Increase in Docetaxel-Resistance of Ovarian Carcinoma-Derived RMG-1 Cells with Enhanced Expression of Lewis Y Antigen

Abstract
:1. Introduction
2. Results
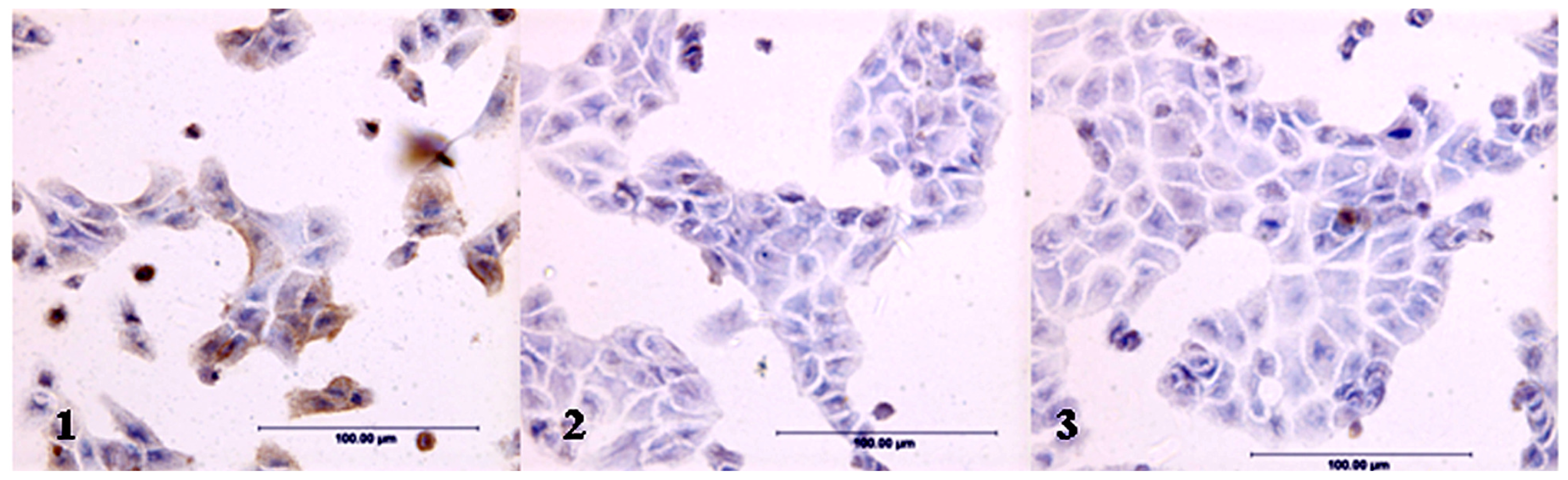
2.1. Expression of LeY Antigen in Cells Transfected with the hFUT Gene
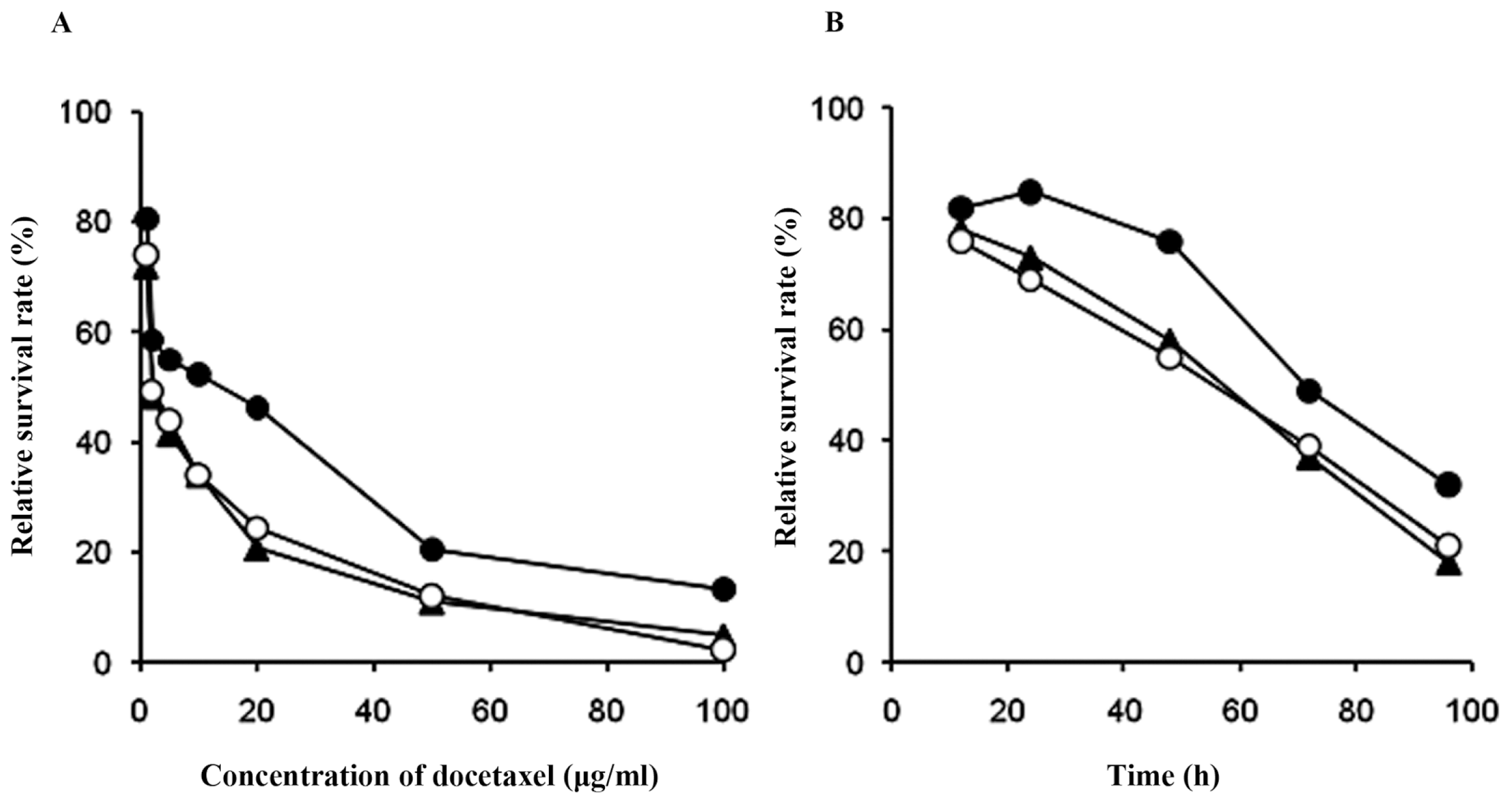
2.2. Effects of Docetaxel on the Survival Rates of Cells
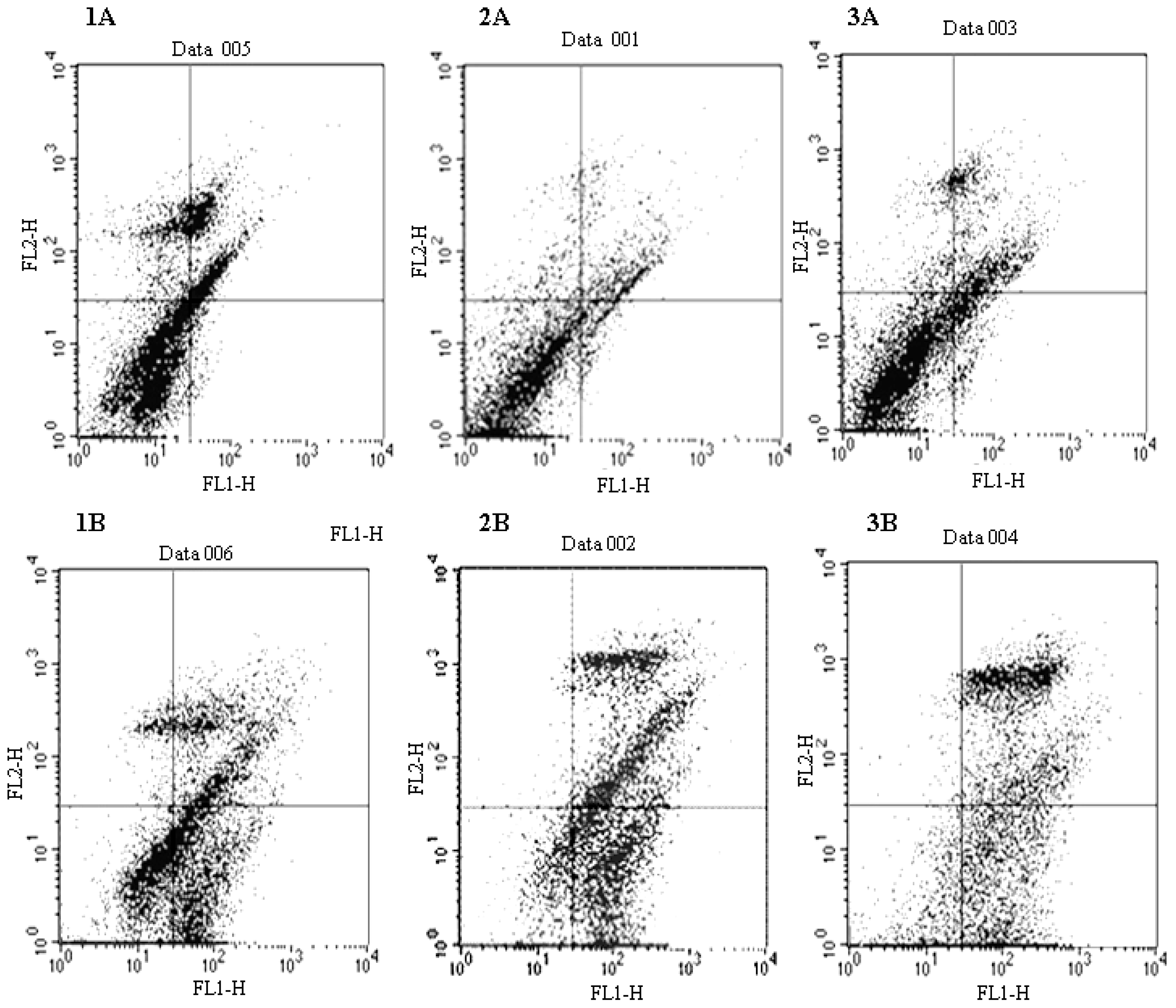
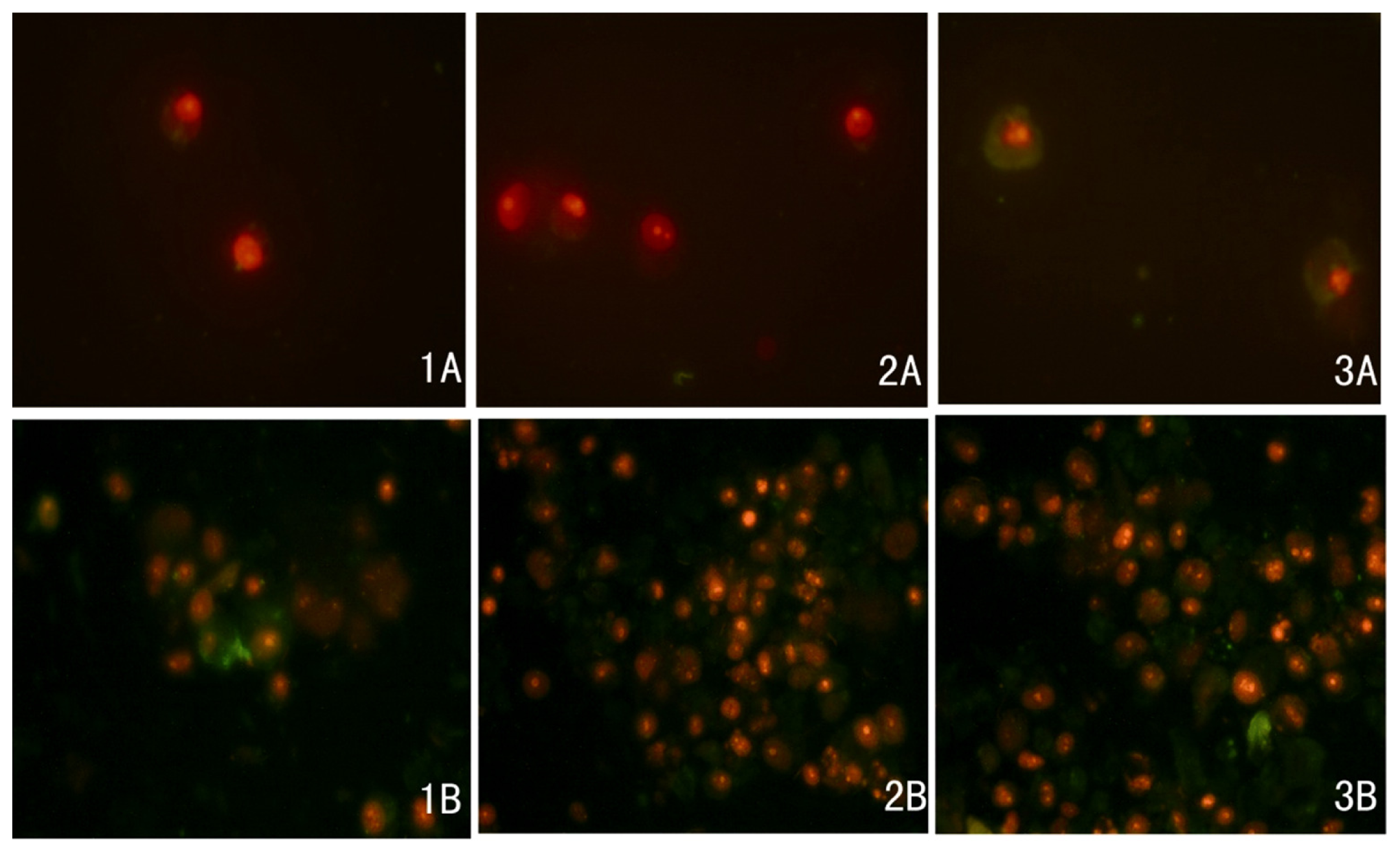
2.3. Docetaxel-Induced Apoptosis
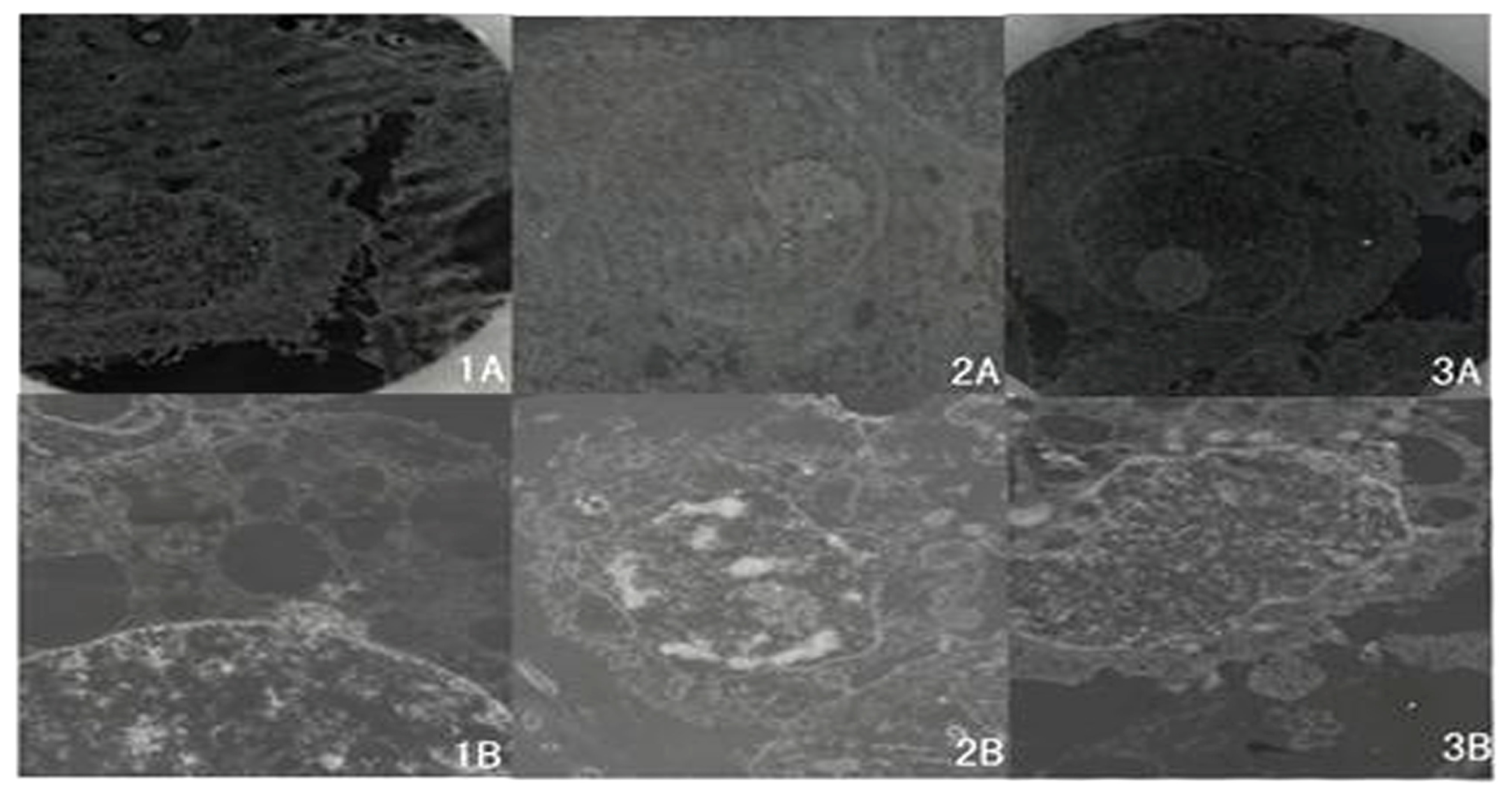
2.4. Ultrastructure of Cells Examined by TEM
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Transfection of the Fucosyltransferase Gene
4.4. Immunocytochemical Assaying
4.5. Cell Viability Determined by MTT Assaying
4.6. Flow Cytometry
4.7. Transmission Electron Microscopy (TEM)
5. Conclusions
Acknowledgments
References
- Huang, Y.; Sadée, W. Membrane transporters and channels in chemoresistance and-sensitivity of tumor cells. Cancer Lett 2006, 239, 168–182. [Google Scholar]
- Mahadevan, D.; Shirahatti, N. Strategies for targeting the multidrug resistance-1 (MDR1)/P-gp transporter in human malignancies. Curr. Cancer Drug Targets 2005, 5, 445–455. [Google Scholar]
- Lavie, Y.; Cao, H.; Bursten, S.L.; Giuliano, A.E.; Cabot, M.C. Accumulation of glucosylceramides in multidrug-resistant cancer cells. J. Biol. Chem 1996, 271, 19530–19536. [Google Scholar]
- Lucci, A.; Cho, W.I.; Han, T.Y.; Giuliano, A.E.; Morton, D.L.; Cabot, M.C. Glucosylceramide: A marker for multiple-drug resistant cancers. Anticancer Res 1998, 18, 475–480. [Google Scholar]
- Kiguchi, K.; Iwamori, Y.; Suzuki, N.; Kobayashi, Y.; Ishizuka, B.; Ishiwata, I.; Kita, T.; Kikuchi, Y.; Iwamori, M. Characteristic expression of globotriaosyl ceramide in human ovarian carcinoma-derived cells with anticancer drug resistance. Cancer Sci 2006, 97, 1321–1326. [Google Scholar]
- Iwamori, M.; Iwamori, Y.; Kubushiro, K.; Ishiwata, I.; Kiguchi, K. Characteristic expression of Lewis-antigenic glycolipids in human ovarian carcinoma-derived cells with anticancer drug-resistance. J. Biochem 2007, 141, 309–317. [Google Scholar]
- Iwamori, M.; Tanaka, K.; Kubushiro, K.; Lin, B.; Kiguchi, K.; Ishiwata, I.; Tsukazaki, K.; Nozawa, S. Alterations in the glyolipid composition and cellular properties of ovarian carcinoma-derived RMG-1 cells on transfection of the α,1,2-fucosyltransferase gene. Cancer Sci 2005, 96, 26–30. [Google Scholar]
- Zhao, Y.; Lin, B.; Hao, Y.Y.; Yan, L.M.; Liu, J.J.; Zhu, L.C.; Zhang, S.L. Effects of Lewis (Y) changes in abundances on ovarian cancer cell, RMG-I strain carboplatin resistance. Prog. Biochem. Biophys 2008, 35, 1175–1182. [Google Scholar]
- Bevilacqua, M.P.; Stengelin, S.; Gimbrone, M.A., Jr; Seed, B. Endothelial leukocyte adhesion molecule 1: An inducible receptor for neutrophils related to complement regulatory proteins and lectins. Science 1989, 243, 1160–1165. [Google Scholar]
- Ohmori, K.; Kanda, K.; Mitsuoka, C.; Kanamori, A.; Kurata-Miura, K.; Sasaki, K.; Nishi, T.; Tamatani, T.; Kannagi, R. P- and E-selectins recognize sialyl 6-sulfo Lewis X, the recently identified l-selectin ligand. Biochem. Biophys. Res. Commun 2000, 278, 90–96. [Google Scholar]
- Goupille, C.; Marionneau, S.; Bureau, V.; Hallouin, F.; Meichenin, M.; Rocher, J.; Le, P.J. Alpha1,2-fucosyltransferase increases resistance to apoptosis of rat colon carcinoma cells. Glycobiology 2000, 10, 375–382. [Google Scholar]
- Baldus, S.E.; Hanisch, F.G.; Pütz, C.; Flucke, U.; Mönig, S.P.; Schneider, P.M.; Thiele, J.; Hölscher, A.H.; Dienes, H.P. Immunoreactivity of Lewis blood group and mucin peptide core antigens: Correlations with grade of dysplasia and malignant transformation in the colorectal adenoma-carcinoma sequence. Histol. Histopathol 2002, 17, 191–198. [Google Scholar]
- Chhieng, D.C.; Rodriguez-Burford, C.; Talley, L.I.; Sviglin, H.; Stockard, C.R.; Kleinberg, M.J.; Barnes, M.N.; Partridge, E.E.; Khazaeli, M.B.; Grizzle, W.E. Expression of CEA, Tag-72, and Lewis-Y antigen in primary and metastatic lesions of ovarian carcinoma. Hum. Pathol 2003, 34, 1016–1021. [Google Scholar]
- Zhu, K.; Amin, M.A.; Zha, Y.; Harlow, L.A.; Koch, A.E. Mechanism by which H-2g, a glucose analog of blood group H antigen, mediates angiogenesis. Blood 2005, 105, 2343–2349. [Google Scholar]
- Sabbatini, P.J.; Ragupathi, G.; Hood, C.; Aghajanian, C.A.; Juretzka, M.; Iasonos, A.; Hensley, M.L.; Spassova, M.K.; Ouerfelli, O.; Spriggs, DR. Pilot study of a heptavalent vaccine-keyhole limpet hemocyanin conjugate plus QS21 in patients with epithelial ovarian fallopian tube or peritoneal cancer. Clin. Cancer Res 2007, 13, 4170–4177. [Google Scholar]
- Cordel, S.; Goupille, C.; Hallouin, F.; Meflah, K.; Le Pendu, J. Role for alpha1,2-fucosyltransferase and histo-blood group antigen H type 2 in resistance of rat colon carcinoma cells to 5-fluorouracil. Int. J. Cancer 2000, 85, 142–148. [Google Scholar]
- Yazawa, S.; Nishimura, T.; Ide, M.; Asao, T.; Okamura, A.; Tanaka, S.; Takai, I.; Yagihashi, Y.; Saniabadi, A.R.; Kochibe, N. Tumor-related expression of alpha1,2-fucosylated antigens on colorectal carcinoma cells and its suppression by cell-mediated priming using sugar acceptors for alpha1,2-fucosyltransferase. Glycobiology 2002, 12, 545–553. [Google Scholar]
- Basu, A.; Murthy, U.; Rodeck, U.; Herlyn, M.; Mattes, L.; Das, M. Presence of tumor-associated antigens in epidermal growth factor receptors from different human carcinomas. Cancer Res 1987, 47, 2531–2536. [Google Scholar]
- Klinger, M.; Farhan, H.; Just, H.; Drobny, H.; Himmler, G.; Loibner, H.; Mudde, G.C.; Freissmuth, M.; Sexl, V. Antibodies directed against Lewis-Y antigen inhibit signaling of Lewis-Y modified ErbB receptors. Cancer Res 2004, 64, 1087–1093. [Google Scholar]
- Garcia, R.; Franklin, R.A.; McCubrey, J.A. EGF induces cell motility and muti-drug resistance gene expression in breast cancer cells. Cell Cycle 2006, 5, 2820–2826. [Google Scholar]
- Venugopal, J.; Hanashiro, K.; Yang, Z.Z.; Nagamine, Y. Identification and modulation of a caveolae-dependendent signal pathway that regulates plasminogen activator inhibitor-1 in insulin-resistant adipocytes. Proc. Natl. Acad. Sci. USA 2004, 101, 17120–17125. [Google Scholar]
- Lee, H.H.; Chiang, W.H.; Chiang, S.H.; Liu, Y.C.; Hwang, J.; Ng, S.Y. Regulation of cyclin D1, DNA topoiso-merase I, and proliferating cell nuclear antigen promoters during the cell cycle. Gene Expr 1995, 4, 95–109. [Google Scholar]
- Ying, T.H.; Hsieh, Y.H.; Hsieh, Y.S.; Liu, J.Y. Antisense oligonucleotide Elk-1 suppresses the tumorigenicity of human hepatocellular carcinoma cells. Cell Biol. Int 2008, 32, 210–216. [Google Scholar]
- Wang, H.; Wang, Q.Y.; Zhang, Y.; Shen, Z.H.; Chen, H.L. Alpha1,3 Fucosyltransferase-VII modifies the susceptibility of apoptosis induced by ultraviolet and retinoic acid in human hepatocarcinoma cells. Glycoconj. J 2007, 24, 207–220. [Google Scholar]
- Zhu, L.C.; Lin, B.; Hao, Y.Y.; Li, F.F.; Diao, B.; Zhang, S.L. Impact of alpha1,2-fucosyltransferase gene transfection on cancer related gene expression profile of human ovarian cancer cell line RMG-I. Ai Zheng 2008, 27, 934–941. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell line | IC50 (μg/mL) | ta | Pa | tb | Pb |
|---|---|---|---|---|---|
| RMG-1-hFUT | 7.98 ± 1.31 | 2.88 | 0.05 | 3.34 | 0.03 |
| RMG-1(-) | 4.60 ± 1.18 | 0.16 | 0.88 | ||
| RMG-1 | 4.76 ± 1.42 |
| Cell line | Apoptotic cells (%) | ta | Pa | tb | Pb |
|---|---|---|---|---|---|
| RMG-1-hFUT | 65.3 ± 2.1 | 7.42 | 0.006 | 9.50 | 0.002 |
| RMG-1(-) | 83.7 ± 5.5 | 0.14 | 0.71 | ||
| RMG-1 | 82.2 ± 6.4 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, F.; Liu, J.; Lin, B.; Liu, Q.; Zhao, Y.; Zhu, L.; Hao, Y.; Zhang, S.; Iwamori, M. Increase in Docetaxel-Resistance of Ovarian Carcinoma-Derived RMG-1 Cells with Enhanced Expression of Lewis Y Antigen. Int. J. Mol. Sci. 2011, 12, 7323-7334. https://doi.org/10.3390/ijms12117323
Zhang F, Liu J, Lin B, Liu Q, Zhao Y, Zhu L, Hao Y, Zhang S, Iwamori M. Increase in Docetaxel-Resistance of Ovarian Carcinoma-Derived RMG-1 Cells with Enhanced Expression of Lewis Y Antigen. International Journal of Molecular Sciences. 2011; 12(11):7323-7334. https://doi.org/10.3390/ijms12117323
Chicago/Turabian StyleZhang, Fan, Juanjuan Liu, Bei Lin, Qing Liu, Yue Zhao, Liancheng Zhu, Yingying Hao, Shulan Zhang, and Masao Iwamori. 2011. "Increase in Docetaxel-Resistance of Ovarian Carcinoma-Derived RMG-1 Cells with Enhanced Expression of Lewis Y Antigen" International Journal of Molecular Sciences 12, no. 11: 7323-7334. https://doi.org/10.3390/ijms12117323