Effect of Peptide Size on Antioxidant Properties of African Yam Bean Seed (Sphenostylis stenocarpa) Protein Hydrolysate Fractions

Abstract
:1. Introduction
2. Results and Discussion
2.1. Amino Acid Composition
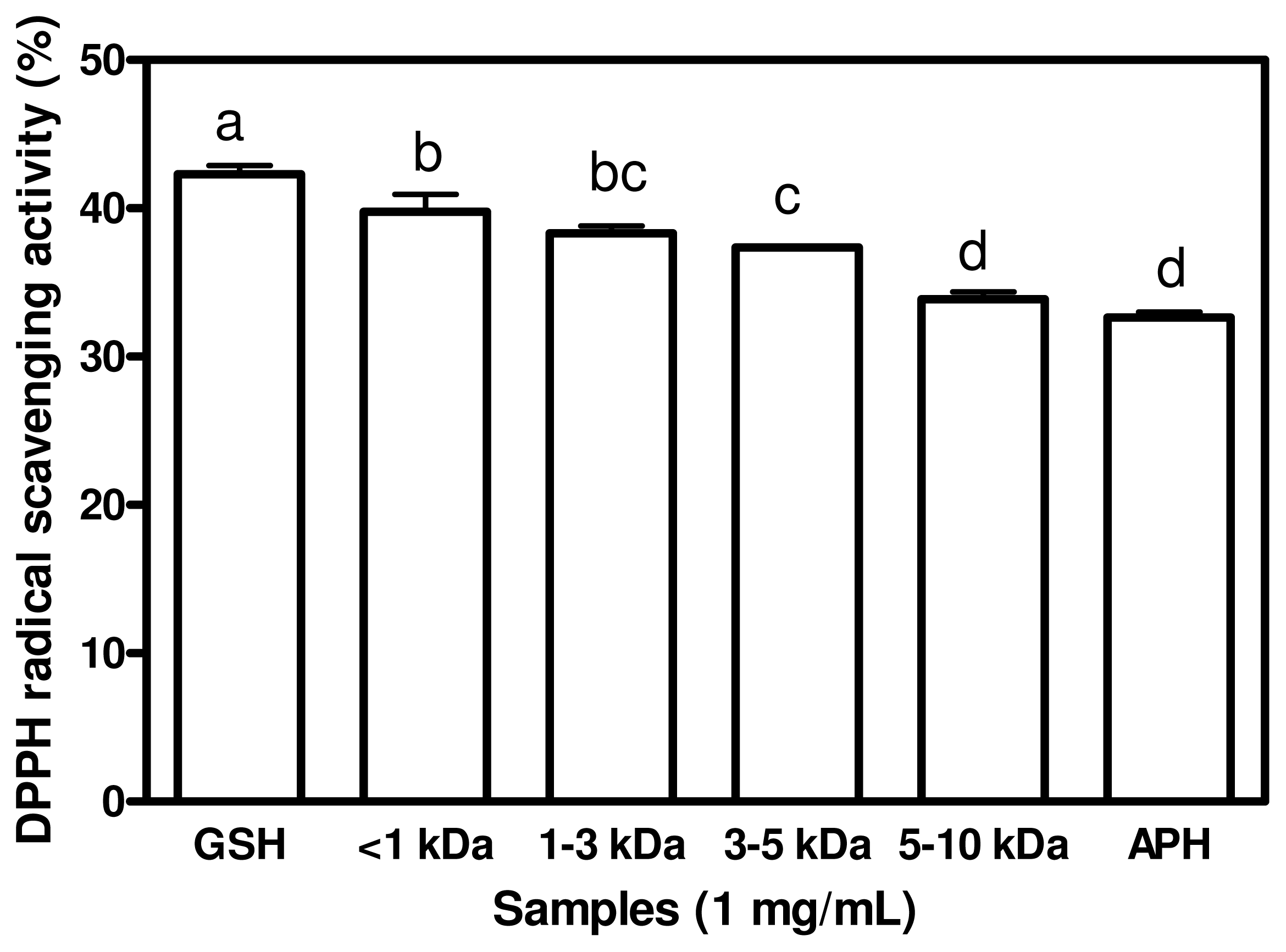
2.2. DPPH Radical Scavenging Activities
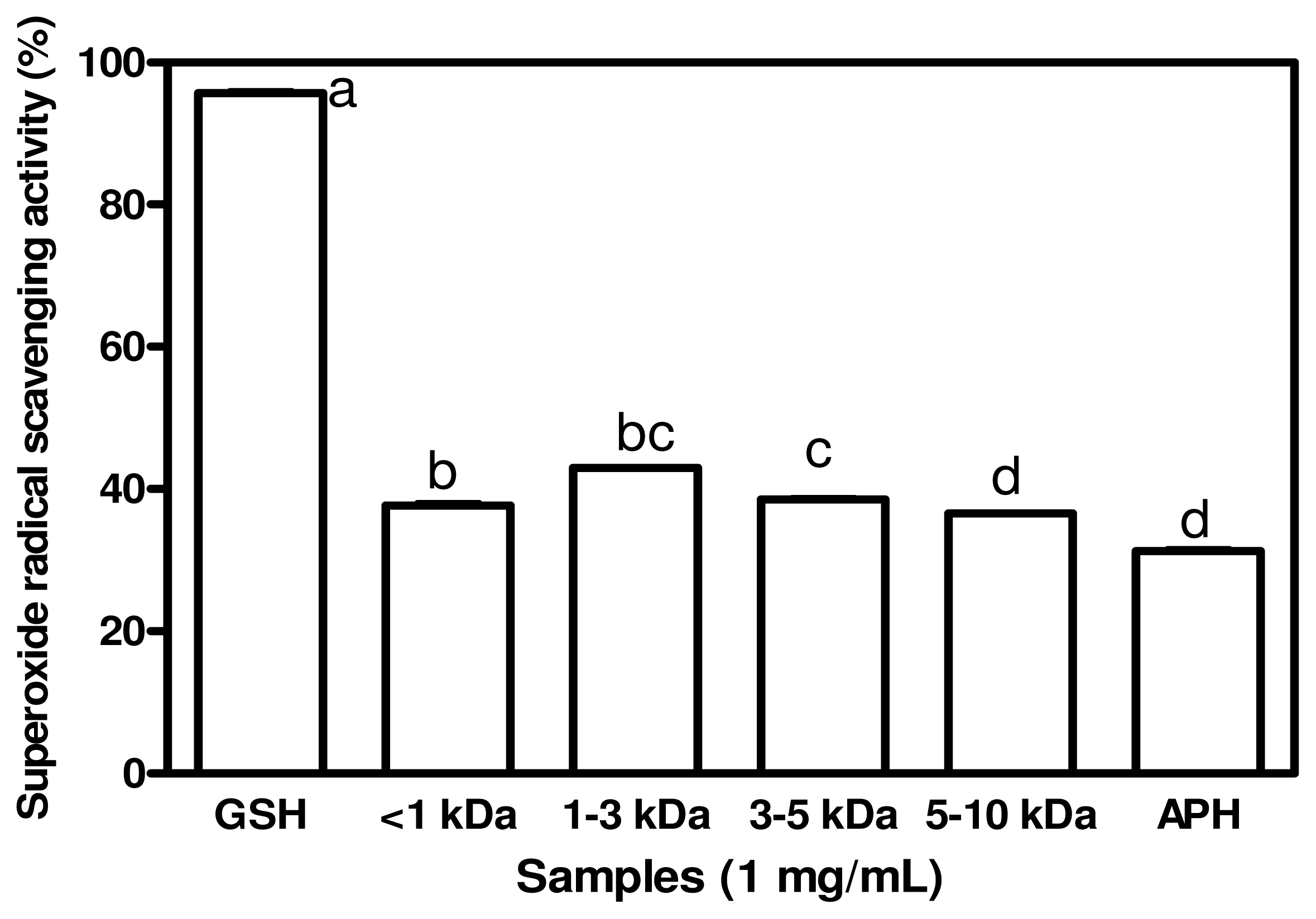
2.3. Superoxide Radical Scavenging Activities
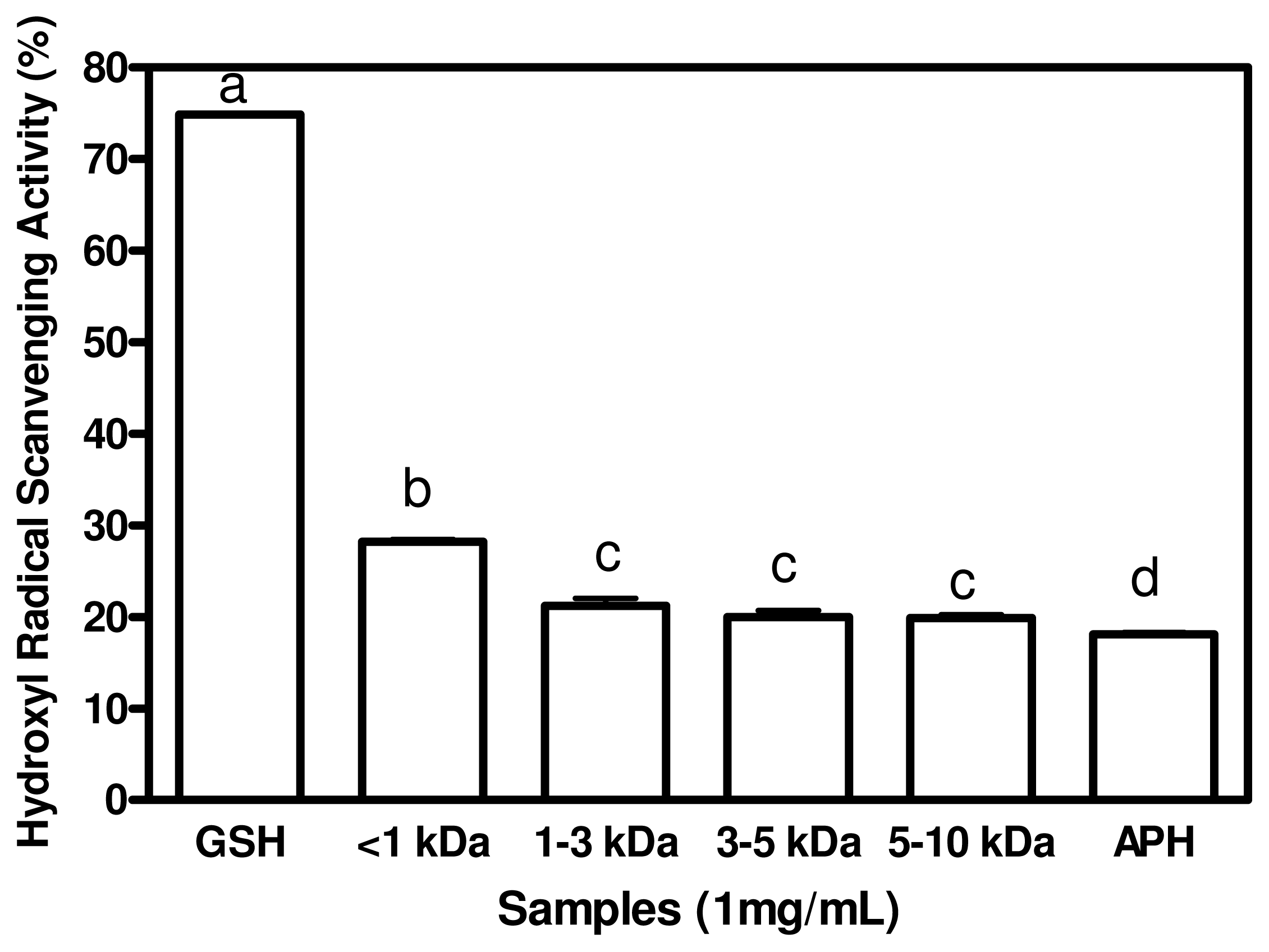
2.4. Hydroxyl Radical Scavenging Activities
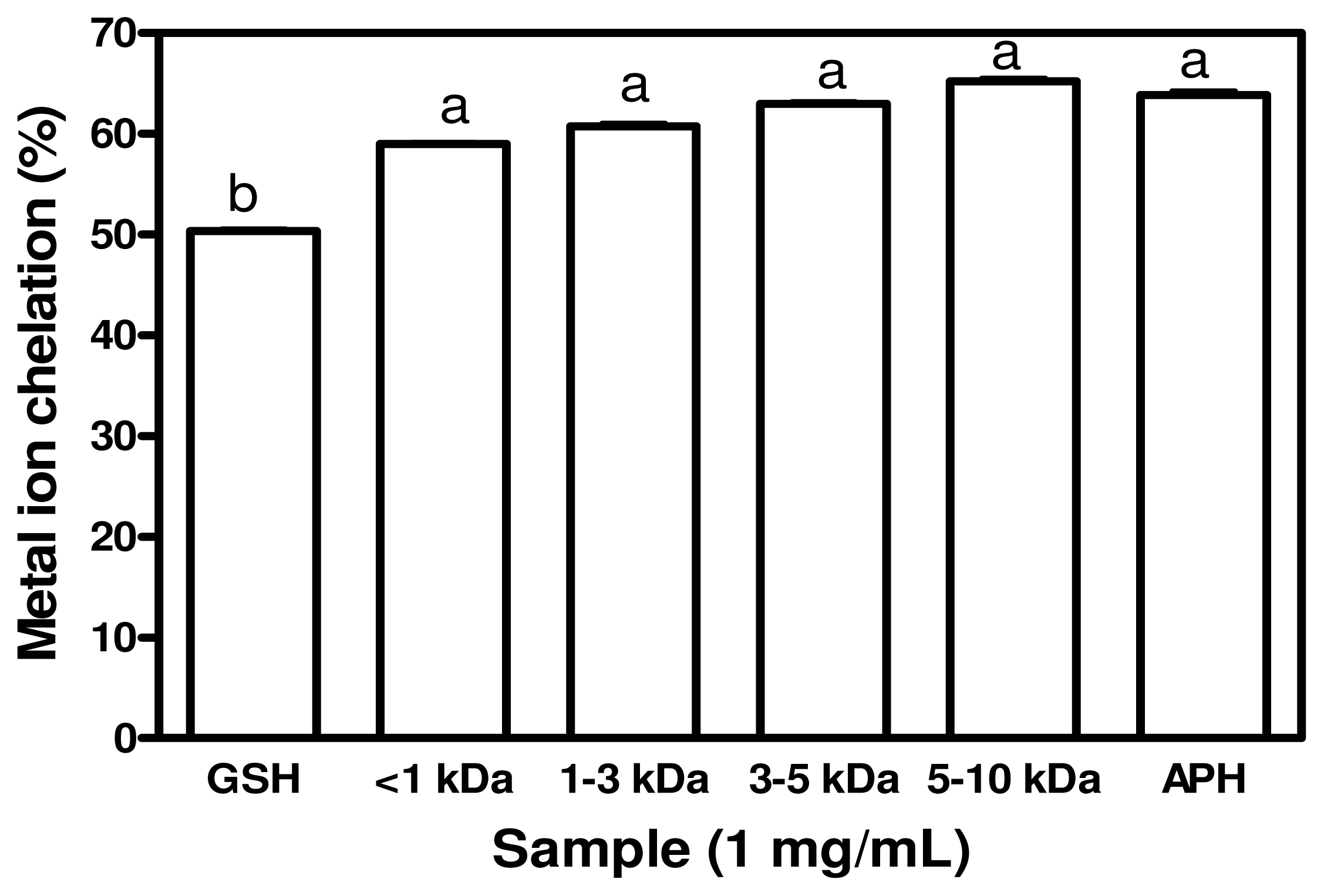
2.5. Metal Chelating Activity
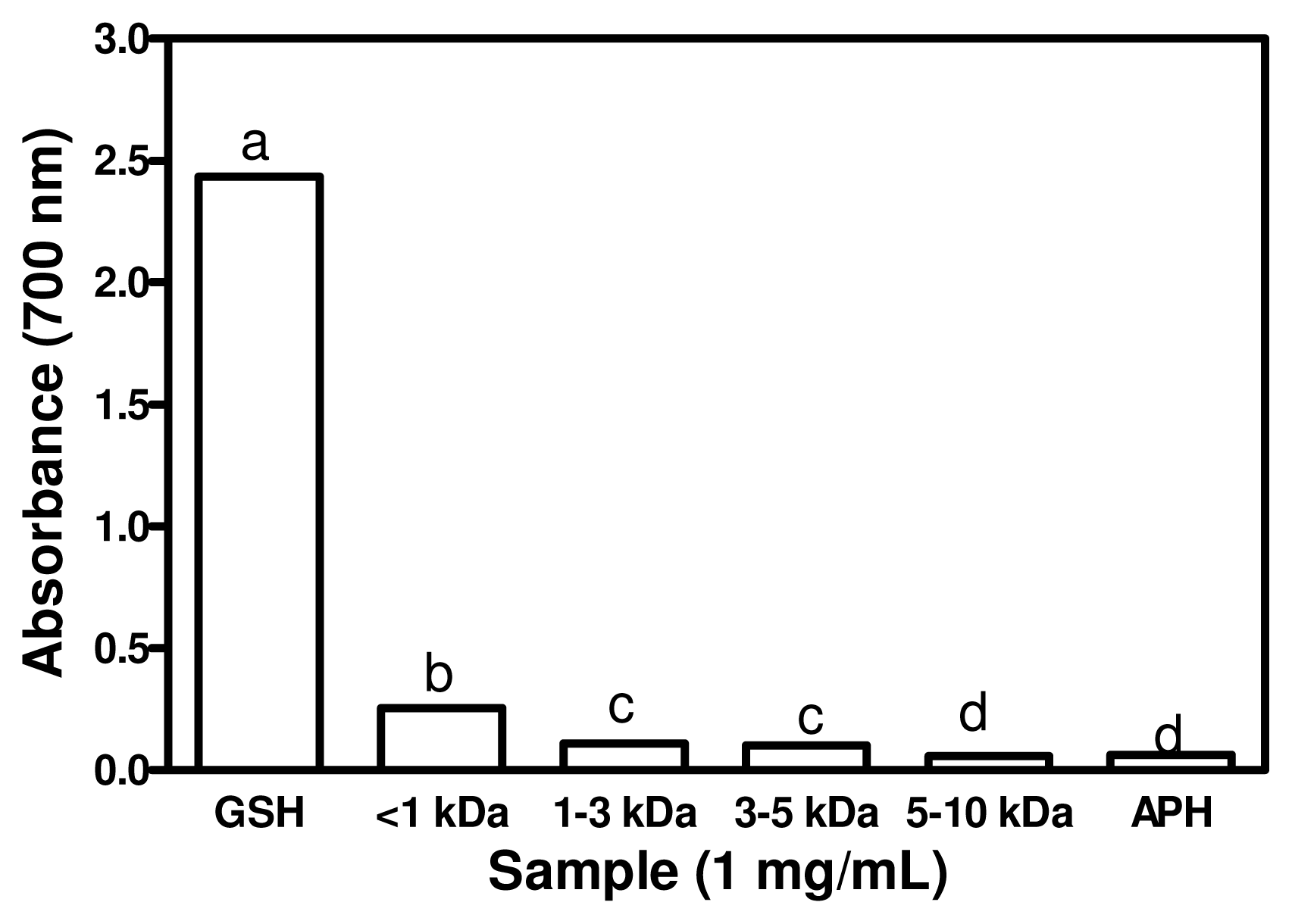
2.6. Ferric Reducing Power Activity (FRAP)
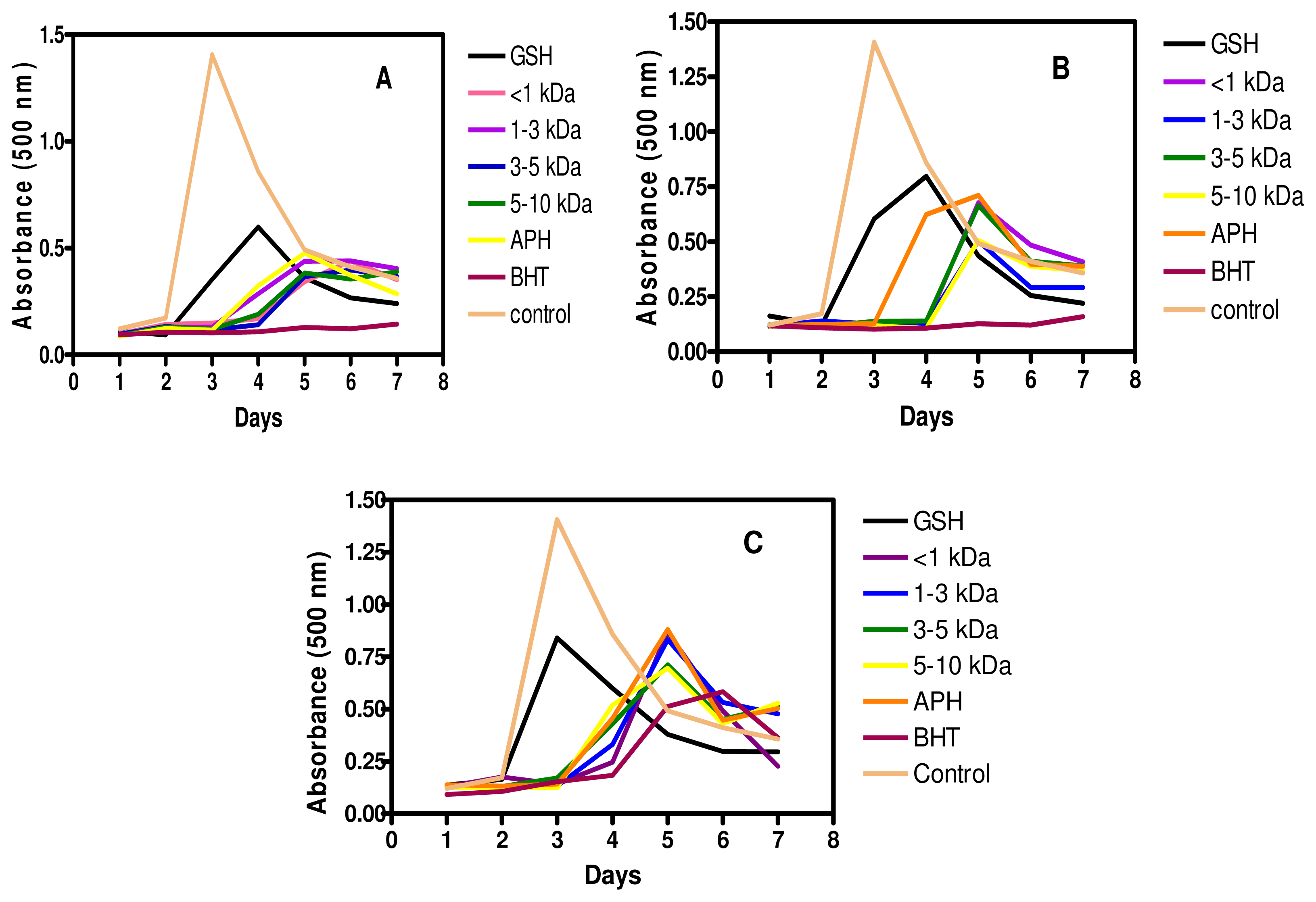
2.7. Inhibition of Linoleic Acid Oxidation
3. Experimental Section
3.1. Materials
3.2. Preparation of AYB Seed Protein Isolate (API)
3.3. Preparation of AYB Protein Hydrolysate (APH) and Membrane Fractions
3.4. Amino Acid Analysis
3.5. DPPH Radical Scavenging Assay
3.6. Assay of Metal Ion Chelation
3.7. Ferric Reducing Power Assay
3.8. Hydroxyl Radical Scavenging Assay
3.9. Superoxide Scavenging Assay
3.10. Inhibition of Linoleic Acid Oxidation
3.11. Statistics Analysis
4. Conclusions
Acknowledgments
References
- Zhu, L; Chen, J; Tang, X; Xiong, YL. Reducing, radical scavenging, and chelation properties of in vitro digests of alcalase-treated zein hydrolysate. J. Agric. Food Chem 2008, 56, 2714–2721. [Google Scholar]
- Xiong, Y; Decker, EA. Alterations in muscle protein functionality by oxidative and antioxidative processes. J. Muscle Foods 1995, 6, 139–160. [Google Scholar]
- Girgih, AT; Udenigwe, CC; Aluko, RE. In vitro antioxidant properties of hemp seed protein hydrolysate fractions. J. Am. Oil Chem. Soc 2011, 88, 381–389. [Google Scholar]
- Pownall, TL; Udenigwe, CC; Aluko, RE. Amino acid composition and antioxidant properties of pea seed (Pisum sativum L.) enzymatic protein hydrolysate fractions. J. Agric. Food Chem 2010, 58, 4712–4718. [Google Scholar]
- Zhu, K; Zhou, H; Qian, H. Antioxidant and free radical-scavenging activities of wheat germ protein hydrolysates (WGPH) prepared with alcalase. Proc. Biochem 2006, 41, 1296–1302. [Google Scholar]
- Moure, A; Domínguez, H; Parajó, JC. Antioxidant properties of ultrafiltration-recovered soy protein fractions from industrial effluents and their hydrolysates. Proc. Biochem 2006, 41, 447–458. [Google Scholar]
- Li, Y; Jiang, B; Zhang, T; Mu, W; Liu, J. Antioxidant and free radical-scavenging activities of chickpea protein hydrolysate (CPH). Food Chem 2008, 106, 444–450. [Google Scholar]
- Cumby, N; Zhong, Y; Naczk, M; Shahidi, F. Antioxidant activity and water holding capacity of canola protein hydrolysates. Food Chem 2008, 109, 144–148. [Google Scholar]
- Tang, C-H; Wang, X-S; Yang, X-Q. Enzymatic hydrolysis of hemp (Cannabis Sativa L.) protein isolate by various proteases and antioxidant properties of the resulting hydrolysates. Food Chem 2009, 114, 1484–1490. [Google Scholar]
- Udenigwe, CC; Lu, Y-L; Han, C-H; Hou, W-C; Aluko, RE. Flaxseed protein-derived peptide fractions: Antioxidant properties and inhibiton of lipopolysaccharide-induced nitric oxide production in murine macrophages. Food Chem 2009, 116, 277–284. [Google Scholar]
- Tang, C-H; Peng, J; Zhen, D-W; Chen, Z. Physiochemical and antioxidant properties of buckwheat (Fagopyrum esculentum Moench) protein hydrolysates. Food Chem 2009, 115, 672–678. [Google Scholar]
- Jamdar, SN; Rajalakshmi, V; Pednekar, MD; Juan, F; Yardi, F; Sharma, A. Influence of degree of hydrolysis on functional properties, antioxidant activity and ACE inhibitor activity of peanut protein hydrolysate. Food Chem 2010, 121, 178–184. [Google Scholar]
- Xie, Z; Haung, J; Xu, X; Jin, Z. Antioxidant activity of peptides isolated from alfafa leaf protein protein hydrolysate. Food Chem 2008, 111, 370–376. [Google Scholar]
- Liu, B-L; Chiang, P-S. Production of hydrolysate with antioxidant activity and function properties by enzymatic hydrolysis of defatted sesame (Sesamum indicum L.). J. Appl. Sci. Eng 2008, 6, 73–83. [Google Scholar]
- Zhang, SB; Wang, Z; Xu, SY; Gao, FX. Purification and characterization of a radical scavenging peptides from rapeseed protein hydrolysates. J. Am. Oil Chem. Soc 2009, 86, 959–966. [Google Scholar]
- Udenigwe, CC; Aluko, RE. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci 2011, 12, 3148–3161. [Google Scholar]
- Evans, JM; Boulter, D. Amino acid composition of seed meals of African yam bean (Sphenostylis stenocarpa) and Lima bean (Phaseolus lunatus). J. Sci. Food Agric 1999, 25, 919–922. [Google Scholar]
- Nwokolo, EA. The need to increase consumption of pulse in the developing world. In Food and Feed from Legumes and Oilseeds; Nwokolo, E, Smart, J, Eds.; Chapman and Hall: London, UK, 1996; pp. 3–11. [Google Scholar]
- Eromosele, CO; Arogundade, LA; Eromosele, IC; Ademuyiwa, O. Extractibility of African yam bean (Sphenostylis stenocarpa) protein in acid, salt and alkaline aqueous media. Food Hydrocolloid 2008, 22, 1622–1628. [Google Scholar]
- Evans, ID; Haismer, DR. Rheology of gelatinized starch suspension. J. Texture Stud 1979, 10, 347–370. [Google Scholar]
- Korhonen, H; Pihlanto, A. Food-derived bioactive peptides - opportunities for designing future foods. Curr. Pharm. Des 2003, 9, 1297–1308. [Google Scholar]
- Rajapakse, N; Mendis, E; Byun, HG; Kim, SK. Purification and in vitro antioxidative effects of giant squid muscle peptides on free radical-mediated oxidative systems. J. Nutr. Biochem 2005, 16, 562–569. [Google Scholar]
- Nam, KA; You, SG; Kim, SM. Molecular and physical characteristics of squid (Todarodes pacificus) skin collagens and biological properties of their enzymatic hydrolysates. J. Food Sci 2008, 73, C243–C255. [Google Scholar]
- Guo, H; Kouzuma, Y; Yonekura, M. Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem 2009, 113, 238–245. [Google Scholar]
- Aluko, RE; Monu, E. Functional and bioactive properties of quinoa seed protein hydrolysates. J. Food Sci 2003, 68, 1254–1258. [Google Scholar]
- Kim, SY; Je, J-Y; Kim, SK. Purification and characterization of antioxidant peptides form hoki (Johnius belengerii) frame protein by gastrointestinal digestion. J. Nutr. Biochem 2007, 81, 31–38. [Google Scholar]
- Elias, RJ; Kellerby, SS; Decker, EA. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr 2008, 48, 430–441. [Google Scholar]
- Gao, D; Cao, Y; Li, H. Antioxidant activity of peptide fractions derived from cottonseed protein hydrolysate. J. Sci. Food Agric 2010, 90, 1855–1860. [Google Scholar]
- Lee, J; Koo, N; Min, DB. Reactive oxygen species, aging, and antioxidant nutraceuticals. Compr. Rev. Food Sci. Food Saf 2004, 3, 21–33. [Google Scholar]
- Dong, S; Zeng, M; Wang, D; Liu, Z; Zhao, Y; Yang, H. Antioxidant and biochemical properties of protein hydrolysates prepared from silver carp (Hypophthalmichthys molitrix). Food Chem 2008, 107, 1485–1493. [Google Scholar]
- Saiga, A; Tanabe, S; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem 2003, 51, 3661–3667. [Google Scholar]
- Wang, H; Gao, XD; Zhou, GC; Cai, L; Yao, WB. In vitro and in vivo antioxidant activity of aqueous extract from Choerospondias axillaries fruit. Food Chem 2008, 106, 888–895. [Google Scholar]
- Bougatef, A; Hajji, M; Balti, R; Lassoued, I; Triki-Ellouz, Y; Nasri, M. Antioxidant and free radical-scavenging activities of smooth hound (Mustelus mustelus) muscle protein hydrolysates obtained by gastrointestinal proteases. Food Chem 2009, 114, 1198–1205. [Google Scholar]
- Qian, ZJ; Jung, WK; Kim, SK. Free radical scavenging activity of novel antioxidative peptide purified from hydrolysate of bullfrog skin (Rana catesbeiana shaw). Bioresource Technol 2008, 99, 1690–1698. [Google Scholar]
- You, L; Zhao, M; Cui, C; Zhao, H; Yang, B. Effect of degree of hydrolysis on the antioxidant activity of loach (Misgurnus anguillicaudatus) protein hydrolysates. Innov. Food Sci. Emerg. Technol 2009, 10, 235–240. [Google Scholar]
- Jayaprakasha, GK; Singh, RP; Sakariah, KK. Antioxidant activity of grape seed (Viti vinifera) extracts on peroxidation models in vitro. Food Chem 2001, 73, 285–290. [Google Scholar]
- Chen, G; Zhao, L; Zhao, L; Cong, T; Bao, S. In vitro study on antioxidant activities of peanut protein hydrolysate. J. Sci. Food Agric 2007, 87, 357–362. [Google Scholar]
- Fasakin, CF; Udenigwe, CC; Aluko, RE. Antioxidant properties of chlorophyll-enriched and chlorophyll-depleted polyphenolic fractions from leaves of Vernonia amygdalina and Gongronema latifolium. Food Res. Int 2011, 44, 2435–2441. [Google Scholar]
- Duan, S; Weng, XC; Dong, XW; Liu, YP; Li, HP; Jin, JR. Antioxidant properties of butylatedhydroxytoluene refluxed in ferric chloride solution. Food Chem 1998, 61, 101–105. [Google Scholar]
- Zhang, SB; Wang, Z; Xu, SY. Antioxidant and antithrombotic activities of rapeseed peptides. J Am Oil Chem Soc 2008, 85, 521–527. [Google Scholar]
- Adebowale, YA; Henle, T; Schwarzenbolz, U. Acetylated and succinylated derivatives of African yam bean (Sphenostylis sternocarpa) protein isolates. J. Mob. Commun 2009, 3, 34–46. [Google Scholar]
- Markwell, MAC; Haas, SM; Biebar, LL; Tolbert, NE. A modification of the Lowry procedure to simplify protein determination in membrane and in protein samples. Anal. Biochem 1978, 87, 206–211. [Google Scholar]
- Omoni, A; Aluko, RE. Mechanism of the inhibition of calmodulin-dependent neuronal nitric oxide synthase by flaxseed protein hydrolysates. J. Am. Oil Chem. Soc 2006, 83, 335–340. [Google Scholar]
- Bidlingmeyer, B; Cohen, S; Tarvin, T. Rapid analysis of amino acids using pre-column derivatization. J. Chromatogr 1984, 336, 93–104. [Google Scholar]
- Gehrke, C; Wall, L; Absheer, J; Kaiser, F; Zumwalt, R. Sample preparation for chromatography of amino acids: acid hydrolysis of proteins. Anal. Chem 1985, 68, 811–821. [Google Scholar]
- Landry, J; Delhaye, S. Simplified procedure for the determination of tryptophan of foods and feedstuffs from barytic hydrolysis. J. Agric. Food Chem 1992, 40, 776–779. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| API | APH | <1 kDa | 1–3 kDa | 3–5 kDa | 5–10 kDa | |
|---|---|---|---|---|---|---|
| ASX | 11.23 | 11.38 | 9.60 | 11.03 | 11.50 | 11.50 |
| THR | 3.92 | 4.06 | 4.54 | 4.07 | 3.63 | 3.79 |
| SER | 6.82 | 6.93 | 8.05 | 6.84 | 6.41 | 6.32 |
| GLX | 12.47 | 13.27 | 11.34 | 13.05 | 13.50 | 13.88 |
| PRO | 1.36 | 1.44 | 0.85 | 1.36 | 1.49 | 1.55 |
| GLY | 9.05 | 8.96 | 8.76 | 8.77 | 8.86 | 8.81 |
| ALA | 7.21 | 7.37 | 9.17 | 7.63 | 7.15 | 7.06 |
| CYS | 1.10 | 0.84 | 0.27 | 0.47 | 0.59 | 0.62 |
| VAL | 4.91 | 4.97 | 5.15 | 5.40 | 5.35 | 5.52 |
| MET | 1.20 | 0.96 | 1.13 | 0.92 | 0.97 | 0.97 |
| ILE | 4.14 | 3.96 | 3.73 | 4.29 | 4.33 | 4.41 |
| LEU | 8.56 | 8.35 | 10.25 | 9.11 | 8.70 | 8.47 |
| TYR | 4.54 | 3.73 | 3.85 | 3.65 | 3.66 | 3.59 |
| PHE | 6.03 | 5.97 | 7.34 | 6.34 | 6.02 | 5.96 |
| HIS | 3.48 | 3.59 | 2.99 | 3.33 | 3.59 | 3.64 |
| LYS | 8.03 | 8.33 | 7.36 | 8.01 | 8.26 | 8.10 |
| ARG | 4.90 | 4.97 | 4.45 | 4.90 | 5.14 | 5.03 |
| TRP | 1.04 | 0.94 | 1.17 | 0.84 | 0.85 | 0.80 |
| HAA | 40.09 | 38.52 | 42.91 | 40.01 | 39.11 | 38.94 |
| PCAA | 16.42 | 16.88 | 14.80 | 16.23 | 16.99 | 16.77 |
| NCAA | 23.70 | 24.65 | 20.94 | 24.08 | 25.00 | 25.38 |
| AAA | 11.62 | 10.64 | 12.36 | 10.82 | 10.53 | 10.35 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ajibola, C.F.; Fashakin, J.B.; Fagbemi, T.N.; Aluko, R.E. Effect of Peptide Size on Antioxidant Properties of African Yam Bean Seed (Sphenostylis stenocarpa) Protein Hydrolysate Fractions. Int. J. Mol. Sci. 2011, 12, 6685-6702. https://doi.org/10.3390/ijms12106685
Ajibola CF, Fashakin JB, Fagbemi TN, Aluko RE. Effect of Peptide Size on Antioxidant Properties of African Yam Bean Seed (Sphenostylis stenocarpa) Protein Hydrolysate Fractions. International Journal of Molecular Sciences. 2011; 12(10):6685-6702. https://doi.org/10.3390/ijms12106685
Chicago/Turabian StyleAjibola, Comfort F., Joseph B. Fashakin, Tayo N. Fagbemi, and Rotimi E. Aluko. 2011. "Effect of Peptide Size on Antioxidant Properties of African Yam Bean Seed (Sphenostylis stenocarpa) Protein Hydrolysate Fractions" International Journal of Molecular Sciences 12, no. 10: 6685-6702. https://doi.org/10.3390/ijms12106685