Effects of Conjugated Linoleic Acid, Fish Oil and Soybean Oil on PPARs (α & γ) mRNA Expression in Broiler Chickens and Their Relation to Body Fat Deposits

Abstract
:1. Introduction
2. Results and Discussion
2.1. Liver Fat Content and Abdominal Fat Pad Deposition
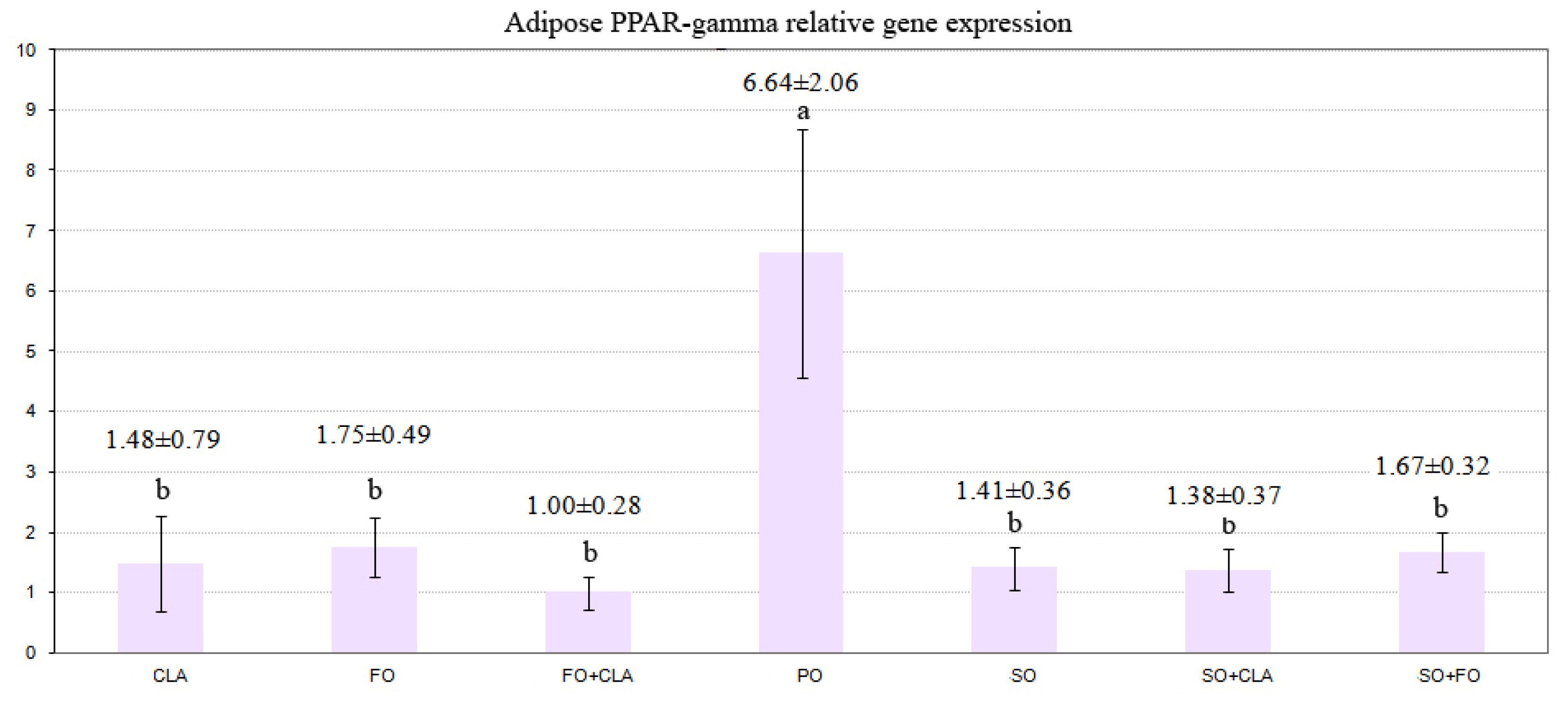
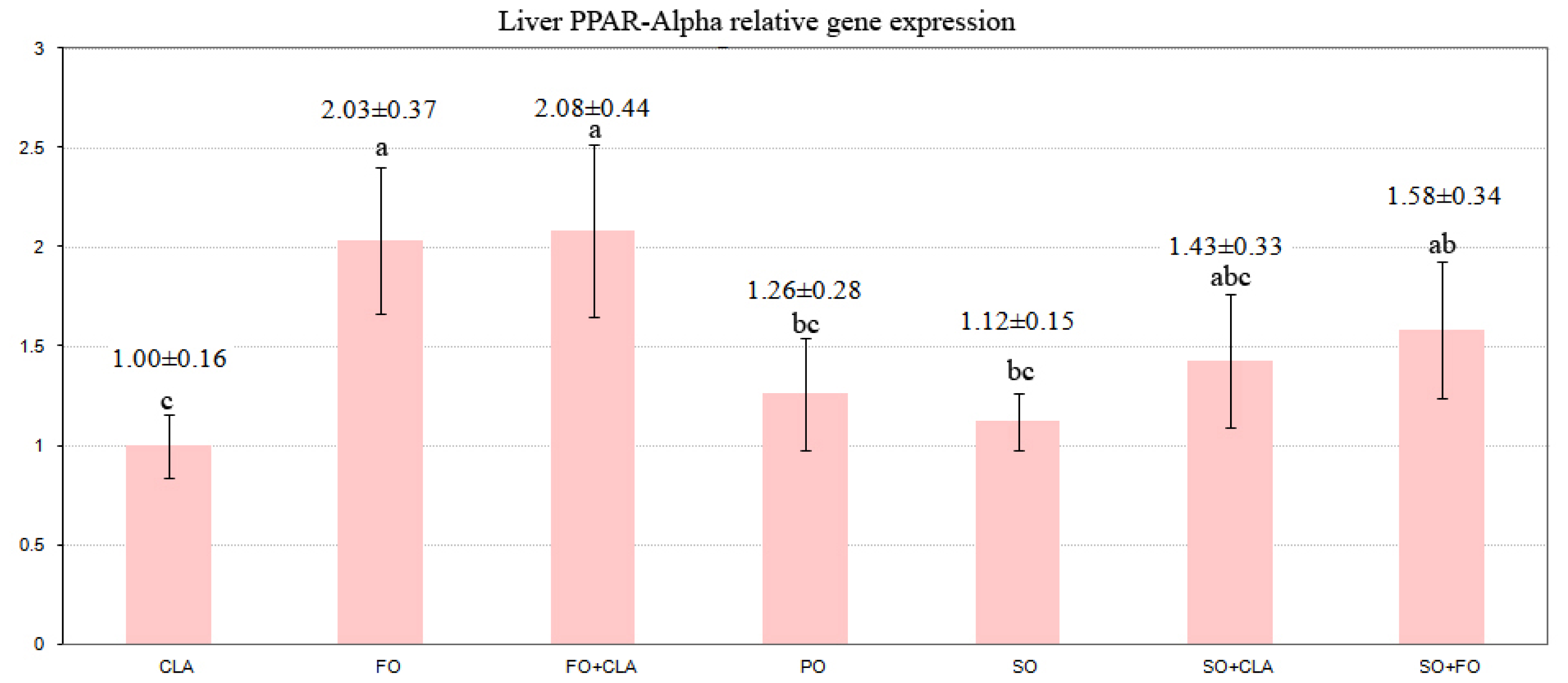
2.2. PPARs Gene Expression
3. Experimental Section
Statistical Analyses
4. Conclusions
Acknowledgments
References
- Scanes, C.G. The physiology of growth, growth hormone and growth factors in poultry. CRC Crit. Rev. Poult. Biol 1987, 1, 51–105. [Google Scholar]
- Furuse, M.; Murai, A.; Kita, K.; Asakura, K.; Okumura, J.I. Lipogenesis depending on sexual maturity in female Japanese quail (Coturnix coturnix Japonica). Comp. Biochem. Physiol. Part B 1991, 100, 343–345. [Google Scholar]
- Griminger, P. Lipid Metabolism. In Avian Physiology, 4th ed.; Sturkie, P.D., Ed.; Springer-Verlag: New York, NY, USA, 1986; pp. 345–358. [Google Scholar]
- Pearce, J. A comparison of the response of hepatic enzyme activity in domestic fowl (Gallus domesticus) and the rat (Rattus norvegicus) to the feeding of diets containing large proportions of glucose and fructose. Comp. Biochem. Physiol 1980, 651B, 423–426. [Google Scholar]
- Nir, I.; Lin, H. The skeleton, an important site of lipogenesis in the chick. Ann. Nutr. Metab 1982, 26, 100–105. [Google Scholar]
- Wilson, M.D.; Hays, R.D.; Clarke, S.D. Inhibition of liver lipogenesis by dietary polyunsaturated fat in severely diabetic rats. J. Nutr 1986, 116, 1511–1518. [Google Scholar]
- Wilson, M.D.; Blake, W.L.; Salati, L.M.; Clarke, S.D. Potency of polyunsaturated and saturated fats as short-term inhibitors of hepatic lipogenesis in rats. J. Nutr 1990, 120, 544–552. [Google Scholar]
- Blake, W.L.; Clarke, S.D. Suppression of hepatic fatty acid synthase and S14 gene transcription by dietary polyunsaturated fat. J. Nutr 1990, 120, 225–231. [Google Scholar]
- Ntambi, J.M. Dietary regulation of stearoyl-CoA desaturase I gene expression in mouse liver. J. Biol. Chem 1991, 267, 10925–10930. [Google Scholar]
- Sanz, M.; Lopez-Bote, C.J.; Menoyo, D.; Bautista, J.M. Abdominal fat deposition and fatty acid synthesis are lower and β-oxidation is higher in broiler chickens fed diets containing unsaturated rather than saturated fat. J. Nutr 2000, 130, 3034–3037. [Google Scholar]
- Shimomura, Y.; Tamura, T.; Suzuki, M. Less body fat accumulation in rats fed a safflower oil diet than in rats fed a beef tallow diet. J. Nutr 1990, 120, 1291–1296. [Google Scholar]
- Madsen, L.; Rustan, A.C.; Vaagenes, H.; Berge, K.; Dyroy, H.; Berge, R.K. Eicosapentaenoic and docosahexaenoic acid affect mitochondrial and peroxisomal fatty acid oxidation in relation to substrate preference. Lipids 1999, 34, 951–963. [Google Scholar]
- Takeuchi, H.; Matsuo, T.; Tokuyama, K.; Shimomura, Y.; Suziki, M. Diet-induced thermogenesis is lower in rats fed a lard diet than in those fed a high oleic acid safflower oil diet, a safflower oil diet of a linseed oil diet. J. Nutr 1995, 125, 920–925. [Google Scholar]
- Rustan, A.C.; Nossen, J.O.; Christiansen, E.N.; Drevon, C.A. Eicosapentaenoic acid reduces hepatic synthesis and secretion of triacylglycerol by decreasing the activity of acylcoenzyme A: 1.2-diacylglycerotrlansferase. J. Lipid Res 1988, 29, 1417–1426. [Google Scholar]
- Simon, O.; Manner, K.; Schafer, K.; Sagredos, A.; Eder, K. Effects of conjugated linoleic acids on protein to fat proportions, fatty acids, and plasma lipids in broilers. Eur. J. Lipid Sci. Technol 2000, 102, 402–410. [Google Scholar]
- Szymczyk, B.; Pisulewski, P.M.; Szczurek, W.; Hanczakowski, P. Effects of conjugated linoleic acid on growth performance, feed conversion efficiency, and subsequent carcass quality in broiler chickens. Br. J. Nutr 2001, 85, 465–473. [Google Scholar]
- Pariza, M.W.; Park, Y.; Cook, M.E. The biologically active isomers of conjugated linoleic acid. Prog. Lipid Res 2001, 40, 283–289. [Google Scholar]
- Wahle, K.W.J.; Heys, S.D.; Rotondo, D. Conjugated linoleic acids: are the beneficial or detrimental to health? Prog. Lipid Res 2004, 43, 553–587. [Google Scholar]
- Bouthegourd, J.C.; Evans, P.C.; Gripois, D.; Tiffon, B.; Blouguit, M.E.; Roseau, S.; Lutton, C.; Tombe, D.; Martin, J.C. A CLA mixture prevents body triglyceride accumulation without affecting energy expenditure in Syrian hamsters. J. Nutr 2002, 132, 2682–2689. [Google Scholar]
- Degrace, P.; Demizieux, L.; Gresti, J.; Chardigny, J.M.; Sébédio, J.L.; Clouet, P. Hepatic steatosis is not due to impaired fatty acid oxidation capacities in C57BL/6J mice fed the conjugates trans-10, cis-12-isomer of linoleic acid. J. Nutr 2004, 134, 861–867. [Google Scholar]
- Park, Y.; Albright, K.J.; Storkson, J.M.; Cook, M.E.; Pariza, M.W. Effect of conjugated linoleic acid on body composition in mice. Lipids 1997, 32, 853–858. [Google Scholar]
- Peters, J.M.; Park, Y.; Gonzalez, F.J.; Pariza, M.W. Influence of conjugated linoleic acid on body composition and target gene expression in peroxisome-proliferator-activated receptor α-null mice. Biochim. Biophys. Acta 2001, 1533, 233–241. [Google Scholar]
- Allman, D.W.; Gibson, D.W. Fatty acid synthesis during early linoleic acid deficiency in the mouse. J. Lipid Res 1969, 6, 51–62. [Google Scholar]
- Clarke, S.D.; Jump, D.B. Dietary polyunsaturated fatty acid regulation of gene transcription. Ann. Rev. Nutr 1994, 14, 83–98. [Google Scholar]
- Mikkelsen, L.; Hansen, H.S.; Grunnet, N.; Dich, J. Inhibition of fatty acid synthesis in rat hepatocytes by exogenous polyunsaturated fatty acids is caused by lipid peroxidation. Biochim. Biophys. Acta 1993, 1166, 99–100. [Google Scholar]
- Wahle, K.W.; Rotondo, D.; Heys, S.D. Polyunsaturated fatty acids and gene expression in mammalian systems. Proc. Nutr. Soc 2003, 62, 349–360. [Google Scholar]
- Wahli, W.; Braissant, O.; Desvergne, B. Peroxisome proliferator activated receptors: Transcriptional regulators of adipogenesis, lipid metabolism and more. Chem. Biol 1995, 2, 261–266. [Google Scholar]
- Xu, H.E.; Lambert, M.H.; Montana, V.G.; Parks, D.J.; Blanchard, S.G.; Brown, P.J.; Sternbach, D.D.; Lehmann, J.M.; Wisely, G.B.; Willson, T.M. Molecular recognition of fatty acids by peroxisome proliferator-activated receptors. Mol. Cell 1999, 3, 397–403. [Google Scholar]
- Mersmann, H.J. Mechanisms for conjugated linoleic acid-mediated reduction in fat deposition. J. Anim. Sci 2002, 80, E126–E134. [Google Scholar]
- Schoonjans, K.; Staels, B.; Auwerx, J. The peroxisome proliferator activated receptors (PPARs) and their effects on lipid metabolism and adipocyte differentiation. Biochim. Biophys. Acta 1996, 1302, 93–109. [Google Scholar]
- Dreyer, C.; Krey, G.; Keller, H.; Givel, F.; Helftenbein, G.; Wahli, W. Control of the peroxisomal beta-oxidation pathway by a novel family of nuclear hormone receptors. Cell 1992, 68, 879–887. [Google Scholar]
- Eubank, D.W.; Duplus, E.; Williams, S.C.; Forest, C.; Beale, E.G. Peroxisome proliferator-activated receptor γ and chicken ovalbumin upstream promoter transcription factor II negatively regulate the phosphoenolpyruvate carboxykinase promoter via a common element. J. Biol. Chem 2001, 276, 30561–30569. [Google Scholar]
- Tontonoz, P.; Kim, J.B.; Graves, R.A.; Spiegelman, B.M. ADD1: A novel helix-loop-helix transcription factor associated with adipocyte determination and differentiation. Mol. Cell. Biol 1993, 13, 4753–4759. [Google Scholar]
- Hua, X.; Wu, J.; Goldstein, J.L.; Brown, M.S.; Hobbs, H.H. Structure of the human gene encoding sterol regulatory element binding protein-1 (SREBF1) and localization of SREBF1 and SREBF2 to chromosomes 17p11.2 and 22q13. Genomics 1995, 25, 667–673. [Google Scholar]
- Bell, A.R.; Savory, R.; Horley, N.J.; Choudhury, A.I.; Dickins, T.J.; Gray, A.M.; Salter, D.R. Molecular basis of non-responsiveness to peroxisome proliferators: The guinea-pig PPARalpha is functional and mediates peroxisome proliferator-induced hypolipidaemia. Biochem. J 1998, 332, 689–693. [Google Scholar]
- Pineda Torra, I.; Gervois, P.; Staels, B. Peroxisome proliferator activated receptor alpha in metabolic disease, inflammation, atherosclerosis and aging. Curr. Opin. Lipidol 1999, 10, 151–159. [Google Scholar]
- Power, G.W.; Newsholme, E.A. Dietary fatty acids influence the activity and metabolic control of mitochondrial carnitine palmitoyltransferase I in rat heart and skeletal muscle. J. Nutr 1997, 127, 2142–2150. [Google Scholar]
- Sato, K.; Fukao, K.; Seki, Y.; Akiba, Y. Expression of the chicken peroxisome proliferator-activated receptor-γ gene is influenced by aging, nutrition, and agonist administration. Poult. Sci 2004, 83, 1342–1347. [Google Scholar]
- Sato, K.; Matsushita, K.; Matsubara, Y.; Kamada, T.; Akiba, Y. Adipose tissue fat accumulation is reduced by a single intraperitoneal injection of peroxisome proliferator-activated receptor gamma agonist when given to newly hatched chicks. Poult. Sci 2008, 87, 2281–2286. [Google Scholar]
- Carmona, M.C.; Louche, K.; Nibbelink, M.; Prunet, B.; Bross, A.; Desbazeille, M.; Dacquet, C.; Renard, P.; Casteilla, L.; Penicaud, L. Fenofibrate prevents rosiglitazone-induced body weight gain in ob/ob mice. Int. J. Obes. (Lond.) 2005, 29, 864–871. [Google Scholar]
- Lo Verme, J.; Gaetani, S.; Fu, J.; Oveisi, F.; Burton, K.; Piomelli, D. Regulation of food intake by oleoylethanolamide. Cell. Mol. Life Sci 2005, 62, 708–716. [Google Scholar]
- Meng, H.; Li, H.; Zhao, J.G.; Gu, Z.L. Differential expression of peroxisome proliferator-activated receptors alpha and gamma gene in various chicken tissues. Domest. Anim. Endocrinol 2005, 28, 105–110. [Google Scholar]
- Griffin, H.D.; Guo, K.; Windsor, D.; Butterwith, S.C. Adipose tissue lipogenesis and fat deposition in leaner broiler chickens. J. Nutr 1992, 122, 363–368. [Google Scholar]
- Griffin, H.D.; Hermier, D. Plasma Lipoprotein Metabolism and Fattening on Poultry. In Leanness in Domestic Birds; Leclercq, B., Whitehead, C.C., Eds.; Butterworths: London, UK, 1988; pp. 175–201. [Google Scholar]
- Sato, K.; Akiba, Y.; Chida, Y.; Takahashi, K. Lipoprotein hydrolysis and fat accumulation in chicken adipose tissues are reduced by chronic administration of lipoprotein lipase monoclonal antibodies. Poult. Sci 1999, 78, 1286–1291. [Google Scholar]
- Kliewer, S.A.; Sundseth, S.S.; Jones, S.A.; Brown, P.J.; Wisely, G.B.; Koble, C.S.; Devchand, P.; Wahli, W.; Wilson, T.M.; Lenhard, J.M.; et al. Fatty acids and eicosanoids regulate gene expression through direct interactions with peroxisome proliferator-activated receptors alpha and gamma. Proc. Natl. Acad. Sci. USA 1997, 94, 4318–4323. [Google Scholar]
- Hwang, D. Fatty acids and immune responses—a new perspective in searching for clues to mechanism. Annu. Rev. Nutr 2000, 20, 431–456. [Google Scholar]
- Kersten, S. Effects of fatty acids on gene expression: Role of peroxisome proliferator-activated receptor, liver X receptor and sterol regulatory element-binding protein-1c. Proc. Nutr. Soc 2002, 61, 371–374. [Google Scholar]
- Vidal-Puig, A.; Jimenez-Linan, M.; Lowell, B.B.; Hamann, A.; Hu, E.; Spiegelman, B.; Flier, J.S.; Moller, D.E. Regulation of PPAR gene expression by nutrition and obesity in rodents. J. Clin. Invest 1996, 97, 2553–2561. [Google Scholar]
- Nisoli, E.; Carruba, M.O.; Tonello, C.; Macor, C.; Federspil, G.; Vettor, R. Induction of fatty acid translocase/CD36, peroxisome proliferator-activated receptor-γ2, leptin, uncoupling proteins 2 and 3, and tumor necrosis factor-γ gene expression in human subcutaneous fat by lipid infusion. Diabetes 2000, 49, 319–324. [Google Scholar]
- Roche, H.M. Fatty acids and the metabolic syndrome. Proc. Nutr. Soc 2005, 64, 23–29. [Google Scholar]
- Zhao, A.; Yu, J.; Lew, J.L.; Huang, L.; Wright, S.D.; Cui, J. Polyunsaturated fatty acids are FXR ligands and differentially regulate expression of FXR targets. DNA Cell. Biol 2004, 23, 519–526. [Google Scholar]
- Tontonoz, P.; Hu, E.; Graves, R.A.; Budavari, A.I.; Spiegelman, B.M. mPPARgamma 2: Tissue-specific regulator of an adipocyte enhancer. Genes Dev 1994, 8, 1224–1234. [Google Scholar]
- Nakatani, T.; Katsumata, A.; Miura, S.; Kamei, Y.; Ezaki, O. Effects of fish oil feeding and fasting on LXRalpha/RXRalpha binding to LXRE in the SREBP-1c promoter in mouse liver. Biochim. Biophys. Acta 2004, 1736, 77–86. [Google Scholar]
- Ding, S.T.; Li, Y.C.; Nestor, K.E.; Velleman, S.G.; Mersmann, H.J. Expression of Turkey transcription factors and acyl-coenzyme oxidase in different tissues and genetic populations. Poult. Sci 2003, 82, 17–24. [Google Scholar]
- Issemann, I; Green, S. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature 1990, 347, 645–650. [Google Scholar]
- Schmidt, A.; Endo, N.; Rutledge, S.J.; Vogel, R.; Shinar, D.; Rodan, G.A. Identification of a new member of the steroid hormone receptor superfamily that is activated by a peroxisome proliferator and fatty acids. Mol. Endocrinol 1992, 6, 1634–1641. [Google Scholar]
- Frenkel, B.; Bisbara-Shieban, J.; Bar-Tana, J. The effect of beta, beta-tetramethylhexadecanedioic acid (MEDICA 16) on plasma very-low-density lipoprotein metabolism in rats: Role of apolipoprotein C-III. Biochem. J 1994, 298, 409–414. [Google Scholar]
- Frenkel, B.; Mayorek, N.; Hertz, R.; Bar-Tana, J. The hypochylomicronemic effect of beta, beta’-methyl-substituted hexadecanedioic acid (MEDICA 16) is mediated by a decrease in apolipoprotein C-III. J. Biol. Chem 1988, 263, 8491–8497. [Google Scholar]
- Shearer, B.G.; Hoekstra, W.J. Recent advances in peroxisome proliferator-activated receptor science. Curr. Med. Chem 2003, 10, 267–280. [Google Scholar]
- Sher, T.; Yi, H.F.; McBride, O.W.; Gonzalez, F.J. cDNA cloning, chromosomal mapping, and functional characterization of the human peroxisome proliferator activated receptor. Biochemistry 1993, 32, 5598–5604. [Google Scholar]
- Cherian, G.; Goeger, M.P. Hepatic lipid characteristics and histopathology of laying hens fed CLA or n-3 fatty acids. Lipids 2004, 39, 31–36. [Google Scholar]
- Folch, J.; Lees, M.; Sloan-Stanley, G.H. Asimple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem 1957, 226, 497–507. [Google Scholar]
- Konig, B.; Kluge, K.; Haase, K.; Brandsh, C.; Stangle, G.I.; Eder, K. Effects of clofibrate treatment in laing hens. Poult. Sci 2007, 86, 1187–1195. [Google Scholar]
- Zhang, H.J.; Guoab, Y.M.; Yangab, Y.; Yuanab, J.M. Dietary conjugated linoleic acid enhances spleen PPAR-γ mRNA expression in broiler chicks. Br. Poult. Sci 2006, 47, 726–733. [Google Scholar]
- Livak, K.J.; Schmittengen, T.D. Methods. Analysis of relative gene expression data using realtime quantitative PCR and the 2(-Delta Delta C(T)). Methods 2001, 25, 402–408. [Google Scholar]
- SAS Software Version 9.1.; SAS Institute Inc: Cary, NC, USA, 2002.



{kind=link}
{kind=link}
| Liver Fat content (% of dry matter) | Abdominal Fat Pad (% of live weight) | |
|---|---|---|
| PO 1 | 3.34 b | 2.35 a |
| SO | 2.85 c | 1.86 b |
| FO | 2.42 d | 2.03 b |
| CLA | 4.22 a | 2.14 b |
| CLA + SO | 4.13 a | 2.13 b |
| CLA + FO | 3.86 a,b | 2.14 b |
| FO + SO | 3.16 b,c | 2.12 b |
| Pooled SEM | 0.16 | 0.08 |
| Ingredients (as-fed basis) | Starter (1-10 d) | Grower (11–28 days of age) | Finisher (29–42 days of age) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PO2 | SO | FO | CLA | FO + SO | CLA + SO | CLA + FO | PO | SO | FO | CLA | FO + SO | CLA + SO | CLA + FO | ||
| Corn (%) | 60.23 | 48.46 | 53.99 | 53.99 | 55.8 | 53.99 | 54.13 | 54.00 | 52.62 | 57.98 | 57.98 | 59.50 | 57.98 | 59.06 | 58.92 |
| Soy meal (%) | 30.81 | 30.56 | 32.27 | 32.27 | 28.6 | 32.27 | 31.96 | 32.26 | 27.76 | 30.27 | 30.27 | 26.38 | 30.27 | 28.30 | 28.60 |
| Fish Meal (%) | 5.37 | 5.00 | 3.00 | 3.00 | 5.00 | 3.00 | 3.20 | 3.01 | 3.00 | 1.00 | 1.00 | 2.99 | 1.00 | 1.70 | 1.51 |
| Soybean oil (%) | - | - | 7.00 | - | - | 3.50 | 3.50 | - | - | 7.00 | - | - | 3.50 | 3.50 | - |
| Fish oil | - | - | - | 7.00 | - | 3.50 | - | 3.50 | - | - | 7.00 | - | 3.50 | - | 3.50 |
| Palm oil (%) | - | 12.60 | - | - | - | - | - | - | 12.92 | - | - | - | - | - | - |
| CLA (%) 1 | - | - | - | - | 7.00 | - | 3.50 | 3.50 | - | - | - | 7.40 | - | 3.50 | 3.50 |
| Oyster shell (%) | 1.41 | 1.34 | 1.42 | 1.42 | 1.33 | 1.42 | 1.41 | 1.42 | 1.30 | 1.39 | 1.39 | 1.30 | 1.39 | 1.35 | 1.36 |
| DCP (%) | 0.51 | 0.48 | 0.66 | 0.66 | 0.52 | 0.66 | 0.64 | 0.66 | 0.69 | 0.84 | 0.84 | 0.71 | 0.84 | 0.81 | 0.82 |
| Salt (%) | 0.25 | 0.29 | 0.32 | 0.32 | 0.28 | 0.32 | 0.31 | 0.32 | 0.32 | 0.35 | 0.35 | 0.32 | 0.35 | 0.34 | 0.34 |
| Vit-Min P (%) 3 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| DL-Met (%) | 0.26 | 0.23 | 0.25 | 0.25 | 0.23 | 0.25 | 0.25 | 0.25 | 0.17 | 0.18 | 0.18 | 0.16 | 0.18 | 0.18 | 0.18 |
| L-Lys (%) | 0.15 | 0.04 | 0.09 | 0.09 | 0.24 | 0.09 | 0.09 | 0.09 | 0.22 | - | - | 0.25 | - | 0.27 | 0.27 |
| Analysis | |||||||||||||||
| ME (Kcal/Kg) | 2860 | 3175 | 3205 | 3211 | 3175 | 3208 | 3175 | 3175 | 3225 | 3235 | 3241 | 3225 | 3238 | 3225 | 3225 |
| CP (%) | 22.5 | 21.00 | 21.00 | 21.00 | 21.00 | 21.00 | 21.00 | 21.00 | 19.00 | 19.00 | 19.00 | 19.00 | 19.00 | 19.00 | 19.00 |
| Crude Fat (%) | 2.86 | 15.00 | 9.52 | 9.52 | 9.65 | 9.52 | 9.54 | 9.52 | 15.34 | 9.55 | 9.55 | 10.07 | 9.50 | 9.61 | 9.60 |
| Linoleic a (%) | 1.46 | 2.63 | 5.37 | 1.39 | 5.27 | 3.38 | 5.30 | 3.31 | 2.74 | 5.45 | 1.46 | 5.56 | 3.46 | 5.40 | 3.40 |
| Ca (%) | 0.95 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 | 0.85 | 0.85 | 0.85 | 0.85 | 0.85 | 0.85 | 0.85 |
| Ava P (%) | 0.48 | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 | 0.425 | 0.425 | 0.425 | 0.43 | 0.43 | 0.43 | 0.43 |
| Na (%) | 0.15 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 |
| Lys (%) | 1.37 | 1.23 | 1.23 | 1.23 | 1.34 | 1.23 | 1.23 | 1.23 | 1.20 | 1.02 | 1.02 | 1.20 | 1.01 | 1.20 | 1.20 |
| Met (%) | 0.66 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 | 0.48 | 0.48 | 0.48 | 0.48 | 0.48 | 0.49 | 0.49 |
| Met + Cys (%) | 1.04 | 0.95 | 0.95 | 0.95 | 0.95 | 0.95 | 0.95 | 0.95 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 |
| Grower | Finisher | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PO 1 | SO | FO | CLA 2 | FO + SO | CLA + SO | CLA + FO | PO | SO | FO | CLA | FO + SO | CLA + SO | CLA + FO | |
| C14:0 | 1.52 | 0.24 | 3.211 | 0.33 | 1.65 | 0.23 | 1.72 | 1.52 | 0.12 | 3.13 | 0.22 | 1.62 | 0.11 | 1.62 |
| C16:0 | 58.63 | 10.71 | 20.4 | 8.43 | 15.52 | 9.32 | 14.10 | 59.51 | 10.16 | 19.91 | 8.12 | 15.06 | 8.92 | 13.81 |
| C16:1 | 0.86 | 0.66 | 4.50 | 0.38 | 2.51 | 0.37 | 2.42 | 0.84 | 0.32 | 4.31 | 0.22 | 2.36 | 0.18 | 3.00 |
| C17:0 | 0.13 | 0.13 | 1.03 | 0.08 | 0.46 | 0.11 | 0.45 | 0.12 | 0.02 | 0.91 | 0.13 | 0.47 | 0.03 | 0.52 |
| C17:1 | 0.12 | 0.04 | 0.53 | 0.02 | 0.28 | 0.04 | 0.31 | 0.04 | 0.01 | 0.52 | 0.01 | 0.27 | 0.02 | 0.21 |
| C18:0 | 6.12 | 3.50 | 4.27 | 3.71 | 3.89 | 3.53 | 3.92 | 6.12 | 3.40 | 4.17 | 3.74 | 3.82 | 3.42 | 3.84 |
| C18:1c | 54.33 | 23.72 | 31.42 | 24.24 | 27.59 | 23.61 | 27.31 | 55.00 | 23.22 | 30.86 | 24.47 | 27.12 | 23.31 | 27.12 |
| C18:2c | 23.91 | 49.41 | 15.84 | 15.43 | 32.62 | 32.32 | 15.48 | 24.88 | 50.0-6 | 16.50 | 16.08 | 33.26 | 33.14 | 16.3 |
| C18:3 alpha | 0.02 | 0.02 | 0.12 | 0.03 | 0.07 | 0.02 | 0.07 | 0.02 | 0.02 | 0.11 | 0.03 | 0.07 | 0.02 | 0.07 |
| C18:3 gamma | 0.31 | 0.34 | 1.11 | 0.33 | 0.73 | 0.27 | 0.71 | 0.21 | 0.20 | 1.07 | 0.32 | 0.61 | 0.34 | 0.74 |
| C20:0 | 0.02 | 0.03 | 0.12 | 0.04 | 0.11 | 0.03 | 0.12 | 0.02 | 0.04 | 0.14 | 0.02 | 0.09 | 0.01 | 0.11 |
| CLA-9c, 11t | 0.05 | 0.00 | 0.00 | 21.00 | 0.00 | 10.49 | 10.51 | 0.00 | 0.00 | 0.00 | 22.20 | 0.00 | 10.50 | 10.50 |
| CLA-10c,12t | 0.04 | 0.00 | 0.00 | 21.00 | 0.00 | 10.52 | 10.46 | 0.00 | 0.00 | 0.00 | 22.20 | 0.00 | 10.50 | 10.50 |
| C20:5 n-3 | 0.44 | 0.18 | 4.11 | 0.44 | 2.23 | 0.20 | 2.22 | 0.23 | 0.13 | 4.03 | 0.21 | 2.00 | 0.08 | 2.06 |
| C22:6 n-3 | 0.92 | 0.51 | 9.76 | 0.92 | 5.07 | 0.61 | 5.20 | 0.46 | 0.16 | 9.50 | 0.54 | 4.82 | 0.32 | 4.92 |
| n6/n3 | 18.31 | 69.00 | 1.23 | 11.82 | 4.54 | 39.84 | 2.18 | 34.92 | 157.22 | 1.33 | 22.54 | 4.91 | 79.48 | 2.40 |
| Gene | Primer | Base pairs | Annealing temperature(Ċ) | References |
|---|---|---|---|---|
| B-actin | F-5′ATGAAGCCCAGAGCAAAAGA3′ | 223 | 62 | [64] |
| R-5′GGGGTGTTGAAGGTCTCAAA3′ | ||||
| PPARα | F-5′AGGCCAAGTTGAAAGCAGAA3′ | 217 | 60 | [64] |
| R-5′GTCTTCTCTGCCATGCACAA3′ | ||||
| PPARγ | F-5′GACCTTAATTGTCGCATCCAT3′ | 237 | 61 | [65] |
| R-5′CGGGAAGGACTTTATGTATGA3′ |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Royan, M.; Meng, G.Y.; Othman, F.; Sazili, A.Q.; Navidshad, B. Effects of Conjugated Linoleic Acid, Fish Oil and Soybean Oil on PPARs (α & γ) mRNA Expression in Broiler Chickens and Their Relation to Body Fat Deposits. Int. J. Mol. Sci. 2011, 12, 8581-8595. https://doi.org/10.3390/ijms12128581
Royan M, Meng GY, Othman F, Sazili AQ, Navidshad B. Effects of Conjugated Linoleic Acid, Fish Oil and Soybean Oil on PPARs (α & γ) mRNA Expression in Broiler Chickens and Their Relation to Body Fat Deposits. International Journal of Molecular Sciences. 2011; 12(12):8581-8595. https://doi.org/10.3390/ijms12128581
Chicago/Turabian StyleRoyan, Maryam, Goh Yong Meng, Fauziah Othman, Awis Qurni Sazili, and Bahman Navidshad. 2011. "Effects of Conjugated Linoleic Acid, Fish Oil and Soybean Oil on PPARs (α & γ) mRNA Expression in Broiler Chickens and Their Relation to Body Fat Deposits" International Journal of Molecular Sciences 12, no. 12: 8581-8595. https://doi.org/10.3390/ijms12128581