3.1. Confirmation of Measurement Conditions
We first confirmed that no rupture event occurred when streptavidin molecules were not prepared on the substrate. This is because we covered the gold surfaces of the probe and substrate with closely packed SAMs of spacer molecules to prevent direct bonding between their bare surfaces.
Secondly, the stretching motion of a PEG molecule was observed for each rupture event.
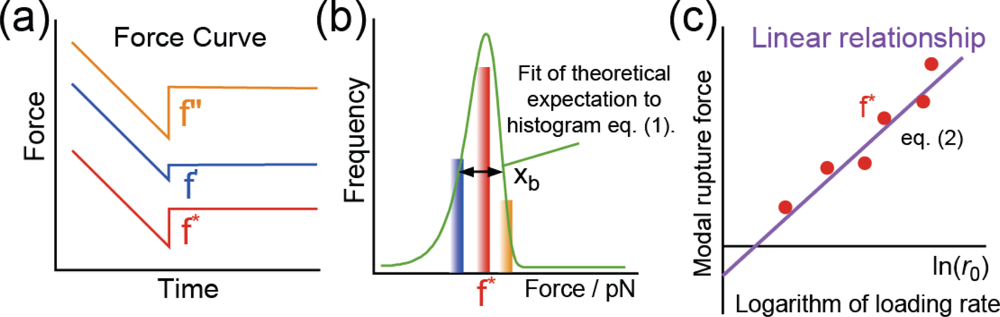
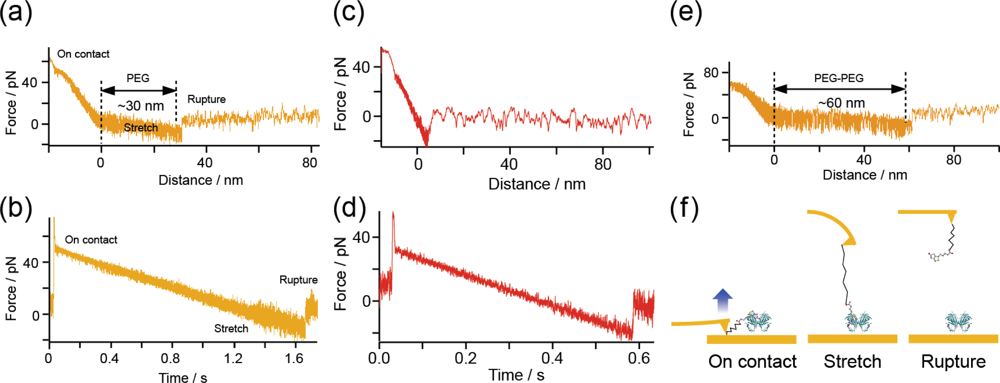
Figure 5(a),(b) show two different forms of the typical force curve obtained at a loading rate of 39 pN/s under the SM condition (
Figure 2(b)) as functions of (a) distance and (b) time. The distance represents the length of the cantilever retraction measured from the zero-force position. Since the cantilever was first moved inward until the force reached 50 pN, a region of negative distance exists. Although the force does not exhibit a linear change in (a) owing to the existence of a PEG molecule, a linear change is shown in (b). This result indicates that a constant loading rate is achieved during the stretching of the PEG molecule. The gradient of the force-distance curve in
Figure 5(a) after reaching the zero-force position is smaller than that in the negative-distance region because of the existence of the PEG molecule. The rupture force is 11.5 pN, which is the value expected for the measurement at the loading rate.
When there is a direct interaction between the cantilever and the substrate, no stretching of the PEG molecule is observed and the rupture force should be much larger than 11.5 pN (
Figure 5(c),(d)). The distance between the zero-force position and the rupture point includes the amount of bending of the cantilever, which is obtained by dividing the rupture force by the spring constant of the cantilever (6 pN/nm). Therefore, the actual length of the PEG molecule can be estimated by subtracting the amount of bending of the cantilever from the distance of 30.4 nm between the zero-force and rupture positions obtained from
Figure 5(a). The obtained length is 28 nm, which is in good agreement with the length expected from the PEG molecules that we used (molecular weight, ∼3400: length, ∼30 nm).
Figure 5(e) shows a typical force curve obtained for the B-PEG condition (
Figure 2(c)). The length of the PEG molecule, obtained from the PEG-PEG distance (61.4 nm), the rupture force (18.9 pN) and the spring constant of the cantilever (6 pN), is about 58 nm, which is in good agreement with twice the length of a single PEG molecule, as expected.
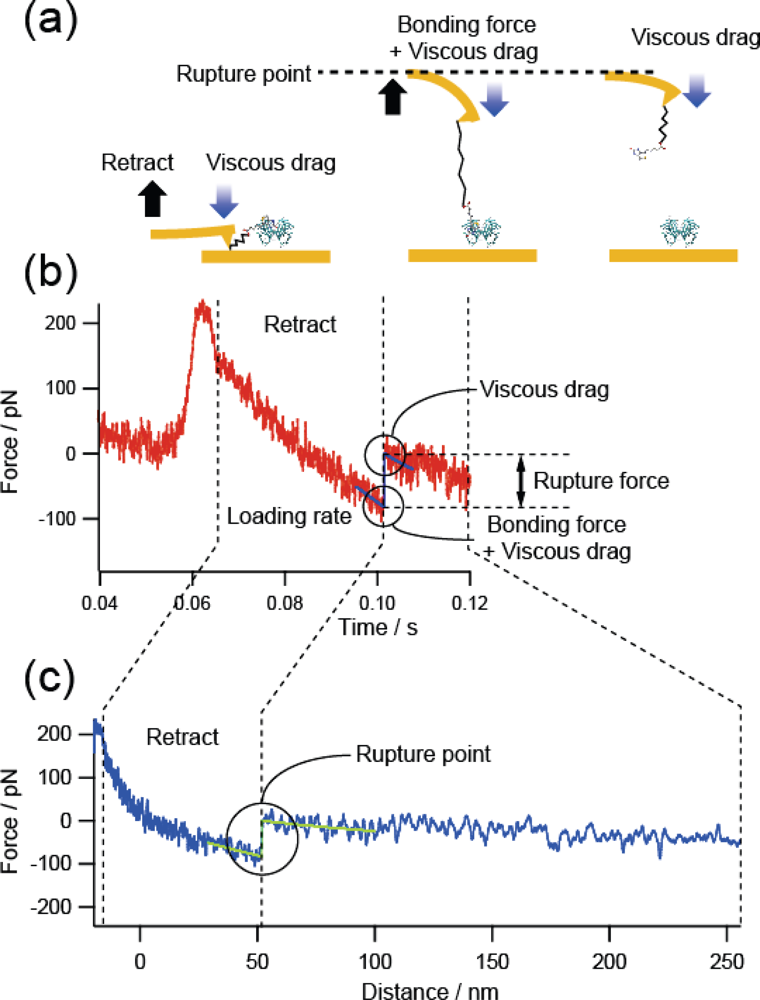
Next, we examined the effect of the viscous drag on the cantilever.
Figure 6 shows a force curve obtained at a high loading rate of 10
4 pN/s for the B-PEG condition. The observed force includes the viscous drag on the cantilever. In fact, although the cantilever (Bio-lever) we used was smaller than the cantilever typically used for AFM, an increase in force was observed after the rupture event, where the retraction of the cantilever was accelerated to realize a constant loading rate without the formation of a molecular bond. To remove such undesirable effects, we estimated the rupture force by subtracting the force immediately after the rupture event. We examined the validity of this treatment by plotting 23 histograms obtained using three cantilevers with different spring constants and shapes (6 pN/nm, rectangular, Au-coated; 30 pN/nm, rectangular, Au-coated; and 20 pN/nm, triangular, noncoated Si
2N
3). No marked dependence on the type of cantilever was observed [
15].
Thirdly, we confirmed the formation of a streptavidin-biotin structure from the reduction in bonding probability following the introduction of free biotin molecules into the solution.
Figure 7 shows the change in binding rate, obtained under the SM condition, induced by the introduction of free biotin molecules. When free biotin molecules were added, the peak signal, indicated by an arrow, gradually decreased in intensity and eventually almost disappeared, whereas the peak position did not shift, as expected (
Figure 7). The histograms in
Figure 7 appear to show distributions with different shapes. This is due to the reduction in the amount of data obtained resulting from the introduction of free biotin molecules, which greatly reduced the probability of forming the streptavidin-biotin structure. In fact, each histogram in
Figure 7, for which a sufficient amount of data was obtained, has the expected shape. This shows the importance of collecting a sufficient amount of data. In addition, the phosphate buffer ensures that the pH remains constant even when biotin molecules are added.
From these results, the observed force curves are considered to originate from the biotin-streptavidin interaction.
3.2. Rupture Force Distribution and Potential Barrier Position
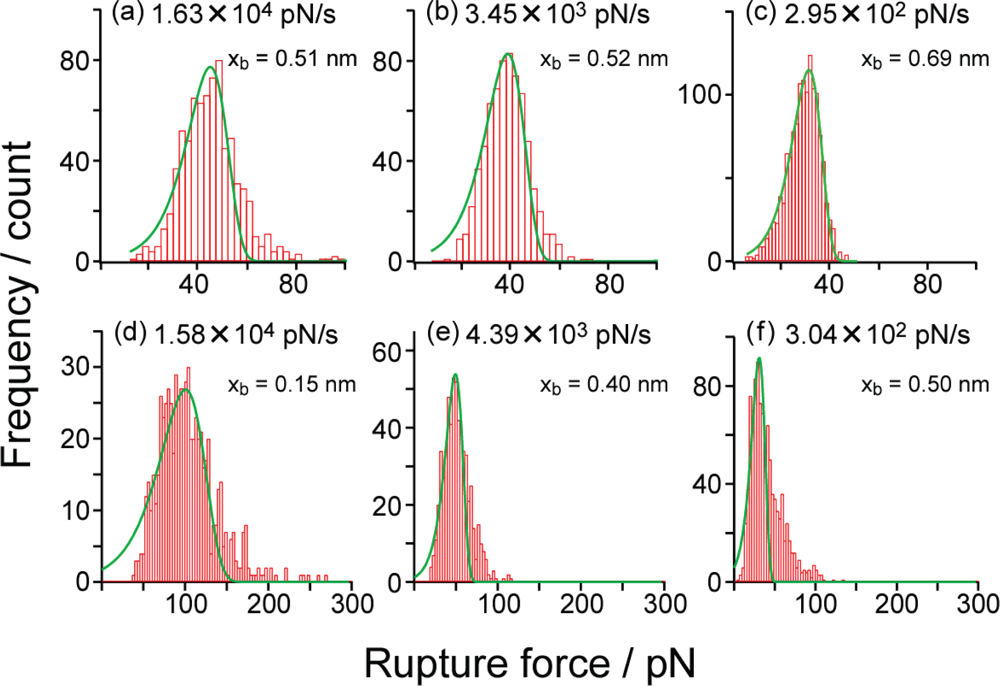
Figure 8(a)–(c) shows typical histograms of the rupture forces obtained for the SM condition in PBS (pH 7.4) at the loading rates of (a) 1.63 × 10
4 pN/s, (b) 3.45 × 10
3 pN/s and (c) 2.95 × 10
2 pN/s, where the modal rupture forces are (a) 45.1 pN, (b) 38.5 pN and (c) 31.0 pN. The modal rupture force decreases with the loading rate, as expected [
7–
26]. As mentioned in Section 2, the potential barrier position can be estimated by analyzing the shape of the rupture force distribution [
15,
16]. The solid green lines in
Figure 8 show theoretical fitting curves (
Equation 1), and the potential barrier position
xb obtained from each distribution in
Figure 8 using the method in [
15] and [
16] is determined to be (a) 0.51 nm, (b) 0.52 nm and (c) 0.69 nm. The barrier position obtained by this method is estimated to be smaller than the actual value, when there is a broadening of the histogram. The results are consistent with the barrier position of 0.68 nm obtained from the slope in
Figure 8, which is the best-fit value, because of the many data points used for estimation. The analysis based on the Bell model [
9–
26] is valid when the change in the potential shape of the system is small [
9–
26], and the obtained results validate the results of measurements performed in this study and indicate the stability of the potential barrier determined in each experiment.
On the other hand,
Figure 8(d)–(f) shows typical histograms of the rupture forces obtained for the B-PEG condition in PBS (pH 7.4). Similarly to the results obtained for the SM condition, the modal rupture force decreases with the loading rate. From the theoretical fitting curves, the potential barrier position
xb is estimated to be (d) 0.15 nm, (e) 0.40 nm and (f) 0.50 nm. Although these estimated potential barrier positions are similar to those for the SM condition, the potential barrier position obtained from the fitting in
Figure 8(d) is smaller than those obtained from the fitting in
Figure 8(e),(f).
When the Bell model can be applied with the assumption of the shape of potential landscapes described in previous studies [
9,
10,
25], as in the present case, the deformation of the histogram is considered to be small at high and low loading rates. However, the shape of the histogram is broadened at a loading rate where two potential barriers are probed, as shown in
Figure 8(d). In fact, two potential barriers are observed as will be discussed later for the results shown in
Figure 9. There are other methods for the analysis of more complex cases [
23,
24].
3.3. Analysis of Molecular Interactions–Potential Barrier Position
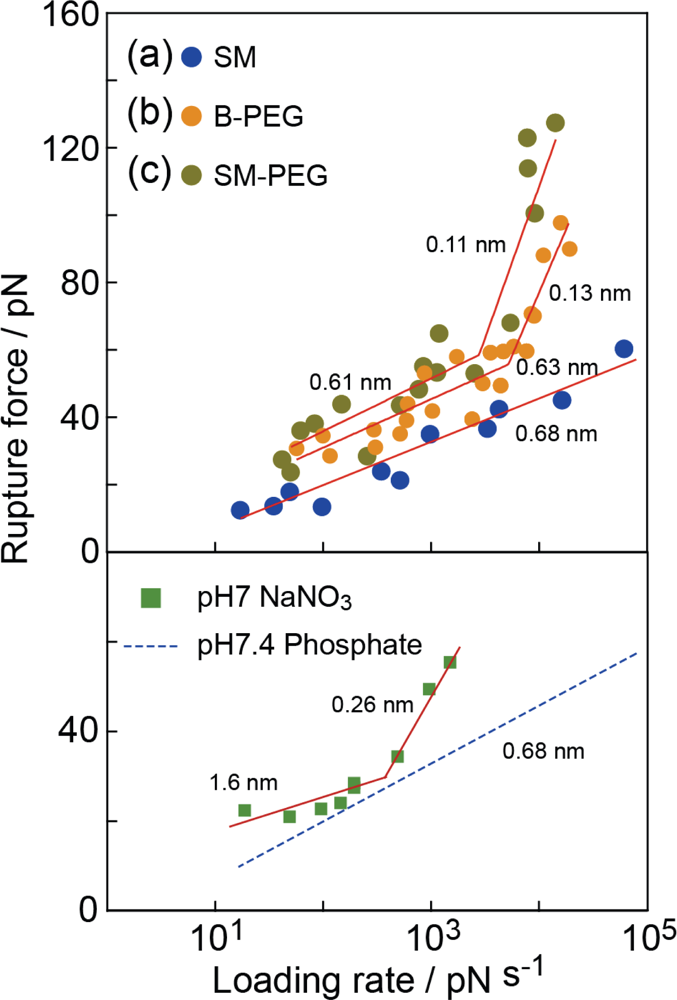
Figure 9 shows the relationships between the modal rupture force and the logarithm of the loading rate obtained in 0.01 M PBS (pH 7.4) for the (a) SM, (b) B-PEG and (c) SM-PEG conditions shown in
Figure 2. According to the DFS theory (
Equation 2), each slope provides the potential barrier of one unbinding process, and the distance of the barrier position from the potential bottom,
xb, can be deduced from the reciprocal of the gradient [
7–
26].
For the B-PEG and SM-PEG conditions, the gradient of the slopes exhibits a marked increase at 2 × 10
3 pN/s. The potential barrier positions were estimated from the two slopes to be 0.13 ± 0.01 nm and 0.63 ± 0.09 nm for the B-PEG condition and 0.11 ± 0.05 nm and 0.61 ± 0.27 nm for the SM-PEG condition, In pioneering studies on the DFS of streptavidin-biotin complex, a potential landscape with two barrier widths (0.12 nm and 0.5 nm) was observed [
9,
10]. On the basis of the results obtained by X-ray diffraction and MD calculations, the small (0.12 nm) and large (0.5 nm) potential barrier positions were attributed to the direct hydrogen bonding of the inner amino acid residues, such as ASP128, TYR43 and ASN23, with the biotin molecule, and the indirect interaction of the middle sites, such as SER45, with the biotin molecule via water molecules, respectively [
29–
31]. The results obtained under B-PEG and SM-PEG conditions are consistent with that obtained in previous studies.
For the SM condition, in contrast, only one slope was observed and the potential barrier position estimated from this slope was 0.68 ± 0.05 nm. Namely, only the bridged bonding at the middle reaction sites was observed. To confirm the validity of this result, we plotted the values from several histograms obtained using three cantilevers with different spring constants and shapes (6 pN/nm, rectangular, Au-coated; 30 pN/nm, rectangular, Au-coated; and 20 pN/nm, triangular, noncoated Si
2N
3). No marked dependence on the type of cantilever was observed. Since the waiting time (30 ms or 300 ms),
i.e., the time before the cantilever was retracted after coming into contact with the sample, had no significant effect on the results, the bonds are considered to be fully formed [
15].
Although a more detailed study is necessary to determine the mechanism, it is considered that the biotin molecule attached to the cantilever can enter deep into the binding pocket and conjugate even with the inner amino acid residues in streptavidin under the B-PEG or SM-PEG condition (
Figure 10(a)). Under the SM condition (
Figure 10(b)), in contrast, this process is prevented. This may be due to the lack of flexibility or to the effect of the hydrophobicity of the substrate surface on the formation of a hydrogen bond. Since the SAM of octanethiol molecules formed on the Au substrate has a hydrophobic characteristic, the reduction in the distance between the streptavidin molecule and the substrate may increase the effect of the hydrophobicity of the substrate. The potential barriers obtained for the 3 nm linkers in [
12] (in
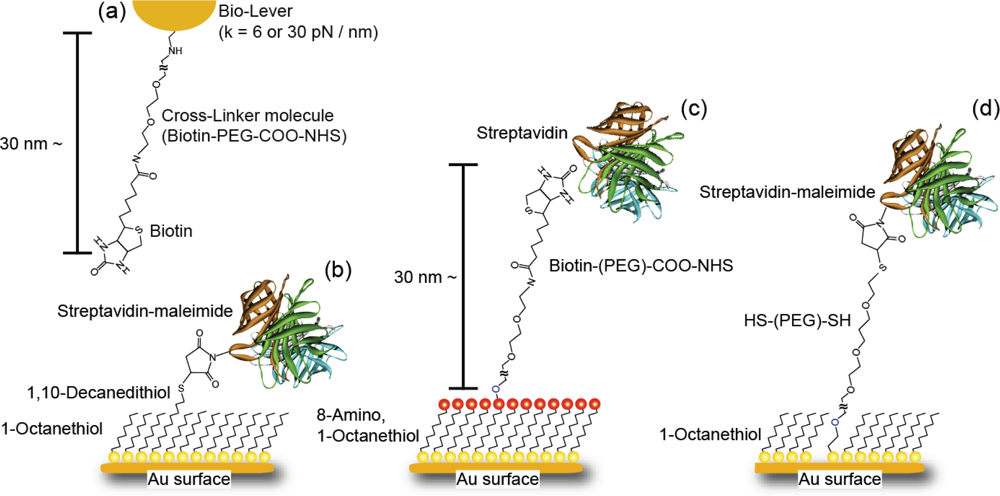
Table 1) are similar to those obtained for the B-PEG condition, suggesting that the effect of the molecular chain is negligible in the case of 3 nm linkers, enabling precise measurements. Since the linker length for the SM condition is 1.5 nm, the effect of the SM condition may occur between linker lengths of 1 nm and 3 nm.
With this method, bonding at the middle reaction sites can be selectively probed. To analyze the origins of the potential barriers of 0.68 nm and 0.63 nm in more detail, we changed the buffer solution from phosphate (pH 7.4) to sodium nitrate (pH 7) [
15]. The measurements were performed under the SM condition. For the sodium nitrate, the slope changed at a loading rate of about 10
2–10
3 pN/s, and the two potential barrier positions were estimated to be 0.26 nm and 1.6 nm.
Since no bond corresponding to the potential barriers of 0.68 nm and 0.63 nm exists in the 0.05 M sodium nitrate solution, the bond related to the potential barrier positions of 0.68 nm and 0.63 nm is suggested to be formed by molecular bridging between the amino acid residues at the middle reaction sites and the biotin molecule [
15]. Namely, the potential barrier positions of 0.68 nm and 0.63 nm are formed by the phosphate molecules in the buffer solution [
15] instead of the water molecules, in contrast to the mechanism predicted in previous studies [
9,
10].
The bonding at the inner or middle sites was distinguished and separately analyzed for the SM, B-PEG and SM-PEG conditions. To the best of our knowledge, this is the first demonstration of site-selective analysis by DFS. Using a combination of cross-linkers and the atomic force microscope that we developed for precise analysis by DFS, direct and bridging interactions at each reaction site in a ligand-receptor system were distinguished and individually analyzed.
3.4. Analysis of Molecular Interactions–Lifetime
The lifetime of bonds can be estimated from the intercept obtained by extrapolating the linear relationship between the modal rupture force and the logarithm of loading rate (
Equation 2), as shown in
Figure 9 [
7–
26]. The rupture forces obtained for the B-PEG condition were larger than those for the SM condition at all loading rates, as shown in
Figure 9, and the lifetimes obtained for the SM, B-PEG and SM-PEG conditions were 1.0 s, 6.4 s and 13 s, respectively.
Since two rupture points exist for the B-PEG condition (
Figure 11(a)) and one rupture point exists for the SM-PEG condition (
Figure 11(b)), the rupture probability for the B-PEG condition is considered to be two times larger than that for the SM-PEG condition. Namely, the lifetime for the B-PEG condition is expected to be half that for the SM-PEG condition. From the result of the B-PEG (PEG-biotin-streptavidin-biotin-PEG) condition (6.4 s), the lifetime for streptavidin-biotin-PEG is estimated to be 12.8 s [
12]. This value is in good agreement with the lifetime obtained for the SM-PEG condition (13 s). The obtained values are ten times larger than that obtained for the SM condition. This is caused by the fact that the direct bonds with the inner amino acid residues, such as ASP128, TYR43 and ASN23, are stable under the B-PEG and SM-PEG conditions because the effect of the hydrophobicity of the substrate, for example, is less than that under the SM condition. Similarly to the potential barriers mentioned in Section 3.3, the lifetime obtained for the 3 nm linkers in the previous experiment [
12] (in
Table 1) is similar to that obtained for the B-PEG condition, suggesting that the effect of the SM condition is negligible at 3 nm from the substrate in the case of steric hindrance.
The lifetimes obtained from DFS measurements are, in general, much shorter than that expected from the dissociation constant (
Kd ∼ 10
−15) of the streptavidin-biotin complex [
28,
29]. This is because the dissociation constant is measured under an equilibrium condition where biotin molecules form bonds with all three group sites in streptavidin. However, the lifetime of each bond is evaluated using the results obtained by DFS measurement instead of the total strength, resulting in the observed shortening of the lifetime.
The lifetime, and thereby the observed rupture force, changes with the experimental conditions. Despite this fact, different conditions were used in previous experiments. Namely, some data were obtained using a cantilever without a PEG molecule, and other data were obtained using a PEG molecule at different buffer concentrations. In addition, the loading rate was not kept constant. Therefore, the difference in lifetime between our study and previous studies (for example, that indicated in [
9] may be caused by the differences in pH and ion concentration. Such variations among the reported values must be due to important effects yet to be understood in detail, which, therefore, must be clarified in the future. In any case, a precise DFS measurement technique will be essential for the analysis.
3.5. Analysis of Molecular Interactions – Reaction Processes
The results obtained in this study are shown in
Table 2. As shown in
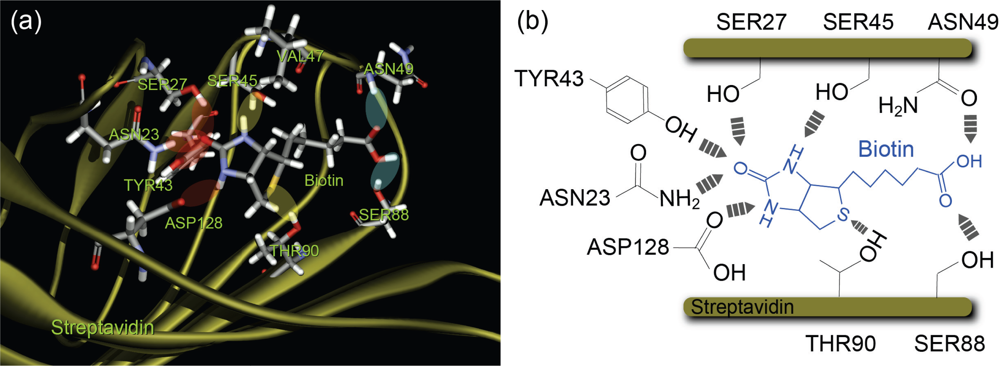
Figure 1, the binding pocket of streptavidin has several reaction sites with a hydrogen-bonding network, which are classified into the following three groups depending on the distance from the bottom of the binding pocket: (1) the inner binding sites of amino acid residues SER27, ASN23, TYR43 and ASP128, (2) the middle binding sites SER45 and THR90, and (3) the outer binding sites ASN49 and SER88 [
28,
29]. The barrier positions observed correspond to these three types: two direct bonds at inner and middle sites, and one indirect bond via solvent molecules at middle sites. Since the –COOH of the biotin molecule was attached to the PEG molecule, the interactions between the –COOH of the biotin molecule and the outer binding sites ASN 49 and SER88 are considered to be very weak, resulting in the absence of potential barriers between a biotin-PEG molecule and the outer binding sites.
These results obtained by site-selective analysis enable us to discuss the step-by-step processes for various conditions: For the B-PEG and SM-PEG conditions in PBS, the biotin molecule is trapped at the inner sites due to direct bonding and at the middle sites via the bridging of buffer molecules (
Figure 12(a)). For the SM condition in PBS, the biotin molecule is not trapped at the inner sites but only at the middle sites via the bridging of buffer molecules (
Figure 12(b)). For the SM condition in sodium nitrate solution, the biotin molecule is trapped at the middle sites via the hydrogen bonding of amino acid residues (
Figure 12(c)).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}