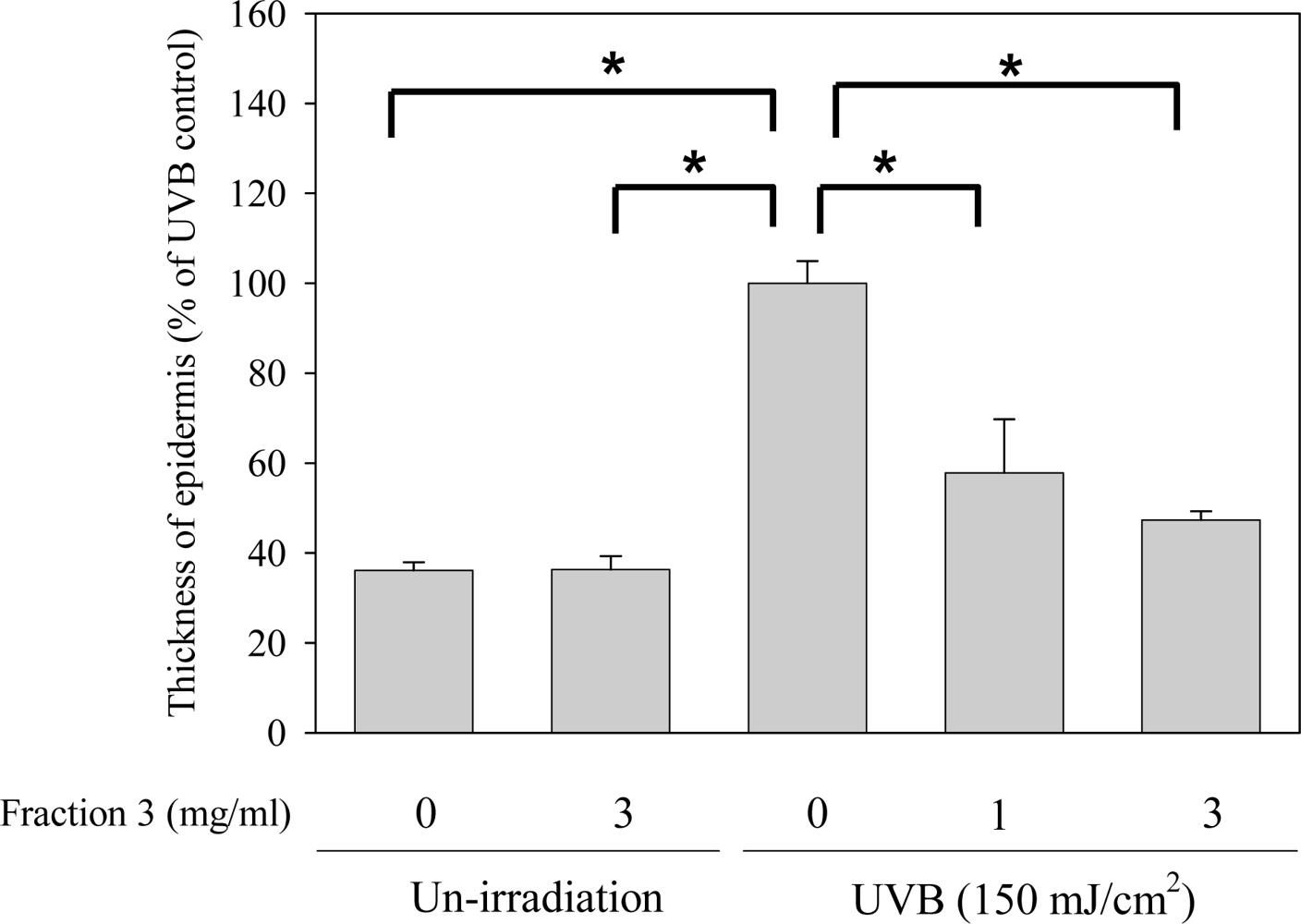
2.3. Soy Isoflavone Extract Fraction 3 Reduces the Epidermal Thickness
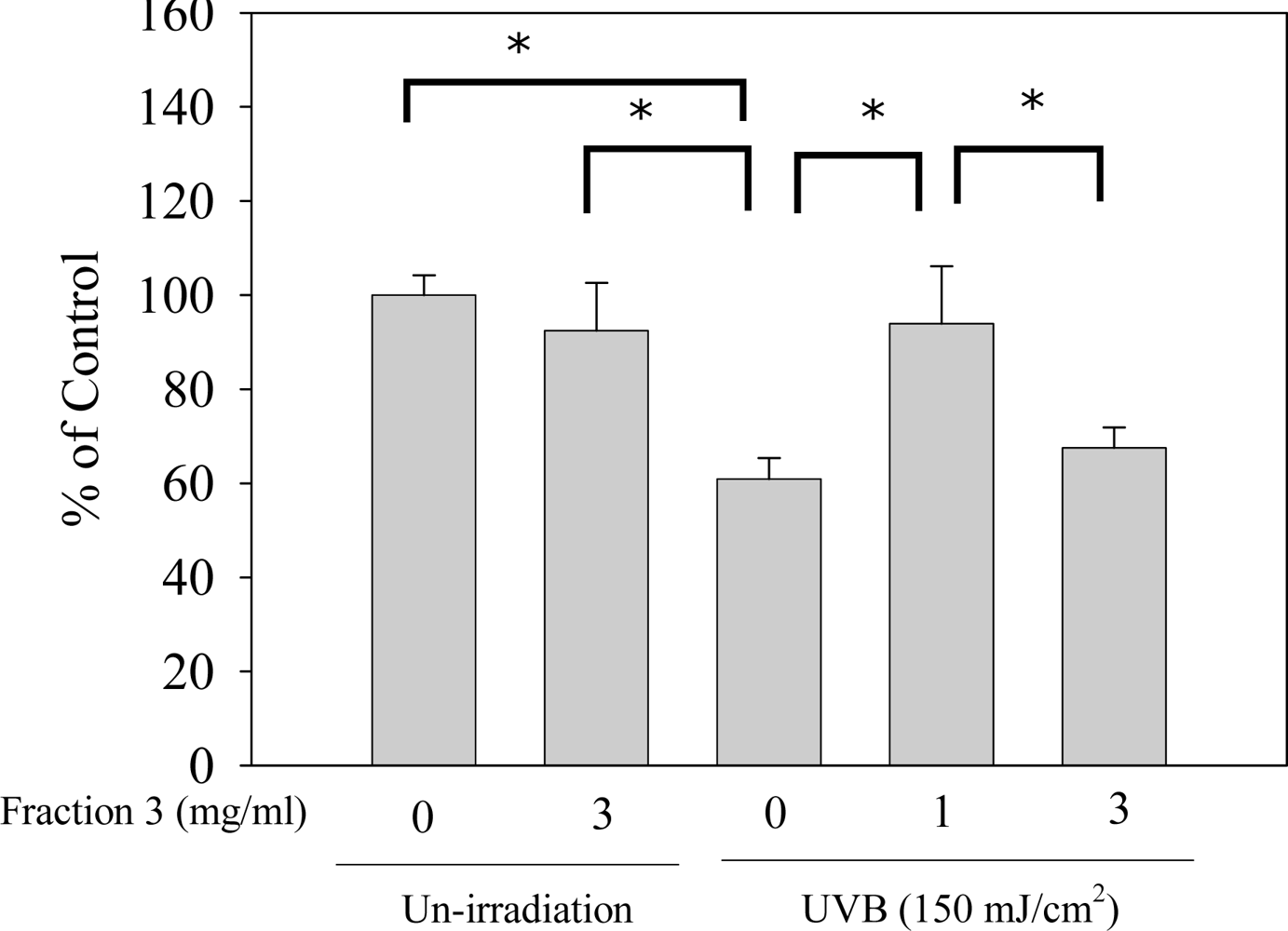
2.4. Soy Isoflavone Extract Fraction 3 Decreases the Depletion of Catalase after UVB Exposure
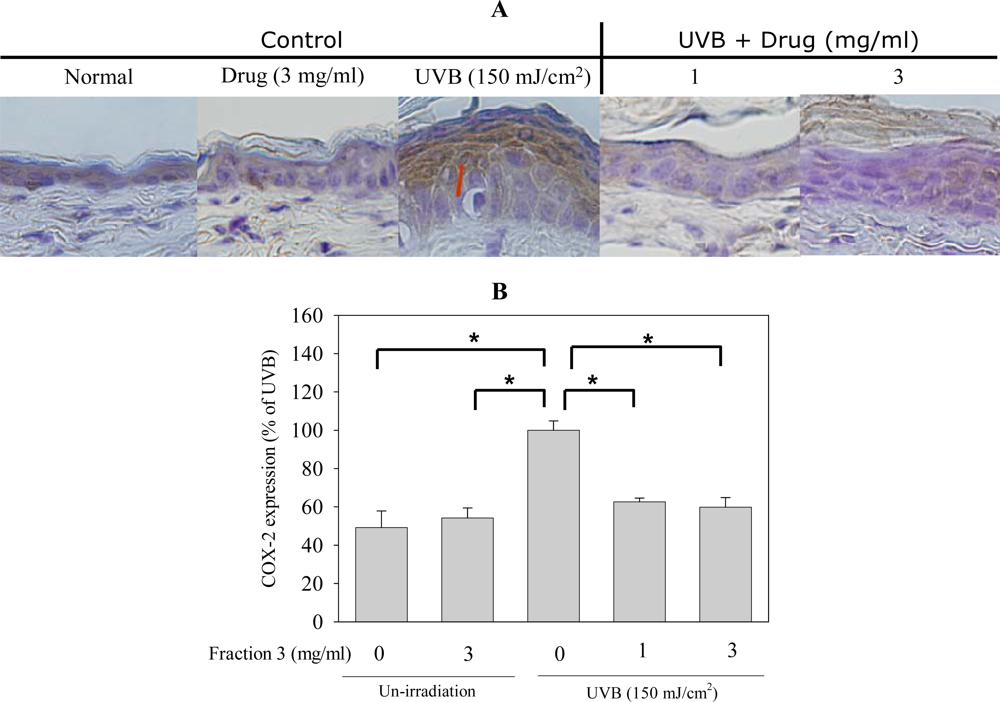
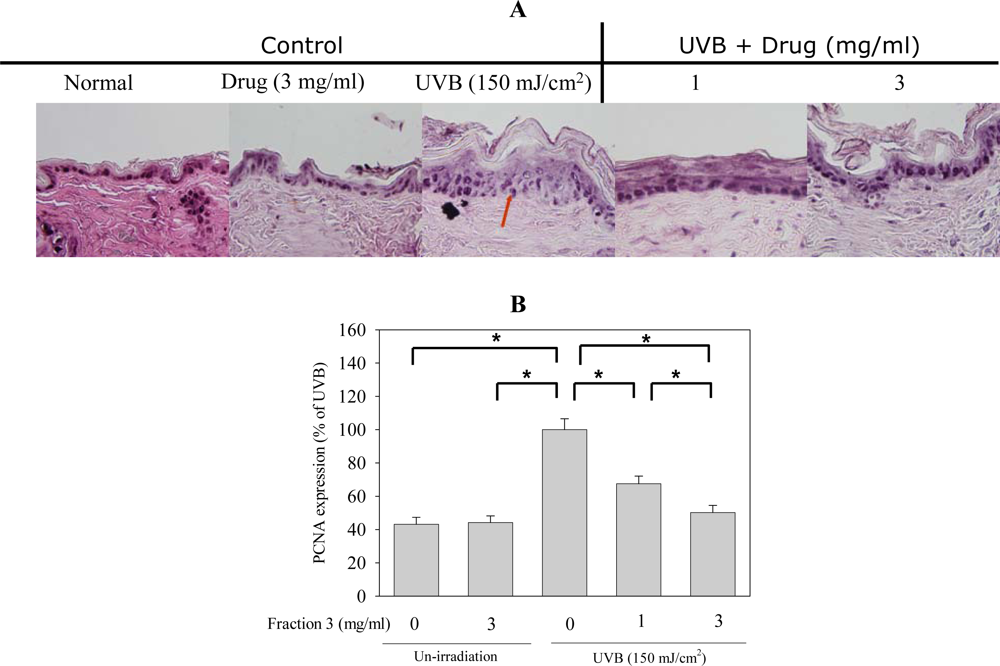
2.5. Soy Isoflavone Extract Fraction 3 Suppresses the Expression of COX-2 and PCNA after UVB Exposure
3. Experimental Section
3.1. Chemicals
3.2. Extraction of Soybean Isoflavones
3.3. Cell Viability Assay (MTT Assay)
3.4. Western Blotting
3.5. Animals and Measurement of Physiological Skin Functions
3.6. Histology and Immunostaining
3.7. Catalase Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Afaq, F; Mukhtar, H. Botanical antioxidants in the prevention of photocarcinogenesis and photoaging. Exp. Dermatol 2006, 15, 678–684. [Google Scholar]
- de Gruijl, FR; Forbes, PD. UV-induced skin cancer in a hairless mouse model. Bioessays 1995, 17, 651–660. [Google Scholar]
- Godar, DE. UV doses worldwide. Photochem. Photobiol 2005, 81, 736–749. [Google Scholar]
- Podda, M; Traber, MG; Weber, C; Yan, LJ; Packer, L. UV-irradiation depletes antioxidants and causes oxidative damage in a model of human skin. Free Radic. Biol. Med 1998, 24, 55–65. [Google Scholar]
- Chiang, HS; Wu, WB; Fang, JY; Chen, BH; Kao, TH; Chen, YT; Huang, CC; Hung, CF. UVB-protective effects of isoflavone extracts from soybean cake in human keratinocytes. Int. J. Mol. Sci 2007, 8, 651–661. [Google Scholar]
- Wei, H; Saladi, R; Lu, Y; Wang, Y; Palep, SR; Moore, J; Phelps, R; Shyong, E; Lebwohl, MG. Isoflavone genistein: Photoprotection and clinical implications in dermatology. J. Nutr 2003, 133, 3811S–3819S. [Google Scholar]
- Kao, TH; Chen, BH. Functional components in soybean cake and their effects on antioxidant activity. J. Agric. Food Chem 2006, 54, 7544–7555. [Google Scholar]
- Kao, TH; Huang, RF; Chen, BH. Antiproliferation of hepatoma cell and progression of cell cycle as affected by isoflavone extracts from soybean cake. Int. J. Mol. Sci 2007, 8, 1095–1110. [Google Scholar]
- Kao, TH; Wu, WM; Hung, CF; Wu, WB; Chen, BH. Anti-inflammatory effects of isoflavone powder produced from soybean cake. J. Agric. Food Chem 2007, 55, 11068–11079. [Google Scholar]
- Dalais, FS; Ebeling, PR; Kotsopoulos, D; McGrath, BP; Teede, HJ. The effects of soy protein containing isoflavones on lipids and indices of bone resorption in postmenopausal women. Clin. Endocrinol. (Oxf.) 2003, 58, 704–709. [Google Scholar]
- Migliaccio, S; Anderson, JJ. Isoflavones and skeletal health: Are these molecules ready for clinical application? Osteoporos. Int 2003, 14, 361–368. [Google Scholar]
- Chiu, TM; Huang, CC; Lin, TJ; Fang, JY; Wu, NL; Hung, CF. In vivo and in vivo anti-photoaging effects of an isoflavone extract from soybean cake. J. Ethnopharmacol 2009, 126, 108–113. [Google Scholar]
- Assefa, Z; Van Laethem, A; Garmyn, M; Agostinis, P. Ultraviolet radiation-induced apoptosis in keratinocytes: On the role of cytosolic factors. Biochim. Biophys. Acta 2005, 1755, 90–106. [Google Scholar]
- Wu, WB; Chiang, HS; Fang, JY; Chen, SK; Huang, CC; Hung, CF. (+)-Catechin prevents ultraviolet B-induced human keratinocyte death via inhibition of JNK phosphorylation. Life Sci 2006, 79, 801–807. [Google Scholar]
- Peus, D; Vasa, RA; Beyerle, A; Meves, A; Krautmacher, C; Pittelkow, MR. UVB activates ERK1/2 and p38 signaling pathways via reactive oxygen species in cultured keratinocytes. J. Invest. Dermatol 1999, 112, 751–756. [Google Scholar]
- Assefa, Z; Vantieghem, A; Garmyn, M; Declercq, W; Vandenabeele, P; Vandenheede, JR; Bouillon, R; Merlevede, W; Agostinis, P. p38 mitogen-activated protein kinase regulates a novel, caspase-independent pathway for the mitochondrial cytochrome c release in ultraviolet B radiation-induced apoptosis. J. Biol. Chem 2000, 275, 21416–21421. [Google Scholar]
- Lei, K; Davis, RJ. JNK phosphorylation of Bim-related members of the Bcl2 family induces Bax-dependent apoptosis. Proc. Natl. Acad. Sci. USA 2003, 100, 2432–2437. [Google Scholar]
- Kim, SY; Kim, SJ; Lee, JY; Kim, WG; Park, WS; Sim, YC; Lee, SJ. Protective effects of dietary soy isoflavones against UV-induced skin-aging in hairless mouse model. J. Am. Coll. Nutr 2004, 23, 157–162. [Google Scholar]
- Yarosh, D; Dong, K; Smiles, K. UV-induced degradation of collagen I is mediated by soluble factors released from keratinocytes. Photochem. Photobiol 2008, 84, 67–68. [Google Scholar]
- El-Abaseri, TB; Fuhrman, J; Trempus, C; Shendrik, I; Tennant, RW; Hansen, LA. Chemoprevention of UV light-induced skin tumorigenesis by inhibition of the epidermal growth factor receptor. Cancer Res 2005, 65, 3958–3965. [Google Scholar]
- Smith, WL; DeWitt, DL; Garavito, RM. Cyclooxygenases: Structural, cellular, and molecular biology. Annu. Rev. Biochem 2000, 69, 145–182. [Google Scholar]
- Rundhaug, JE; Fischer, SM. Cyclo-oxygenase-2 plays a critical role in UV-induced skin carcinogenesis. Photochem. Photobiol 2008, 84, 322–329. [Google Scholar]
- Isoherranen, K; Punnonen, K; Jansen, C; Uotila, P. Ultraviolet irradiation induces cyclooxygenase-2 expression in keratinocytes. Br. J. Dermatol 1999, 140, 1017–1022. [Google Scholar]
- Chen, W; Tang, Q; Gonzales, MS; Bowden, GT. Role of p38 MAP kinases and ERK in mediating ultraviolet-B induced cyclooxygenase-2 gene expression in human keratinocytes. Oncogene 2001, 20, 3921–3926. [Google Scholar]
- Tang, Q; Gonzales, M; Inoue, H; Bowden, GT. Roles of Akt and glycogen synthase kinase 3beta in the ultraviolet B induction of cyclooxygenase-2 transcription in human keratinocytes. Cancer Res 2001, 61, 4329–4332. [Google Scholar]
- Ashida, M; Bito, T; Budiyanto, A; Ichihashi, M; Ueda, M. Involvement of EGF receptor activation in the induction of cyclooxygenase-2 in HaCaT keratinocytes after UVB. Exp. Dermatol 2003, 12, 445–452. [Google Scholar]
- de Gruijl, FR. Photocarcinogenesis: UVA vs. UVB radiation. Skin Pharmacol. Appl. Skin Physiol 2002, 15, 316–320. [Google Scholar]
- Huang, CC; Fang, JY; Wu, WB; Chiang, HS; Wei, YJ; Hung, CF. Protective effects of (-)-epicatechin-3-gallate on UVA-induced damage in HaCaT keratinocytes. Arch. Dermatol. Res 2005, 296, 473–481. [Google Scholar]
- Gali-Muhtasib, HU; Perchellet, JP; Khatib, SH. Inhibitory effects of plant tannins on ultraviolet light-induced epidermal DNA synthesis in hairless mice. Photochem. Photobiol 1998, 67, 663–668. [Google Scholar]
- Moison, RM; Steenvoorden, DP; Beijersbergen van Henegouwen, GM. Topically applied eicosapentaenoic acid protects against local immunosuppression induced by UVB irradiation, cis-urocanic acid and thymidine dinucleotides. Photochem. Photobiol 2001, 73, 64–70. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (mg/g powder) | |
|---|---|---|
| Ac | acetyldaidzin | 1.11 |
| acetylglycitin | 0.06 | |
| acetylgenistin | 4.73 | |
| Ag | daidzein | 0.68 |
| glycitein | 0.18 | |
| genistein | 1.11 | |
| Total | 7.86 | |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Huang, C.-C.; Hsu, B.-Y.; Wu, N.-L.; Tsui, W.-H.; Lin, T.-J.; Su, C.-C.; Hung, C.-F. Anti-Photoaging Effects of Soy Isoflavone Extract (Aglycone and Acetylglucoside Form) from Soybean Cake. Int. J. Mol. Sci. 2010, 11, 4782-4795. https://doi.org/10.3390/ijms11124782
Huang C-C, Hsu B-Y, Wu N-L, Tsui W-H, Lin T-J, Su C-C, Hung C-F. Anti-Photoaging Effects of Soy Isoflavone Extract (Aglycone and Acetylglucoside Form) from Soybean Cake. International Journal of Molecular Sciences. 2010; 11(12):4782-4795. https://doi.org/10.3390/ijms11124782
Chicago/Turabian StyleHuang, Chieh-Chen, Bo-Yang Hsu, Nan-Lin Wu, Wen-Huei Tsui, Tzu-Ju Lin, Ching-Chieh Su, and Chi-Feng Hung. 2010. "Anti-Photoaging Effects of Soy Isoflavone Extract (Aglycone and Acetylglucoside Form) from Soybean Cake" International Journal of Molecular Sciences 11, no. 12: 4782-4795. https://doi.org/10.3390/ijms11124782