Effects of Time of Initial Exposure to MSV Sarcoma on Bone Induction by Dentine Matrix Implants and on Orthotopic Femora

Abstract
:1. Introduction
2. Materials and Methods
2.1. Demineralized Incisor Preparation and Characteristics
2.2. Implantation of Demineralized Incisors
2.3. Assessment of Excised Implants and of Femoral Bone
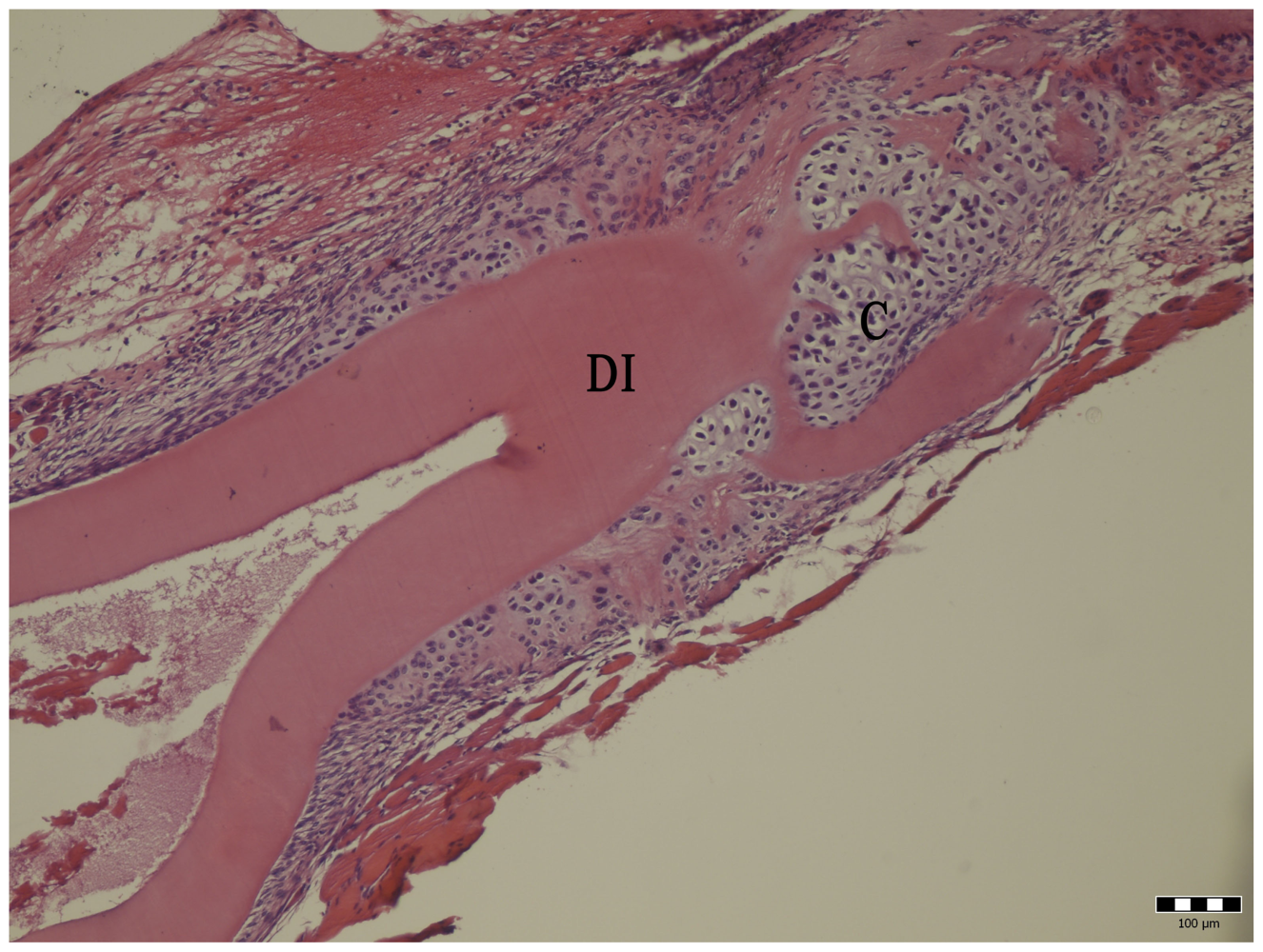
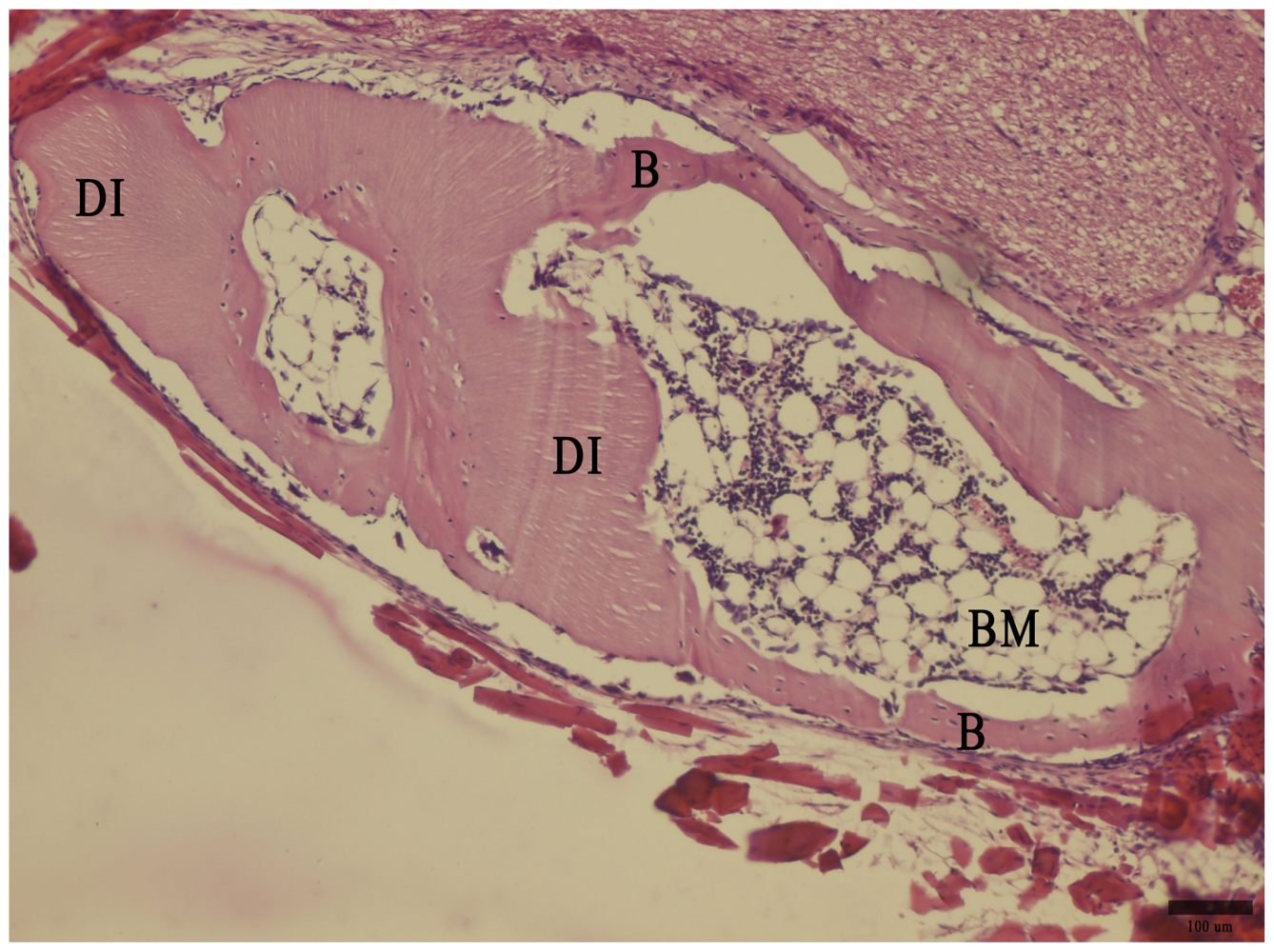
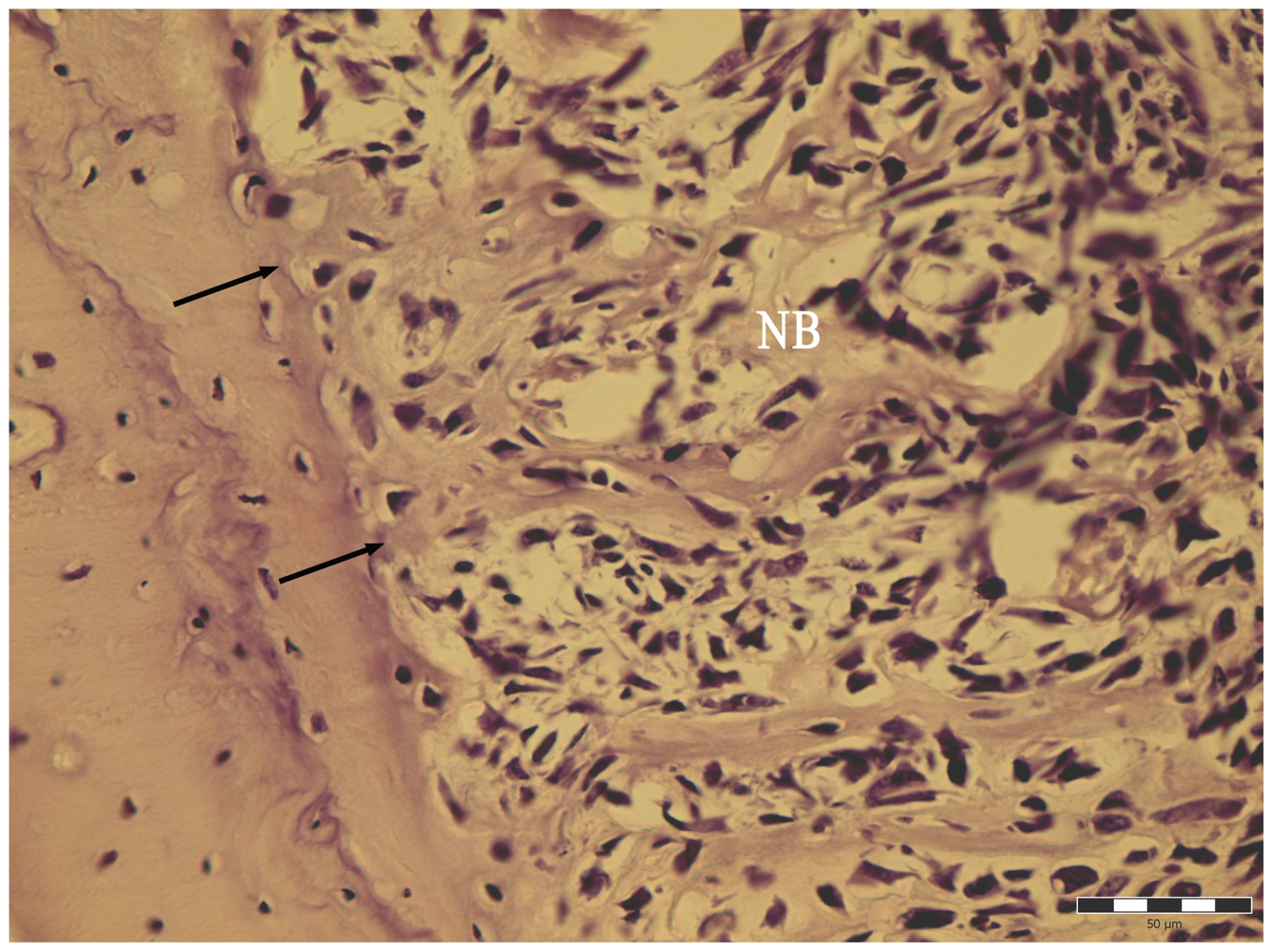
2.4. Histology
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusion
References
- Włodarski, K; Kobus, M; Łuczak, M. Orthotopic bone induction at sites of Moloney murine sarcoma virus inoculation in mice. Nature 1979, 281, 386–387. [Google Scholar]
- Fefer, A; McCoy, JL; Glynn, JP. Induction and regression of primary Moloney sarcoma virus-induced tumors in mice. Cancer Res 1967, 27, 1626–1631. [Google Scholar]
- Fefer, A; McCoy, JL; Glynn, JP. Immunologic, virologic and pathological studies of regression of autochtonous Moloney sarcoma virus-induced tumors in mice. Cancer Res 1968, 28, 1577–1585. [Google Scholar]
- Fefer, A. Immunotherapy and chemotherapy of Moloney sarcoma virus-induced tumors in mice. Cancer Res 1969, 29, 2177–2183. [Google Scholar]
- Stutman, O. Delayed tumor appearance and absence of regression in nude mice infected with murine sarcoma virus. Nature 1975, 253, 142–144. [Google Scholar]
- Russel, SW; McIntosh, AT. Macrophages isolated from regressing Moloney sarcoma are more cytotoxic than those recovered from progressing sarcomas. Nature 1977, 268, 68–71. [Google Scholar]
- Friedlander, GE; Mitchell, MS. A virally induced osteosarcoma in rats. A model for immunological studies of human osteosarcoma. J. Bone Joint Surg 1976, 58, 295–302. [Google Scholar]
- Huggins, CB; Urist, MR. Dentine matrix transplantation. Rapid induction of alkaline phosphatase and cartilage. Science 1970, 167, 896–898. [Google Scholar]
- Urist, MR. Bone histogenesis and morphogenesis in implants of demineralized enamel and dentine. J. Oral Surg 1971, 29, 88–102. [Google Scholar]
- Amar, S; Sires, B; Clohisy, J; Veis, A. The isolation and partial characterization of a root incisor dentin matrix polypeptide with in vitro chondrogenic activity. J. Biol. Chem 1991, 266, 8609–8618. [Google Scholar]
- Ike, M; Urist, MR. Recycled dentine root matrix for a carrier of recombinant human bone morphogenetic protein. J. Oral Implantol 1998, 24, 124–132. [Google Scholar]
- Wang, EA; Rosen, V; Cordes, P; Hewick, RM; Kriz, MJ; Luxenberg, DP; Sibley, BS; Wozney, JM. Purification and characterization of other distinct bone-inducing factors. Proc. Natl. Acad. Sci. USA 1988, 85, 9484–9488. [Google Scholar]
- Wozney, JM; Rosen, V; Celeste, AJ; Mitsock, LM; Whitters, MJ; Kriz, RW; Hewick, RM; Wang, EA. Novel regulators of bone formation: molecular clones and activities. Science 1988, 242, 1528–1534. [Google Scholar]
- Sampath, TK; Maliakal, JC; Hauschka, PV; Jones, WK; Sasak, H; Tucker, RF; White, KH; Coughlin, JE; Tucker, MM; Pang, RH; et al. Recombinant human osteogenic protein-1 (hOP-1) induces new bone formation in vivo with a specific activity comparable with natural bovine osteogenic protein and stimulates osteoblast proliferation and differentiation in vitro. J. Biol. Chem 1992, 267, 20352–20362. [Google Scholar]
- De Vlam, K; Lories, RJ; Luyten, FP. Mechanisms of pathological new bone formation. Curr. Rheumatol. Rev 2006, 8, 332–337. [Google Scholar]
- Włodarski, K; Reddi, AH. Heterotopically induced bone does not develop functional periosteal membrane. Arch. Immunol. Therap. Exp 1986, 34, 385–395. [Google Scholar]
- Kaysinger, KK; Ramp, WK; Lang, GJ; Gruber, HE. Comparison of human osteoblasts and osteogenic cells from heterotopic bone. Clin. Orthop. Rel. Res 1997, 342, 181–191. [Google Scholar]
- Tom, A; Arend, A; Gunnarsson, D; Ulfsparre, R; Suutre, S; Haviko, T; Selstam, G. Bone formation zones in heterotopic ossifications: histology findings and increase expression of bone morphogenetic protein 2 and transforming growth factors beta2 and beta3. Calcif. Tissue. Int 2007, 80, 259–267. [Google Scholar]
- Chauvean, D; Devedjian, JC; Blary, MC; Delecourt, C; Hardouin, P; Jeanfils, J; Broux, O. Gene expression in human osteoblastic cells from normal and heterotopic ossification. Exp. Mol. Pathol 2004, 76, 37–43. [Google Scholar]
- Brownell, AG. Osteogenesis inhibitory protein: a (p)review. Connect. Tissue Res 1990, 24, 13–16. [Google Scholar]
- Amedee, J; Bareille, R; Rouais, F; Cunningham, N; Reddi, AH; Harmand, MF. Osteogenin (bone morphogenetic protein 3) inhibits proliferation and stimulates differentiation of osteoprogenitors in human bone marrow. Differentiation 1994, 58, 157–164. [Google Scholar]
- Maliakal, JC; Asahima, I; Hauschka, PV; Sampath, TK. Osteogenic protein-1 (BMP-7) inhibits cell proliferation and stimulates the expression of markers characteristicc of osteoblast phenotype in rat osteosarcoma (17/2.8) cells. Growth Factors 1994, 11, 227–234. [Google Scholar]
- Goldring, SR. Inflammatory mediators as essential elements in bone remodeling. Calcif. Tissue Int 2003, 73, 97–100. [Google Scholar]
- Frost, A; Jonsson, KB; Nilsson, O; Ljunggern, O. Inflammatory cytokines regulate proliferation of cultured human osteoblasts. Acta. Orthop. Scand 1997, 68, 91–96. [Google Scholar]
- Tomoda, K; Kitaoka, M; Iyama, K; Usuku, G. Endosteal new bone formation in the long bones of adjuvant treated rats. Pathol. Res. Pract 1986, 181, 331–338. [Google Scholar]
- Włodarski, P; Galus, R; Włodarski, KH; Brodzikowska, A. Heterotopic osteogenesis by murine demineralized incisors at lesions sites induced by Concanavalin A in mice. Connect. Tissue. Res 2009, 50, 1–6. [Google Scholar]
- Chanaille, P; Levy, JP; Tacitan, GJ. Routine method for concentration and partial purification of a murine leukemia virus (Rausher). Nature 1967, 213, 107–109. [Google Scholar]
- Włodarski, P; Sevignani, C; Fernandez, MJ; Calabretta, B; Włodarski, KH. Tumor induced by Moloney sarcoma virus causes periosteal osteogenesis engaging osteopontin, stromelysin-1 and tenascin. Neoplasma 2007, 54, 173–179. [Google Scholar]
- Włodarski, K; Galus, K. Osteoblastic and chondroblastic response to variety of locally administered immunomodulators in mice. Folia Biol. (Praha) 1992, 38, 284–292. [Google Scholar]
- Friedenstein, AJ. Precursor cells of mechanocytes. Int. Rev. Cytol 1976, 47, 327–359. [Google Scholar]
- Ohno, T; Oboki, K; Kajiwara, N; Morii, E; Aozasa, K; Flawell, RA; Okumura, K; Saito, H; Nakae, S. Caspase-1, caspase-8, and calpain are dispensable for IL-33 release by macrophage. J. Immunol 2009, 183, 7890–7897. [Google Scholar]
- Gocheva, V; Wang, HW; Gadea, BB; Shree, T; Hunter, KE; Garfall, AL; Berman, T; Joyce, JA. IL-4 induces cathepsin protease activity in tumor-associated macrophages to promote cancer growth and invasion. Genes Dev 2010, 24, 241–255. [Google Scholar]
- Schett, G; Sieper, J. Inflammation and repair mechanisms. Clin. Rheumatol 2009, 27, 33–35. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of MSV-inoculation post incisor implantation (day) | Yield of bone induction (mg) at MSV-sarcoma lesions* | Yield of bone induction (mg) at contralateral site, not exposed to MSV* | Yield of femoral bone increment following MSV sarcoma exposure (% of original weight)** |
|---|---|---|---|
| 4 | (53) 0.37 ± 0.34 | (55) 0.46 ± 0.38 | (90) 17.6 ± 16.7 |
| 12 | (32) 0.73 ± 0.39 | (32) 0.48 ± 0.34 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Włodarski, K.; Włodarski, P.; Galus, R.; Brodzikowska, A. Effects of Time of Initial Exposure to MSV Sarcoma on Bone Induction by Dentine Matrix Implants and on Orthotopic Femora. Int. J. Mol. Sci. 2010, 11, 3277-3287. https://doi.org/10.3390/ijms11093277
Włodarski K, Włodarski P, Galus R, Brodzikowska A. Effects of Time of Initial Exposure to MSV Sarcoma on Bone Induction by Dentine Matrix Implants and on Orthotopic Femora. International Journal of Molecular Sciences. 2010; 11(9):3277-3287. https://doi.org/10.3390/ijms11093277
Chicago/Turabian StyleWłodarski, Krzysztof, Paweł Włodarski, Ryszard Galus, and Aniela Brodzikowska. 2010. "Effects of Time of Initial Exposure to MSV Sarcoma on Bone Induction by Dentine Matrix Implants and on Orthotopic Femora" International Journal of Molecular Sciences 11, no. 9: 3277-3287. https://doi.org/10.3390/ijms11093277