Assessing Molecular Signature for Some Potential Date (Phoenix dactylifera L.) Cultivars from Saudi Arabia, Based on Chloroplast DNA Sequences rpoB and psbA-trnH

Abstract
:1. Introduction
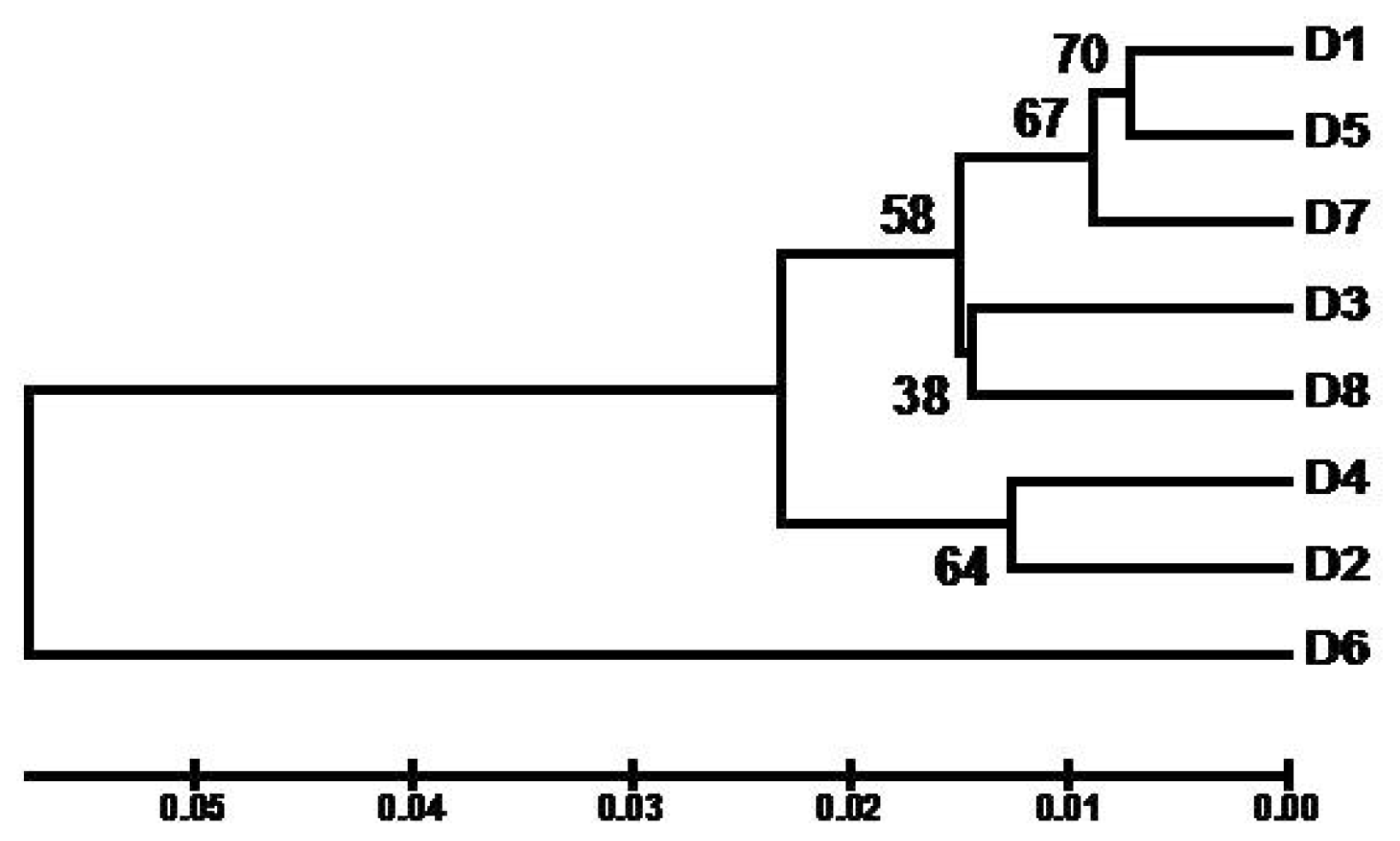
2. Results and Discussion
3. Materials and Methods
3.1. Sampling
3.2. Genomic DNA Extraction
3.2.1. Procedure
3.2.2. Amplification and Sequencing
3.3. DNA Sequence Data Analysis
4. Conclusion
Acknowledgement
References
- Sghaier-Hammami, B; Valledor, L; Drira, N; Jorrin-Novo, JV. Proteomic analysis of the development and germination of date palm (Phoenix dactylifera L.) zygotic embryos. Proteomics 2009, 9, 2543–2554. [Google Scholar]
- Anonymous. The fruit of the desert; Saudi Date Market, Al-Butain Agricultural Cooperative Association (BACA). issue June 7, 2004. Available online: http://www.dates.com.sa/SaudiDates/SaudiDates.htm accessed on 8 August 2011.
- Yang, M; Zhang, X; Liu, G; Yin, Y; Chen, K; Yun, Q; Zhao, D; Al-Mssallem, IS; Yu, J. The complete chloroplast genome sequence of date palm (Phoenix dactylifera L.). PLoS One 2010, 5, e12762. [Google Scholar]
- Saafi, EB; Trigui, M; Thabet, R; Hammami, M; Achour, L. Common date palm in Tunisia: Chemical composition of pulp and Pit. Int. J. Food Sci. Technol 2008, 43, 2033–2037. [Google Scholar]
- Elleuch, M; Besbes, S; Roiseux, O. Date flesh: Chemical composition and characteristics of the dietary fibre. Food Chem 2008, 111, 676–668. [Google Scholar]
- Biglari, F; AlKarkhi, AFM; Easa, AM. Cluster analysis of antioxidant compounds in dates (Phoenix dactylifera): Effect of long-term cold storage. Food Chem 2009, 112, 998–1001. [Google Scholar]
- Biglari, F; AlKarkhi, AFM; Easa, AM. Antioxidant activity and phenolic content of various date palm (Phoenix dactylifera) fruits from Iran. Food Chem 2008, 107, 1636–1641. [Google Scholar]
- Saafi, EB; El-Arem, A; Issaoui, M; Hammami, M; Achour, L. Phenolic content and antioxidant activity of four date palm (Phoenix dactylifera L.) fruit varieties grown in Tunisia. Int. J. Food Sci. Technol 2009, 44, 2314–2319. [Google Scholar]
- Mansouri, A; Embarek, G; Kokkalou, E; Kefalas, P. Phenolic profile and antioxidant activity of the Algerian ripe date palm fruit (Phoenix dactylifera). Food Chem 2005, 89, 411–420. [Google Scholar]
- Hong, YJ; Tomas-Barberan, FA; Kader, A; Mitchell, EA. The flavonoid glycosides and procyanidin composition of Deglet Noor Dates (Phoenix dactylifera). J. Agric. Food Chem 2006, 54, 2405–2411. [Google Scholar]
- Mrabet, A; Rejili, M; Lachiheb, B; Toivonen, P; Chaira, N; Ferchichi, A. Microbiological and chemical characterizations of organic and conventional date pastes (Phoenix dactylifera L.) from Tunisia. Ann. Microbiol 2008, 58, 453–459. [Google Scholar]
- Baaziz, M; Saaidi, M. Preliminary identification of date palm cultivars by esterase isoenzymes and peroxidase activities. Can. J. Bot 1988, 66, 89–93. [Google Scholar]
- Majourhat, K; Medraoui, KBL; Baaziz, M. Diversity of leaf peroxidases in date palm (Phoenix dactylifera L.) as revealed in an example of marginal (seedling derived) palm groves. Sci. Hort 2002, 95, 31–38. [Google Scholar]
- Ould Mohamed Salem, A; Trifi, M; Rhouma, A; Marrakchi, M. Genetic inheritance analyses of four enzymes in date palm (Phoenix dactylifera L.). Genet. Resour. Crop Evol 2001, 48, 361–368. [Google Scholar]
- Abdulla, M; Gamal, O. Investigation on molecular phylogeny of some date palm (Phoenix dactylifra L.) cultivars by protein, RAPD and ISSR markers in Saudi Arabia. Aust. J Crop Sci 2010, 4, 23–28. [Google Scholar]
- Aitchitt, M; Mantell, S; Thangavelu, M; Ainsworth, C. Cloning date palm (Phoenix dactylifera L.) DNA and characterization of low, medium and high copy DNA sequences. Elaeis 1995, 7, 57–63. [Google Scholar]
- Al-Khalifah, NS; Askari, E. Molecular phylogeny of date palm (Phoenix dactylifera L.) cultivars from Saudi Arabia by DNA fingerprinting. Theor. Appl. Genet 2003, 107, 1266–1270. [Google Scholar]
- Rawashdeh, I; Amri, A. Genetic characterization of date palm varieties using RAPD markers. Jordan J. Agri. Sci 2006, 2, 234–242. [Google Scholar]
- Soliman, SS; Ali, BA; Ahmed, MM. Genetic comparisons of Egyptian date palm cultivars (Phoenix dactylifera L.) by RAPD-PCR. Afr. J. Biotechnol 2003, 2, 86–87. [Google Scholar]
- Zehdi, S; Trifi, M; Billotte, N; Marakchi, M; Pintaud, JC. Genetic diversity of Tunisian date palms (Phoenix dactylifera L.) revealed by nuclear microsatellite polymorphism. Hereditas 2004, 141, 278–287. [Google Scholar]
- Elshibli, S; Korpelainen, H. Microsatellite markers reveal high genetic diversity in date palm (Phoenix dactylifera L.) germplasm from Sudan. Genetica 2008, 134, 251–260. [Google Scholar]
- Ali, MA; Choudhary, RK. India needs more plant taxonomists. Nature 2011, 471, 37. [Google Scholar]
- Nock, CJ; Waters, DLE; Edwards, MA; Bowen, SG; Rice, N; Cordeiro, GM; Henry, RJ. Chloroplast genome sequences from total DNA for plant identification. Plant Biotechnol. J 2011, 9, 328–333. [Google Scholar]
- Yao, H; Song, JY; Ma, XY; Liu, C; Li, Y; Xu, HX. Identification of Dendrobium species by a candidate DNA barcode sequence: The chloroplast psbA-trnH intergenic region. Planta Med 2009, 75, 667–669. [Google Scholar]
- Song, J; Yao, H; Li, Y; Li, X; Lin, Y; Liu, C. Authentication of the family Polygonaceae in Chinese pharmacopoeia by DNA barcoding technique. J. Ethnopharmacol 2009, 124, 434–439. [Google Scholar]
- Kress, WJ; Wurdack, KJ; Zimmer, EA; Weigt, LA; Janzen, DH. Use of DNA barcodes to identify flowering plants. Proc Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar]
- Dong, J; Wagner, DB; Yanchuk, AD; Carlson, MR; Magnussen, S; Wang, XR; Szmidt, AE. Paternal chloroplast DNA inheritance in Pinus consora and Pinus banksiana: independence of parenetal species or cross direction. J. Hered 1992, 83, 419–422. [Google Scholar]
- Mason, RJ; Holsinger, KE; Jansen, RK. Biparental inheritance of the chloroplast genome in Coreopsis (Asteraceae). J. Hered 1994, 85, 171–173. [Google Scholar]
- Hansen, AK; Escobar, LK; Gilbert, LE; Jansen, RK. Paternal, maternal, and biparental inheritance of the chloroplast genome in Passiflora (Passifloraceae): Implications for phylogenetic studies. Am. J. Bot 2007, 94, 42–46. [Google Scholar]
- Wikipedia. Arecaceae. Available online: http://en.wikipedia.org/wiki/Arecaceae accessed on 10 September 2011.
- Kress, WJ; Erickson, DL. A Two-locus global DNA barcode for land plants: The coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS One 2007, 6, e508. [Google Scholar]
- Goremykin, VV; Holland, B; Hirsch-Ernst, KI; Hellwig, FH. Analysis of Acorus calamus chloroplast genome and its phylogenetic implications. Mol. Phylogenet. Evol 2005, 22, 1813–1822. [Google Scholar]
- Hamrick, JL; Godt, MJW. Allozyme diversity in plant species. In Plant Population Genetics, Breeding and Genetic Resources; Brown, AHD, Clegg, MT, Kahler, AL, Weir, BS, Eds.; Sinauer Associates Inc: Sunderland, UK, 1989. [Google Scholar]
- Nybom, H; Bartish, IV. Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. Perspect. Plant Ecol. Evol. Syst 2000, 3, 93–114. [Google Scholar]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intra-specific genetic diversity in plants. Mol. Ecol 2004, 13, 1143–1155. [Google Scholar]
- Fakir, S; Car Munier, P. Origine de la culture sur palmier dattier et sa propagation en Afrique. Fruits 1981, 36, 437–450. [Google Scholar]
- Sneath, PHA; Sokal, RR. Numerical Taxonomy; WH Freeman & Company: San Francisco, CA, USA, 1973. [Google Scholar]
- Sedra, MH; Lashermes, P; Trouslot, P; Combes, M; Hamon, S. Identification and genetic diversity analysis of date palm (Phoenix dactylifera L.) varieties from Morocco using RAPD markers. Euphytica 1998, 103, 75–82. [Google Scholar]
- Cao, BR; Chao, CT. Identification of date cultivars in California using AFLP. Hort. Sci 2002, 37, 966–968. [Google Scholar]
- Zehdi, S; Sakka, H; Rhouma, A; Ould Mohamed Salem, A; Marrakchi, M; Trifi, M. Analysis of Tunisian date palm germplasm using simple sequence repeats. Afr. J. Biotechnol 2004, 3, 215–519. [Google Scholar]
- Bennaceur, M; Lanaud, C; Chevalier, MH; Bounaga, N. Genetic diversity of the date (Phoenix dactylifera L.) from Algeria revealed by enzyme markers. Plant Breed 1991, 107, 56–69. [Google Scholar]
- Pennisi, E. Taxonomy. Wanted: A barcode for plants. Science 2007, 318, 190–191. [Google Scholar]
- Sass, C; Little, DP; Stevenson, DW; Specht, CD. DNA barcoding in the Cycadales: Testing the potential of proposed barcoding markers for species identification of Cycads. PLoS One 2007, 11, e1154. [Google Scholar]
- Newmaster, SG; Fazekas, AJ; Steeves, RAD; Janovec, J. Testing candidate plant barcode regions in the Myristicaceae. Mol. Ecol. Notes 2008, 8, 480–490. [Google Scholar]
- Taberlet, P; Coissac, E; Pompanon, F; Gielly, L; Miquel, C; Valentini, A; Vermat, T; Corthier, G; Brochmann, C; Willerslev, E. Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. Nucleic Acids Res 2007, 35, e14. [Google Scholar]
- Lahaye, R; van der Bank, M; Bogarin, D; Warner, J; Pupulin, F; Gigot, G; Maurin, O; Duthoit, S; Barraclough, TG; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar]
- Feng, T; Liu, S; He, XJ. Molecular authentication of the traditional Chinese medicinal plant Angelica sinensis based on internal transcribed spacer of nrDNA. Electron. J. Biotechnol 2010, 13, 1–10. [Google Scholar]
- Chen, SL; Yao, H; Han, JP; Liu, C; Song, JY; Shi, LC; Zhu, YJ; Ma, XY; Gao, T; Pang, XH; et al. Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS One 2010, 5, e8613. [Google Scholar]
- Khan, S; Qureshi, MI; Kamaluddin; Alam, T; Abdin, MZ. Protocol for isolation of genomic DNA from dry and fresh roots of medicinal plants suitable for RAPD and restriction digestion. Afr. J. Biotechnol 2007, 6, 175–178. [Google Scholar]
- Thompson, JD; Gibson, TJ; Plewniak, F; Jeanmougin, F; Higgins, DG. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997, 24, 4876–4882. [Google Scholar]
- Hall, TA. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acid Symp. Ser 1999, 41, 95–98. [Google Scholar]
- Rozas, J; Rozas, R. DnaSP, DNA sequence polymorphism: An interactive program for estimating population genetics parameters from DNA sequence data. Comput. Appl. Biosci 1995, 11, 621–625. [Google Scholar]
- Tamura, K; Dudley, J; Nei, M; Kumar, S. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol 2007, 24, 1596–1599. [Google Scholar]
- Tamura, K; Nei, M; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar]


{kind=link}
| Cultivars | Abbreviation | Sequence characteristics | |||||
|---|---|---|---|---|---|---|---|
| rpoB | psbA-trnH | ||||||
| Accession No. | Total length | %GC | Accession No. | Total length | %GC | ||
| Khodry | D1 | JN854236 | 463 | 39 | JN854228 | 674 | 28 |
| Khalas | D2 | JN854237 | 464 | 40 | JN854229 | 674 | 29 |
| Ruthana | D3 | JN854238 | 463 | 39 | JN854230 | 674 | 29 |
| Sukkari | D4 | JN854239 | 469 | 41 | JN854231 | 674 | 29 |
| Sefri | D5 | JN854240 | 464 | 39 | JN854232 | 674 | 29 |
| Segae | D6 | JN854241 | 467 | 40 | JN854233 | 674 | 32 |
| Ajwa | D7 | JN854242 | 463 | 39 | JN854234 | 674 | 29 |
| Hilali | D8 | JN854243 | 463 | 39 | JN854235 | 674 | 29 |
| Sequence polymorphism | psbA-trnH | rpoB |
|---|---|---|
| Number of polymorphic sites | 135 | 33 |
| Variance of haplotype diversity | 0.00391 | 0.0339 |
| Nucleotide diversity (Pi) | 0.0686 | 0.0276 |
| Theta (per site) from Eta | 0.0807 | 0.0276 |
| Average number of nucleotide differences (K) | 45.893 | 11.321 |
| Gene | Primer | Primer sequence 5′-3′ | Reaction conditions |
|---|---|---|---|
| psbA-trnH | Forward | GTTATGCATGAACGTAATGCTC | 94 °C 1 min, 94 °C 30 s, 53 °C 40 s, 72 °C 30 s, 40 cycles, 72 °C 5 min |
| Reverse | CGCGCATGGTGGATTCACAAATC | ||
| rpoB | Forward | ATGCAACGTCAAGCAGTTCC | 94 °C 1 min, 94 °C 30 s, 53 °C 40 s, 72 °C 30 s, 40 cycles, 72 °C 5 min |
| Reverse | GATCCCAGCATCACAATTCC |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Al-Qurainy, F.; Khan, S.; Al-Hemaid, F.M.; Ali, M.A.; Tarroum, M.; Ashraf, M. Assessing Molecular Signature for Some Potential Date (Phoenix dactylifera L.) Cultivars from Saudi Arabia, Based on Chloroplast DNA Sequences rpoB and psbA-trnH. Int. J. Mol. Sci. 2011, 12, 6871-6880. https://doi.org/10.3390/ijms12106871
Al-Qurainy F, Khan S, Al-Hemaid FM, Ali MA, Tarroum M, Ashraf M. Assessing Molecular Signature for Some Potential Date (Phoenix dactylifera L.) Cultivars from Saudi Arabia, Based on Chloroplast DNA Sequences rpoB and psbA-trnH. International Journal of Molecular Sciences. 2011; 12(10):6871-6880. https://doi.org/10.3390/ijms12106871
Chicago/Turabian StyleAl-Qurainy, Fahad, Salim Khan, Fahad M. Al-Hemaid, M. Ajmal Ali, M. Tarroum, and M. Ashraf. 2011. "Assessing Molecular Signature for Some Potential Date (Phoenix dactylifera L.) Cultivars from Saudi Arabia, Based on Chloroplast DNA Sequences rpoB and psbA-trnH" International Journal of Molecular Sciences 12, no. 10: 6871-6880. https://doi.org/10.3390/ijms12106871