Tyrosinase-Expressing Neuronal Cell Line as in Vitro Model of Parkinson’s Disease

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
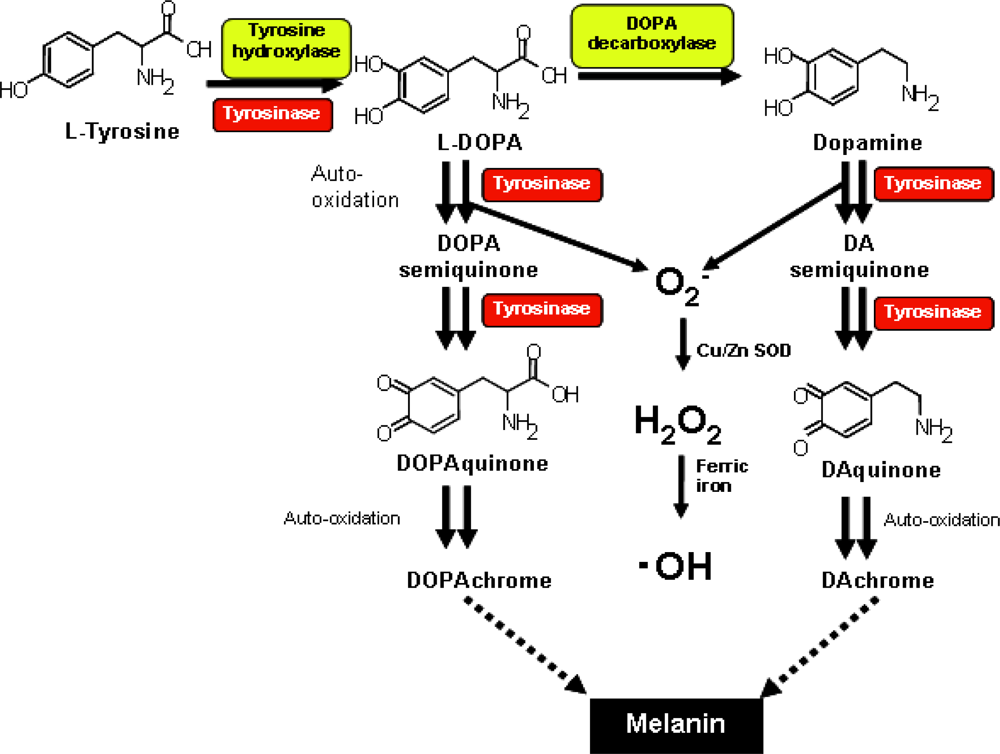
2. Establishment of Neuronal Cell Lines Inducible for the Expression of Human Tyrosinase
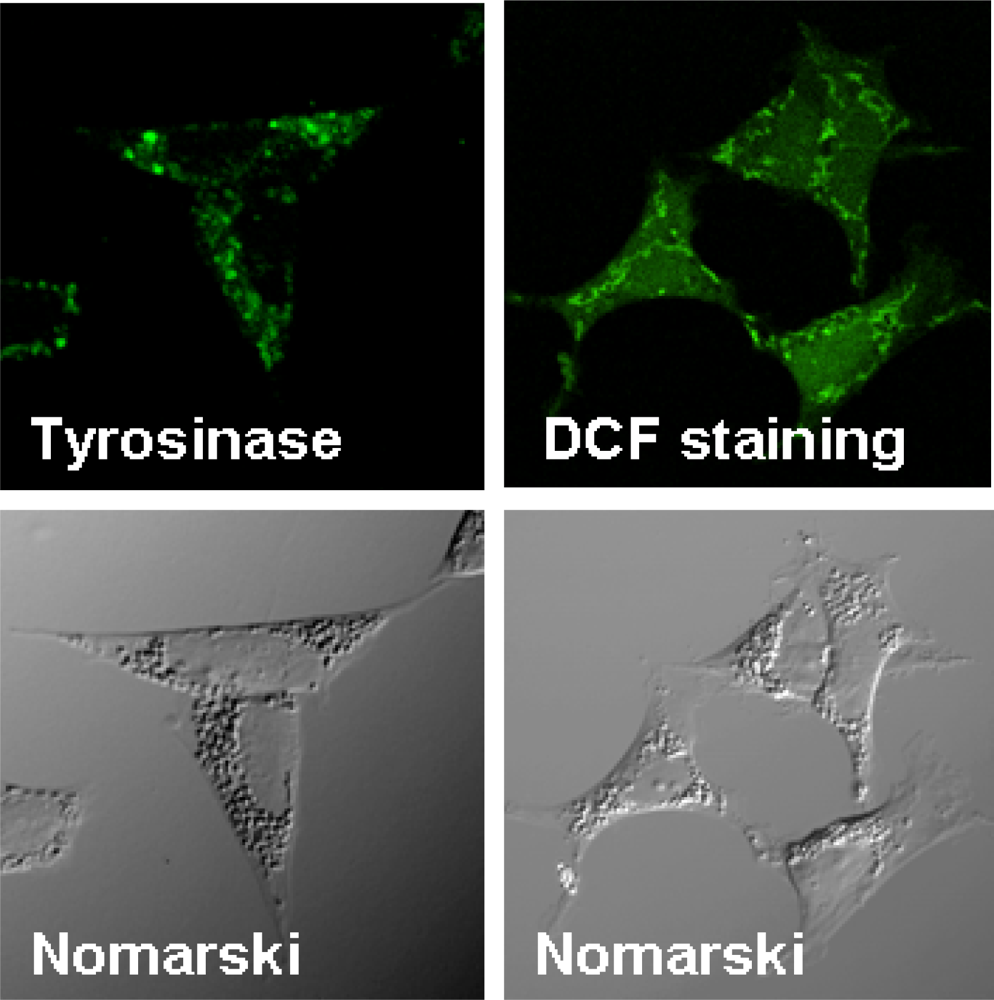
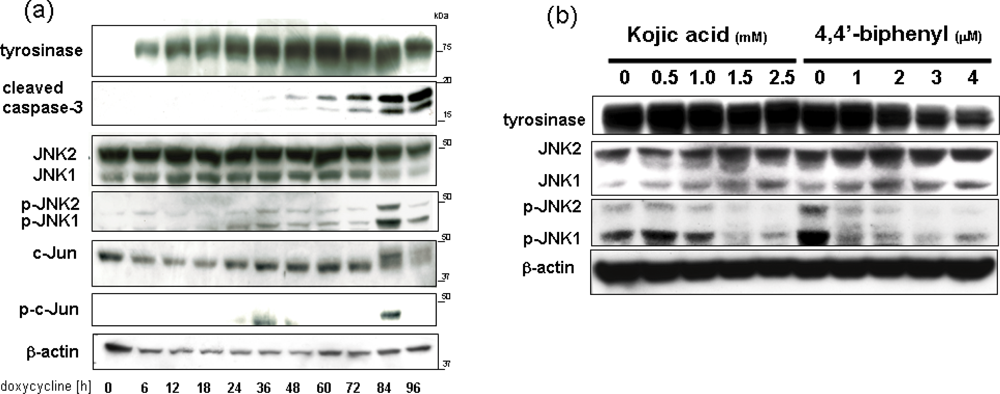
3. Overexpression of Tyrosinase Induces Neuronal Cell Death via Activation of SAPKs and Caspase 3 Apoptosis Pathway
4. Conclusions
Abbreviations:
| DA | dopamine |
| ROS | reactive oxygen species |
| TH | tyrosine hydroxylase |
| ER | endoplasmic reticulum |
| DCF | 2,7-dichlorofluorescein |
| SAPK | stress-activated protein kinase |
| MAPK | mitogen-activated protein kinase |
| JNK | c-Jun N-terminal kinase |
References
- Sayre, LM; Smith, MA; Perry, G. Chemistry and biochemistry of oxidative stress in neurodegenerative disease. Curr. Med. Chem 2001, 8, 721–738. [Google Scholar]
- Jenner, P. Oxidative stress in Parkinson's disease. Ann Neurol 2003, 53(Suppl 3), S26–S36. [Google Scholar]
- Kikuchi, A; Takeda, A; Onodera, H; Kimpara, T; Hisanaga, K; Sato, N; Nunomura, A; Castellani, RJ; Perry, G; Smith, MA; Itoyama, Y. Systemic increase of oxidative nucleic acid damage in Parkinson's disease and multiple system atrophy. Neurobiol. Dis 2002, 9, 244–248. [Google Scholar]
- Cavalieri, EL; Li, KM; Balu, N; Saeed, M; Devanesan, P; Higginbotham, S; Zhao, J; Gross, ML; Rogan, EG. Catechol ortho-quinones: The electrophilic compounds that form depurinating DNA adducts and could initiate cancer and other diseases. Carcinogenesis 2002, 23, 1071–1077. [Google Scholar]
- Ben-Shachar, D; Zuk, R; Glinka, Y. Dopamine neurotoxicity: Inhibition of mitochondrial respiration. J. Neurochem 1995, 64, 718–723. [Google Scholar]
- Stokes, AH; Hastings, TG; Vrana, KE. Cytotoxic and genotoxic potential of dopamine. J. Neurosci. Res 1999, 55, 659–665. [Google Scholar]
- Smythies, J; Galzigna, L. The oxidative metabolism of catecholamines in the brain: A review. Biochim. Biophys. Acta 1998, 1380, 159–162. [Google Scholar]
- Xu, Y; Stokes, AH; Roskoski, R; Vrana, KE. Dopamine, in the presence of tyrosinase, covalently modifies and inactivates tyrosine hydroxylase. J. Neurosci. Res 1998, 54, 691–697. [Google Scholar]
- Bolton, JL; Trush, MA; Penning, TM; Dryhurst, G; Monks, TJ. Role of quinones in toxicology. Chem. Res. Toxicol 2000, 13, 135–160. [Google Scholar]
- Asanuma, M; Miyazaki, I; Ogawa, N. Dopamine- or l-DOPA-induced neurotoxicity: The role of dopamine quinone formation and tyrosinase in a model of Parkinson's disease. Neurotox Res 2003, 5, 165–176. [Google Scholar]
- Polymeropoulos, MH; Lavedan, C; Leroy, E; Ide, SE; Dehejia, A; Dutra, A; Pike, B; Root, H; Rubenstein, J; Boyer, R; Stenroos, ES; Chandrasekharappa, S; Athanassiadou, A; Papapetropoulos, T; Johnson, WG; Lazzarini, AM; Duvoisin, RC; Di Iorio, G; Golbe, LI; Nussbaum, RL. Mutation in the alpha-synuclein gene identified in families with Parkinson's disease. Science 1997, 276, 2045–2047. [Google Scholar]
- Conway, KA; Rochet, JC; Bieganski, RM; Lansbury, PT. Kinetic stabilization of the alpha-synuclein protofibril by a dopamine-alpha-synuclein adduct. Science 2001, 294, 1346–1349. [Google Scholar]
- Norris, EH; Giasson, BI; Hodara, R; Xu, S; Trojanowski, JQ; Ischiropoulos, H; Lee, VM. Reversible inhibition of alpha-synuclein fibrillization by dopaminochrome-mediated conformational alterations. J. Biol. Chem 2005, 280, 21212–21219. [Google Scholar]
- Paris, I; Cardenas, S; Lozano, J; Perez-Pastene, C; Graumann, R; Riveros, A; Caviedes, P; Segura-Aguilar, J. Aminochrome as a preclinical experimental model to study degeneration of dopaminergic neurons in Parkinson's disease. Neurotox. Res 2007, 12, 125–134. [Google Scholar]
- Robb, DA. Tyrosinase; CRC Press: Boca Raton, FL, USA, 1984; Volume 2, pp. 207–241. [Google Scholar]
- Korner, A; Pawelek, J. Mammalian tyrosinase catalyzes three reactions in the biosynthesis of melanin. Science 1982, 217, 1163–1165. [Google Scholar]
- Sanchez-Ferrer, A; Rodriguez-Lopez, JN; Garcia-Canovas, F; Garcia-Carmona, F. Tyrosinase: A comprehensive review of its mechanism. Biochim. Biophys. Acta 1995, 1247, 1–11. [Google Scholar]
- Miranda, M; Botti, D; Bonfigli, A; Ventura, T; Arcadi, A. Tyrosinase-like activity in normal human substantia nigra. Gen. Pharmacol 1984, 15, 541–544. [Google Scholar]
- Miranda, M; Botti, D. Harding-passey mouse-melanoma tyrosinase inactivation by reaction products and activation by L-epinephrine. Gen. Pharmacol 1983, 14, 231–237. [Google Scholar]
- Takeda, A; Tomita, Y; Matsunaga, J; Tagami, H; Shibahara, S. Molecular basis of tyrosinase-negative oculocutaneous albinism. A single base mutation in the tyrosinase gene causing arginine to glutamine substitution at position 59. J. Biol. Chem 1990, 265, 17792–17797. [Google Scholar]
- Zecca, L; Tampellini, D; Gerlach, M; Riederer, P; Fariello, RG; Sulzer, D. Substantia nigra neuromelanin: Structure, synthesis, and molecular behaviour. Mol. Pathol 2001, 54, 414–418. [Google Scholar]
- Xu, Y; Stokes, AH; Freeman, WM; Kumer, SC; Vogt, BA; Vrana, KE. Tyrosinase mRNA is expressed in human substantia nigra. Brain Res. Mol. Brain Res 1997, 45, 159–162. [Google Scholar]
- Tief, K; Schmidt, A; Beermann, F. New evidence for presence of tyrosinase in substantia nigra, forebrain and midbrain. Brain Res. Mol. Brain Res 1998, 53, 307–310. [Google Scholar]
- Greggio, E; Bergantino, E; Carter, D; Ahmad, R; Costin, GE; Hearing, VJ; Clarimon, J; Singleton, A; Eerola, J; Hellstrom, O; Tienari, PJ; Miller, DW; Beilina, A; Bubacco, L; Cookson, MR. Tyrosinase exacerbates dopamine toxicity but is not genetically associated with Parkinson's disease. J. Neurochem 2005, 93, 246–256. [Google Scholar]
- Hasegawa, T; Matsuzaki-Kobayashi, M; Takeda, A; Sugeno, N; Kikuchi, A; Furukawa, K; Perry, G; Smith, MA; Itoyama, Y. Alpha-synuclein facilitates the toxicity of oxidized catechol metabolites: Implications for selective neurodegeneration in Parkinson's disease. FEBS Lett 2006, 580, 2147–2152. [Google Scholar]
- Hasegawa, T; Matsuzaki, M; Takeda, A; Kikuchi, A; Furukawa, K; Shibahara, S; Itoyama, Y. Increased dopamine and its metabolites in SH-SY5Y neuroblastoma cells that express tyrosinase. J. Neurochem 2003, 87, 470–475. [Google Scholar]
- Hasegawa, T; Treis, A; Patenge, N; Fiesel, FC; Springer, W; Kahle, PJ. Parkin protects against tyrosinase-mediated dopamine neurotoxicity by suppressing stress-activated protein kinase pathways. J. Neurochem 2008, 105, 1700–1715. [Google Scholar]
- Hasegawa, T; Sugeno, N; Takeda, A; Matsuzaki-Kobayashi, M; Kikuchi, A; Furukawa, K; Miyagi, T; Itoyama, Y. Role of Neu4L sialidase and its substrate ganglioside GD3 in neuronal apoptosis induced by catechol metabolites. FEBS Lett 2007, 581, 406–412. [Google Scholar]
- Halaban, R; Cheng, E; Zhang, Y; Moellmann, G; Hanlon, D; Michalak, M; Setaluri, V; Hebert, DN. Aberrant retention of tyrosinase in the endoplasmic reticulum mediates accelerated degradation of the enzyme and contributes to the dedifferentiated phenotype of amelanotic melanoma cells. Proc. Natl. Acad. Sci. USA 1997, 94, 6210–6215. [Google Scholar]
- Setaluri, V. Sorting and targeting of melanosomal membrane proteins: Signals, pathways, and mechanisms. Pigment Cell Res 2000, 13, 128–134. [Google Scholar]
- Toyofuku, K; Wada, I; Hirosaki, K; Park, JS; Hori, Y; Jimbow, K. Promotion of tyrosinase folding in COS 7 cells by calnexin. J. Biochem. (Tokyo) 1999, 125, 82–89. [Google Scholar]
- Orlow, SJ. Melanosomes are specialized members of the lysosomal lineage of organelles. J. Invest. Dermatol 1995, 105, 3–7. [Google Scholar]
- Simmen, T; Schmidt, A; Hunziker, W; Beermann, F. The tyrosinase tail mediates sorting to the lysosomal compartment in MDCK cells via a di-leucine and a tyrosine-based signal. J Cell Sci 1999, 112, 45–53. [Google Scholar]
- Tribl, F; Marcus, K; Meyer, HE; Bringmann, G; Gerlach, M; Riederer, P. Subcellular proteomics reveals neuromelanin granules to be a lysosome-related organelle. J. Neural. Transm 2006, 113, 741–749. [Google Scholar]
- Vijayasaradhi, S; Xu, Y; Bouchard, B; Houghton, AN. Intracellular sorting and targeting of melanosomal membrane proteins: identification of signals for sorting of the human brown locus protein, gp75. J. Cell Biol 1995, 130, 807–820. [Google Scholar]
- Singh, MV; Jimbow, K. Tyrosinase transfection produces melanin synthesis and growth retardation in glioma cells. Melanoma Res 1998, 8, 493–498. [Google Scholar]
- Bouchard, B; Fuller, BB; Vijayasaradhi, S; Houghton, AN. Induction of pigmentation in mouse fibroblasts by expression of human tyrosinase cDNA. J. Exp. Med 1989, 169, 2029–2042. [Google Scholar]
- Yamamoto, H; Takeuchi, S; Kudo, T; Sato, C; Takeuchi, T. Melanin production in cultured albino melanocytes transfected with mouse tyrosinase cDNA. Jpn. J. Genet 1989, 64, 121–135. [Google Scholar]



© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hasegawa, T. Tyrosinase-Expressing Neuronal Cell Line as in Vitro Model of Parkinson’s Disease. Int. J. Mol. Sci. 2010, 11, 1082-1089. https://doi.org/10.3390/ijms11031082
Hasegawa T. Tyrosinase-Expressing Neuronal Cell Line as in Vitro Model of Parkinson’s Disease. International Journal of Molecular Sciences. 2010; 11(3):1082-1089. https://doi.org/10.3390/ijms11031082
Chicago/Turabian StyleHasegawa, Takafumi. 2010. "Tyrosinase-Expressing Neuronal Cell Line as in Vitro Model of Parkinson’s Disease" International Journal of Molecular Sciences 11, no. 3: 1082-1089. https://doi.org/10.3390/ijms11031082