Integrated Lipidomics in the Secreted Phospholipase A2 Biology

Abstract
:1. Introduction
2. Biological Functions of sPLA2s in Vivo
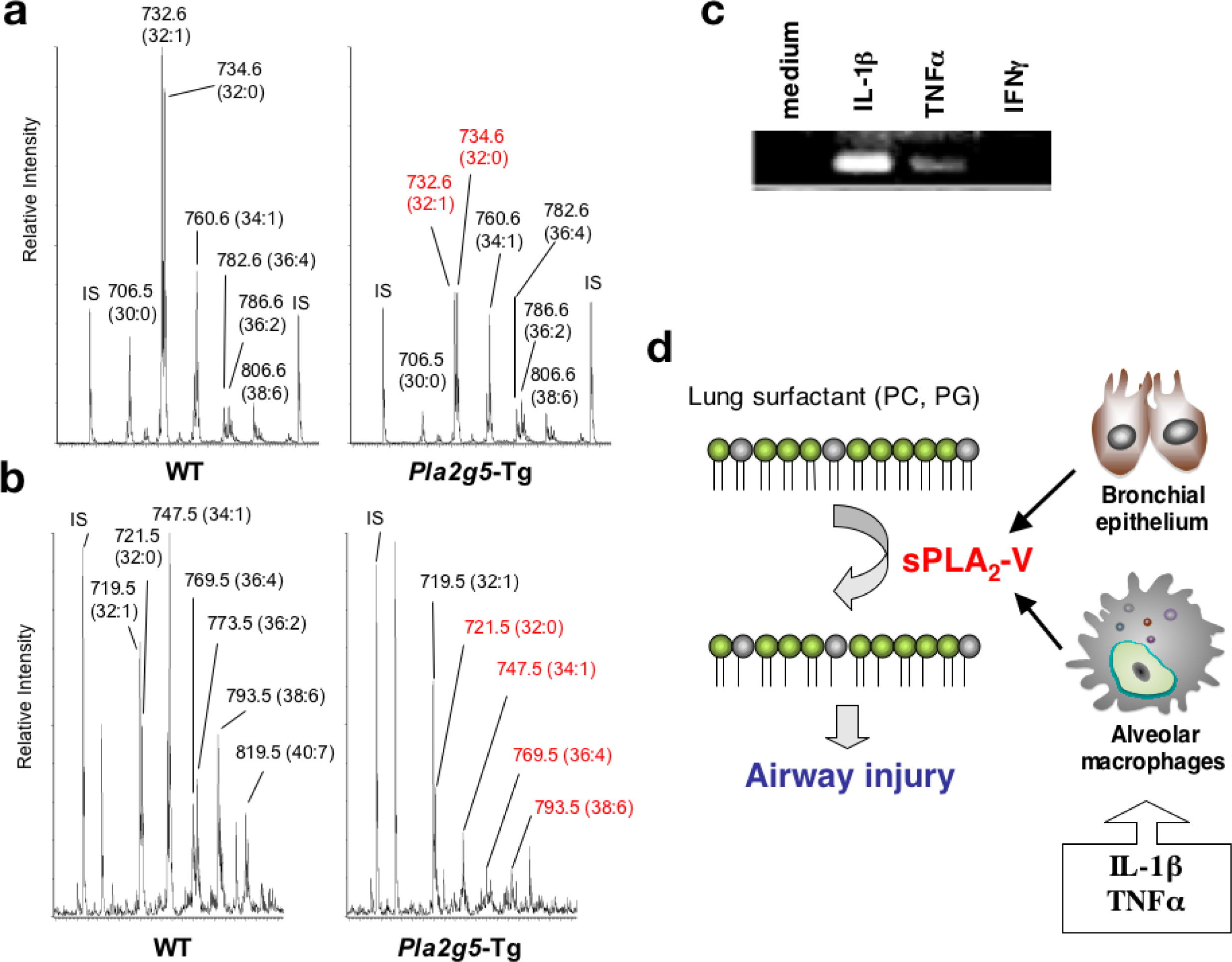
2.1. Group V sPLA2 (sPLA2-V)
2.2. Group X sPLA2 (sPLA2-X)
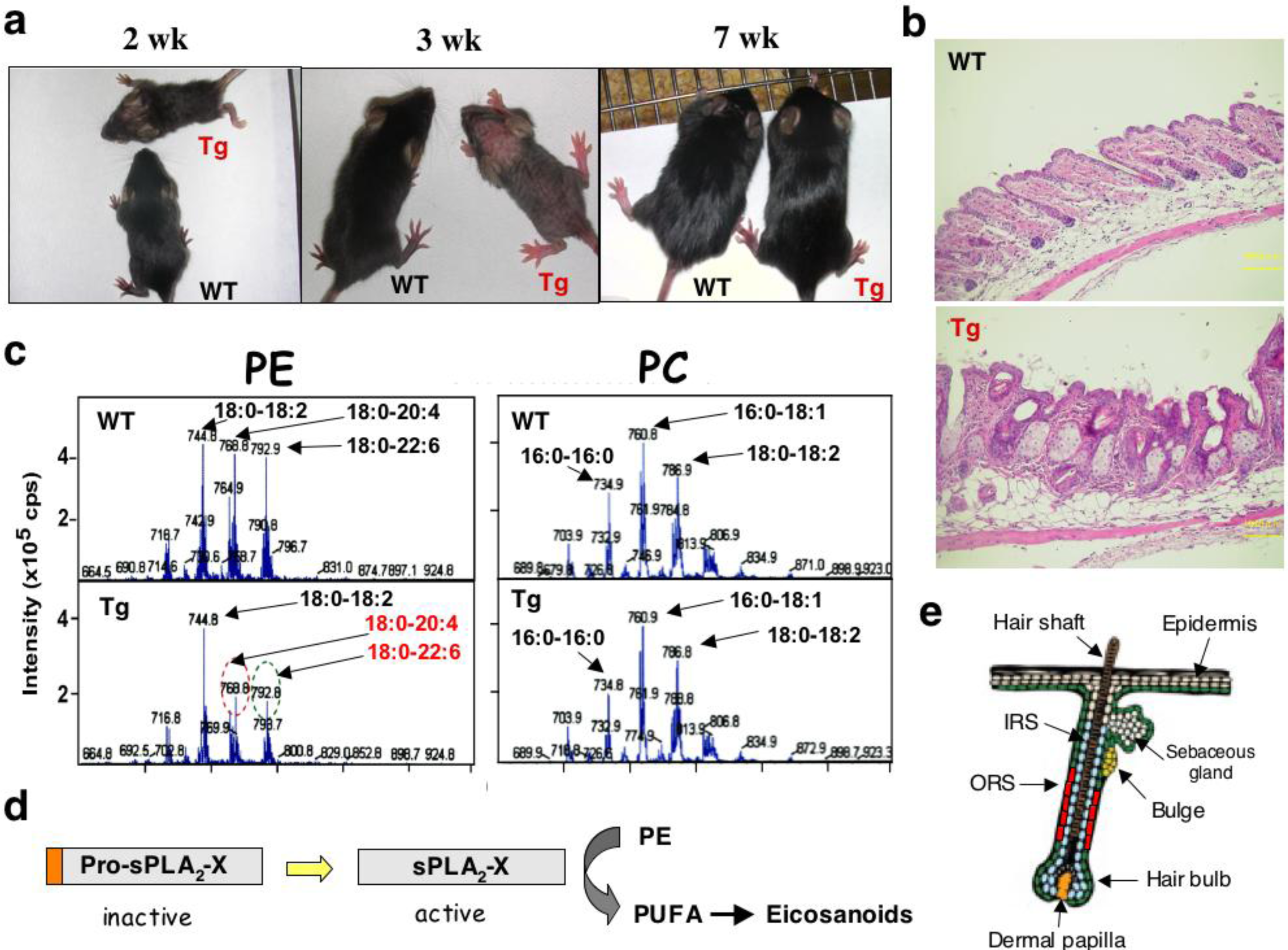
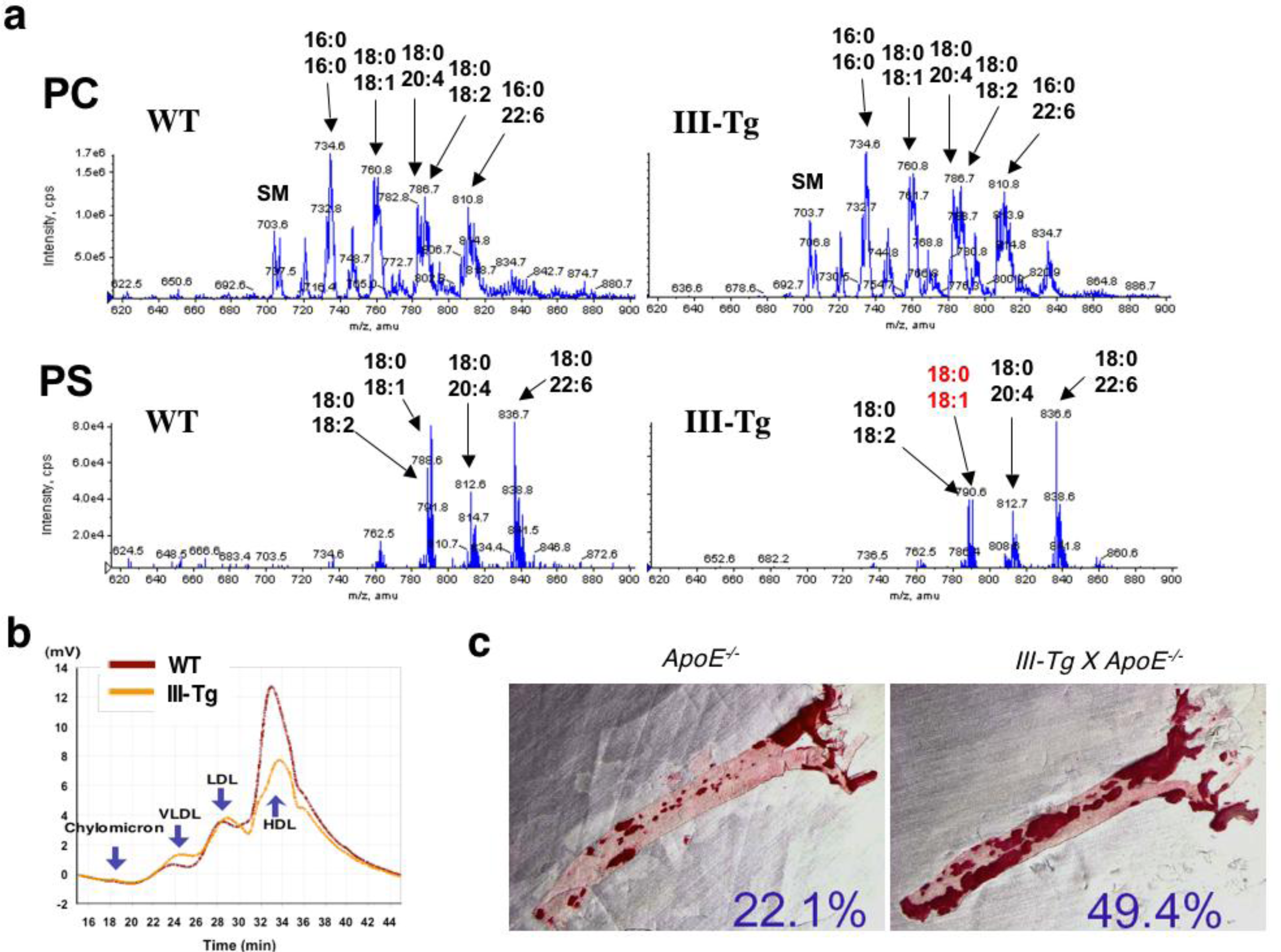
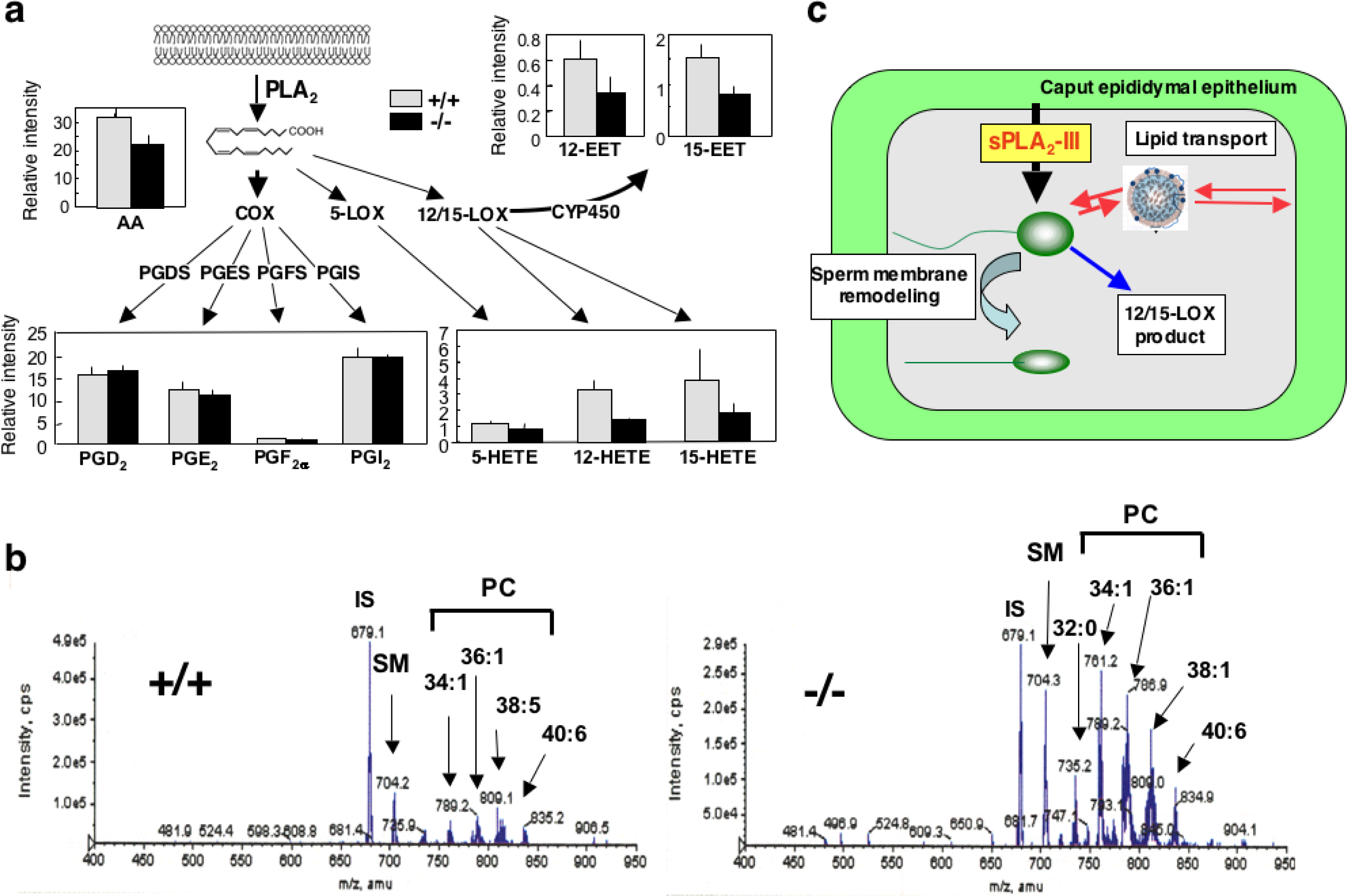
2.3. Group III sPLA2 (sPLA2-III)
2.4. Other sPLA2s: Classical and Novel Enzymes
3. Conclusions
Abbreviations:
| PLA2 | phospholipase A2; |
| sPLA2 | secreted PLA2; |
| cPLA2 | cytosolic PLA2; |
| iPLA2 | Ca2+-independent PLA2; |
| Tg | transgenic; |
| ARDS | acute respiratory distress syndrome; |
| BALF | bronchoalveolar fluid; |
| LPS | lipopolysaccharide; |
| ESI-MS | electrospray ionization mass spectrometry; |
| PC | phosphatidylcholine; |
| LPC | lysophosphatidylcholine; |
| PG | phosphatidylglycerol; |
| PE | phosphatidylethanolamine; |
| PS | phosphatidylserine; |
| LDL | low-density lipoprotein; |
| HDL | high-density lipoprotein; |
| VLDL | very low-density lipoprotein; |
| ABC | ATP-binding cassette; |
| PGD2 | prostaglandin D2; |
| PGE2 | prostaglandin E2; |
| LTB4 | leukotriene B4; |
| PUFA | polyunsaturated fatty acid; |
| DHA | docosahexaenoic acid; |
| DPA | docosapentaenoic acid; |
| LXR | liver X receptor; |
| PPAR | peroxisome proliferator-activated receptor; |
| StAR | steroidogenic acute regulatory protein; |
| DRG | dorsal root ganglion; |
| COX | cyclooxygenase; |
| LOX | lipoxygenase; |
| CYP450 | cytochrome P450; |
| HPLC | high performance liquid chromatography; |
| WT | wild-type. |
Acknowledgments
References
- Uozumi, N; Kume, K; Nagase, T; Nakatani, N; Ishii, S; Tashiro, F; Komagata, Y; Maki, K; Ikuta, K; Ouchi, Y; Miyazaki, J; Shimizu, T. Role of cytosolic phospholipase A2 in allergic response and parturition. Nature 1997, 390, 618–622. [Google Scholar]
- Kudo, I; Murakami, M. Phospholipase A2 enzymes. Prostag Other Lipid Mediat 2002, 68–69, 3–58. [Google Scholar]
- Lambeau, G; Gelb, MH. Biochemistry and physiology of mammalian secreted phospholipases A2. Annu. Rev. Biochem 2008, 77, 495–520. [Google Scholar]
- Murakami, M; Taketomi, Y; Girard, C; Yamamoto, K; Lambeau, G. Emerging roles of secreted phospholipase A2 enzymes: Lessons from transgenic and knockout mice. Biochimie 2010, 92, 561–582. [Google Scholar]
- Chen, J; Engle, SJ; Seilhamer, JJ; Tischfield, JA. Cloning and recombinant expression of a novel human low molecular weight Ca2+-dependent phospholipase A2. J. Biol. Chem 1994, 269, 2365–2368. [Google Scholar]
- Munoz, NM; Meliton, AY; Arm, JP; Bonventre, JV; Cho, W; Leff, AR. Deletion of secretory group V phospholipase A2 attenuates cell migration and airway hyperresponsiveness in immunosensitized mice. J. Immunol 2007, 179, 4800–4807. [Google Scholar]
- Masuda, S; Murakami, M; Mitsuishi, M; Komiyama, K; Ishikawa, Y; Ishii, T; Kudo, I. Expression of secretory phospholipase A2 enzymes in lungs of humans with pneumonia and their potential prostaglandin-synthetic function in human lung-derived cells. Biochem. J 2005, 387, 27–38. [Google Scholar]
- Ohtsuki, M; Taketomi, Y; Arata, S; Masuda, S; Ishikawa, Y; Ishii, T; Takanezawa, Y; Aoki, J; Arai, H; Yamamoto, K; Kudo, I; Murakami, M. Transgenic expression of group V, but not group X, secreted phospholipase A2 in mice leads to neonatal lethality because of lung dysfunction. J. Biol. Chem 2006, 281, 36420–36433. [Google Scholar]
- Touqi, L; Arbibe, L. A role for phospholipase A2 in ARDS pathogenesis. Mol. Med. Today 1999, 5, 244–249. [Google Scholar]
- Nag, K; Munro, JG; Inchley, K; Schürch, S; Petersen, NO; Possmayer, F. SP-B refining of pulmonary surfactant phospholipid films. Am. J. Physiol 1999, 277, L1179–L1189. [Google Scholar]
- Arbibe, L; Koumanov, K; Vial, D; Rougeot, C; Faure, G; Havet, N; Longacre, S; Vargaftig, BB; Béréziat, G; Voelker, DR; Wolf, C; Touqui, L. Generation of lyso-phospholipids from surfactant in acute lung injury is mediated by type-II phospholipase A2 and inhibited by a direct surfactant protein A-phospholipase A2 protein interaction. J. Clin. Invest 1998, 102, 1152–1160. [Google Scholar]
- Wu, Y; Singer, M; Thouron, F; Alaoui-El-Azher, M; Touqui, L. Effect of surfactant on pulmonary expression of type IIA PLA2 in an animal model of acute lung injury. Am. J. Physiol. Lung. Cell Mol. Physiol 2002, 282, L743–L750. [Google Scholar]
- Wu, YZ; Medjane, S; Chabot, S; Kubrusly, FS; Raw, I; Chignard, M; Touqui, L. Surfactant protein-A and phosphatidylglycerol suppress type IIA phospholipase A2 synthesis via nuclear factor-κB. Am. J. Respir. Crit. Care Med 2003, 168, 692–699. [Google Scholar]
- Wang, Z; Schwan, AL; Lairson, LL; O’Donnell, JS; Byrne, GF; Foye, A; Holm, BA; Notter, RH. Surface activity of a synthetic lung surfactant containing a phospholipase-resistant phosphonolipid analog of dipalmitoyl phosphatidylcholine. Am. J. Physiol. Lung Cell Mol. Physiol 2003, 285, L550–L559. [Google Scholar]
- Furue, S; Kuwabara, K; Mikawa, K; Nishina, K; Shiga, M; Maekawa, N; Ueno, M; Chikazawa, Y; Ono, T; Hori, Y; Matsukawa, A; Yoshinaga, M; Obara, H. Crucial role of group IIA phospholipase A2 in oleic acid-induced acute lung injury in rabbits. Crit. Care Med 2001, 29, 719–727. [Google Scholar]
- Chabot, S; Koumanov, K; Lambeau, G; Gelb, MH; Balloy, V; Chignard, M; Whitsett, JA; Touqui, L. Inhibitory effects of surfactant protein A on surfactant phospholipid hydrolysis by secreted phospholipases A2. J. Immunol 2003, 171, 995–1000. [Google Scholar]
- Seeds, MC; Jones, KA; Duncan, HR; Willingham, MC; Borgerink, HM; Woodruff, RD; Bowton, DL; Bass, DA. Cell-specific expression of group X and group V secretory phospholipases A2 in human lung airway epithelial cells. Am. J. Respir. Cell Mol. Biol 2000, 23, 37–44. [Google Scholar]
- Hite, RD; Seeds, MC; Safta, AM; Jacinto, RB; Gyves, JI; Bass, DA; Waite, BM. Lysophospholipid generation and phosphatidylglycerol depletion in phospholipase A2-mediated surfactant dysfunction. Am. J. Physiol. Lung Cell Mol. Physiol 2005, 288, L618–L624. [Google Scholar]
- Shulenin, S; Nogee, LM; Annilo, T; Wert, SE; Whitsett, JA; Dean, M. ABCA3 gene mutations in newborns with fatal surfactant deficiency. N. Engl. J. Med 2004, 350, 1296–1303. [Google Scholar]
- Bridges, JP; Ikegami, M; Brilli, LL; Chen, X; Mason, RJ; Shannon, JM. LPCAT1 regulates surfactant phospholipid synthesis and is required for transitioning to air breathing in mice. J. Clin. Invest 2010, 120, 1736–1748. [Google Scholar]
- Munoz, NM; Meliton, AY; Meliton, LN; Dudek, SM; Leff, AR. Secretory group V phospholipase A2 regulates acute lung injury and neutrophilic inflammation caused by LPS in mice. Am. J. Physiol. Lung Cell Mol. Physiol 2009, 296, L879–L887. [Google Scholar]
- Giannattasio, G; Fujioka, D; Xing, W; Katz, HR; Boyce, JA; Balestrieri, B. Group V secretory phospholipase A2 reveals its role in house dust mite-induced allergic pulmonary inflammation by regulation of dendritic cell function. J. Immunol 2010, 185, 4430–4438. [Google Scholar]
- Satake, Y; Diaz, BL; Balestrieri, B; Lam, BK; Kanaoka, Y; Grusby, MJ; Arm, JP. Role of group V phospholipase A2 in zymosan-induced eicosanoid generation and vascular permeability revealed by targeted gene disruption. J. Biol. Chem 2004, 279, 16488–16494. [Google Scholar]
- Balestrieri, B; Hsu, VW; Gilbert, H; Leslie, CC; Han, WK; Bonventre, JV; Arm, JP. Group V secretory phospholipase A2 translocates to the phagosome after zymosan stimulation of mouse peritoneal macrophages and regulates phagocytosis. J. Biol. Chem 2006, 281, 6691–6698. [Google Scholar]
- Balestrieri, B; Maekawa, A; Xing, W; Gelb, MH; Katz, HR; Arm, JP. Group V secretory phospholipase A2 modulates phagosome maturation and regulates the innate immune response against Candida albicans. J. Immunol 2009, 182, 4891–4898. [Google Scholar]
- Boilard, E; Lai, Y; Larabee, K; Balestrieri, B; Ghomashchi, F; Fujioka, D; Gobezie, R; Coblyn, JS; Weinblatt, ME; Massarotti, EM; Thornhill, TS; Divangahi, M; Remold, H; Lambeau, G; Gelb, MH; Arm, JP; Lee, DM. A novel anti-inflammatory role for secretory phospholipase A2 in immune complex-mediated arthritis. EMBO Mol. Med 2010, 2, 172–187. [Google Scholar]
- Bostrom, MA; Boyanovsky, BB; Jordan, CT; Wadsworth, MP; Taatjes, DJ; De Beer, FC; Webb, NR. Group V secretory phospholipase A2 promotes atherosclerosis: Evidence from genetically altered mice. Arterioscler. Thromb. Vasc. Biol 2007, 27, 600–606. [Google Scholar]
- Wootton, PT; Arora, NL; Drenos, F; Thompson, SR; Cooper, JA; Stephens, JW; Hurel, SJ; Hurt-Camejo, E; Wiklund, O; Humphries, SE; Talmud, PJ. Tagging SNP haplotype analysis of the secretory PLA2-V gene, PLA2G5, shows strong association with LDL and oxLDL levels, suggesting functional distinction from sPLA2-IIA: Results from the UDACS study. Hum. Mol. Genet 2007, 16, 1437–1444. [Google Scholar]
- Cupillard, L; Koumanov, K; Mattéi, MG; Lazdunski, M; Lambeau, G. Cloning, chromosomal mapping, and expression of a novel human secretory phospholipase A2. J. Biol. Chem 1997, 272, 15745–15752. [Google Scholar]
- Murakami, M; Koduri, RS; Enomoto, A; Shimbara, S; Seki, M; Yoshihara, K; Singer, A; Valentin, E; Ghomashchi, F; Lambeau, G; Gelb, MH; Kudo, I. Distinct arachidonate-releasing functions of mammalian secreted phospholipases A2 in fibroblastic and mastocytoma cells through heparan sulfate shuttling and external plasma membrane mechanisms. J. Biol. Chem 2001, 276, 10083–10096. [Google Scholar]
- Bezzine, S; Koduri, RS; Valentin, E; Murakami, M; Kudo, I; Ghomashchi, F; Sadilek, M; Lambeau, G; Gelb, MH. Exogenously added human group X secreted phospholipase A2 but not the group IB, IIA, and V enzymes efficiently release arachidonic acid from adherent mammalian cells. J. Biol. Chem 2000, 275, 3179–3191. [Google Scholar]
- Escoffier, J; Jemel, I; Tanemoto, A; Taketomi, Y; Payre, C; Coatrieux, C; Sato, H; Yamamoto, K; Masuda, S; Pernet-Gallay, K; Pierre, V; Hara, S; Murakami, M; De Waard, M; Lambeau, G; Arnoult, C. Group X phospholipase A2 is released during sperm acrosome reaction and controls fertility outcome in mice. J. Clin. Invest 2010, 120, 1415–1428. [Google Scholar]
- Sato, H; Isogai, Y; Masuda, S; Taketomi, Y; Miki, Y; Kamei, D; Hara, S; Kobayashi, T; Ishikawa, Y; Ishii, T; Ikeda, K; Taguchi, R; Ishimoto, Y; Suzuki, N; Yokota, Y; Hanasaki, K; Suzuki-Yamamoto, T; Yamamoto, K; Murakami, M. Physiological roles of group X secreted phospholipase A2 in reproduction, gastrointestinal phospholipid digestion, and neuronal function. J Biol Chem 2011, in press. [Google Scholar]
- Henderson, WR, Jr; Chi, EY; Bollinger, JG; Tien, YT; Ye, X; Castelli, L; Rubtsov, YP; Singer, AG; Chiang, GK; Nevalainen, T; Rudensky, AY; Gelb, MH. Importance of group X-secreted phospholipase A2 in allergen-induced airway inflammation and remodeling in a mouse asthma model. J. Exp. Med 2007, 204, 865–877. [Google Scholar]
- Hallstrand, TS; Chi, EY; Singer, AG; Gelb, MH; Henderson, WR, Jr. Secreted phospholipase A2 group X overexpression in asthma and bronchial hyperresponsiveness. Am. J. Respir. Crit. Care Med 2007, 176, 1072–1078. [Google Scholar]
- Fujioka, D; Saito, Y; Kobayashi, T; Yano, T; Tezuka, H; Ishimoto, Y; Suzuki, N; Yokota, Y; Nakamura, T; Obata, JE; Kanazawa, M; Kawabata, K; Hanasaki, K; Kugiyama, K. Reduction in myocardial ischemia/reperfusion injury in group X secretory phospholipase A2-deficient mice. Circulation 2008, 117, 2977–2985. [Google Scholar]
- Curfs, DM; Ghesquiere, SA; Vergouwe, MN; Van der Made, I; Gijbels, MJ; Greaves, DR; Verbeek, JS; Hofker, MH; De Winther, MP. Macrophage secretory phospholipase A2 group X enhances anti-inflammatory responses, promotes lipid accumulation, and contributes to aberrant lung pathology. J. Biol. Chem 2008, 283, 21640–21648. [Google Scholar]
- Yamamoto, K; Taketomi, T; Isogai, Y; Miki, Y; Sato, H; Masuda, S; Nishito, Y; Morioka, K; Ishimoto, Y; Suzuki, N; Yokoya, Y; Hanasaki, K; Ishikawa, Y; Ishii, T; Kobayashi, T; Fukami, K; Ikeda, K; Nakanishi, H; Taguchi, R; Murakami, M. Hair follicular expression and function of group X secreted phospholipase A2 in mouse skin. J Biol Chem 2011, in press. [Google Scholar]
- Grass, DS; Felkner, RH; Chiang, MY; Wallace, RE; Nevalainen, TJ; Bennett, CF; Swanson, ME. Expression of human group II PLA2 in transgenic mice results in epidermal hyperplasia in the absence of inflammatory infiltrate. J. Clin. Invest 1996, 97, 2233–2241. [Google Scholar]
- Mulherkar, R; Kirtane, BM; Ramchandani, A; Mansukhani, NP; Kannan, S; Naresh, KN. Expression of enhancing factor/phospholipase A2 in skin results in abnormal epidermis and increased sensitivity to chemical carcinogenesis. Oncogene 2003, 22, 1936–1944. [Google Scholar]
- Hanasaki, K; Yamada, K; Yamamoto, S; Ishimoto, Y; Saiga, A; Ono, T; Ikeda, M; Notoya, M; Kamitani, S; Arita, H. Potent modification of low density lipoprotein by group X secretory phospholipase A2 is linked to macrophage foam cell formation. J. Biol. Chem 2002, 277, 29116–29124. [Google Scholar]
- Zack, M; Boyanovsky, BB; Shridas, P; Bailey, W; Forrest, K; Howatt, DA; Gelb, MH; De Beer, FC; Daugherty, A; Webb, NR. Group X secretory phospholipase A2 augments angiotensin II-induced inflammatory responses and abdominal aortic aneurysm formation in apoE-deficient mice. Atherosclerosis 2010, in press. [Google Scholar]
- Shridas, P; Bailey, WM; Gizard, F; Oslund, RC; Gelb, MH; Bruemmer, D; Webb, NR. Group X secretory phospholipase A2 negatively regulates ABCA1 and ABCG1 expression and cholesterol efflux in macrophages. Arterioscler Thromb. Vasc. Biol 2010, 30, 2014–2021. [Google Scholar]
- Shridas, P; Bailey, WM; Boyanovsky, BB; Oslund, RC; Gelb, MH; Webb, NR. Group X secretory phospholipase A2 regulates the expression of steroidogenic acute regulatory protein (StAR) in mouse adrenal glands. J. Biol. Chem 2010, 285, 20031–20039. [Google Scholar]
- Li, X; Shridas, P; Forrest, K; Bailey, W; Webb, NR. Group X secretory phospholipase A2 negatively regulates adipogenesis in murine models. FASEB J 2010, 24, 4313–4324. [Google Scholar]
- Valentin, E; Ghomashchi, F; Gelb, MH; Lazdunski, M; Lambeau, G. Novel human secreted phospholipase A2 with homology to the group III bee venom enzyme. J. Biol. Chem 2000, 275, 7492–7496. [Google Scholar]
- Murakami, M; Masuda, S; Shimbara, S; Bezzine, S; Lazdunski, M; Lambeau, G; Gelb, MH; Matsukura, S; Kokubu, F; Adachi, M; Kudo, I. Cellular arachidonate-releasing function of novel classes of secretory phospholipase A2s (groups III and XII). J. Biol. Chem 2003, 278, 10657–10667. [Google Scholar]
- Murakami, M; Masuda, S; Shimbara, S; Ishikawa, Y; Ishii, T; Kudo, I. Cellular distribution, post-translational modification, and tumorigenic potential of human group III secreted phospholipase A2. J. Biol. Chem 2005, 280, 24987–24998. [Google Scholar]
- Masuda, S; Yamamoto, K; Hirabayashi, T; Ishikawa, Y; Ishii, T; Kudo, I; Murakami, M. Human group III secreted phospholipase A2 promotes neuronal outgrowth and survival. Biochem. J 2008, 409, 429–438. [Google Scholar]
- Mounier, CM; Wendum, D; Greenspan, E; Flejou, JF; Rosenberg, DW; Lambeau, G. Distinct expression pattern of the full set of secreted phospholipases A2 in human colorectal adenocarcinomas: sPLA2-III as a biomarker candidate. Br. J. Cancer 2008, 98, 587–595. [Google Scholar]
- Sato, H; Taketomi, Y; Isogai, Y; Masuda, S; Kobayashi, T; Yamamoto, K; Murakami, M. Group III secreted phospholipase A2 transgenic mice spontaneously develop inflammation. Biochem. J 2009, 421, 17–27. [Google Scholar]
- Emoto, K; Inadome, H; Kanaho, Y; Narumiya, S; Umeda, M. Local change in phospholipid composition at the cleavage furrow is essential for completion of cytokinesis. J. Biol. Chem 2005, 280, 37901–37907. [Google Scholar]
- Miyanishi, M; Tada, K; Koike, M; Uchiyama, Y; Kitamura, T; Nagata, S. Identification of Tim4 as a phosphatidylserine receptor. Nature 2007, 450, 435–439. [Google Scholar]
- Olson, ED; Nelson, J; Griffith, K; Nguyen, T; Streeter, M; Wilson-Ashworth, HA; Gelb, MH; Judd, AM; Bell, JD. Kinetic evaluation of cell membrane hydrolysis during apoptosis by human isoforms of secretory phospholipase A2. J. Biol. Chem 2010, 285, 10993–11002. [Google Scholar]
- Sato, H; Kato, R; Isogai, Y; Saka, G; Ohtsuki, M; Taketomi, Y; Yamamoto, K; Tsutsumi, K; Yamada, J; Masuda, S; Ishikawa, Y; Ishii, T; Kobayashi, T; Ikeda, K; Taguchi, R; Hatakeyama, S; Hara, S; Kudo, I; Itabe, H; Murakami, M. Analyses of group III secreted phospholipase A2 transgenic mice reveal potential participation of this enzyme in plasma lipoprotein modification, macrophage foam cell formation, and atherosclerosis. J. Biol. Chem 2008, 283, 33483–33497. [Google Scholar]
- Kimura-Matsumoto, M; Ishikawa, Y; Komiyama, K; Tsuruta, T; Murakami, M; Masuda, S; Akasaka, Y; Ito, K; Ishiguro, S; Morita, H; Sato, S; Ishii, T. Expression of secretory phospholipase A2s in human atherosclerosis development. Atherosclerosis 2008, 196, 81–91. [Google Scholar]
- Sato, H; Taketomi, Y; Isogai, Y; Miki, Y; Yamamoto, K; Masuda, S; Hosono, T; Arata, S; Ishikawa, Y; Ishii, T; Kobayashi, T; Nakanishi, H; Ikeda, K; Taguchi, R; Hara, S; Kudo, I; Murakami, M. Group III secreted phospholipase A2 regulates epididymal sperm maturation and fertility in mice. J. Clin. Invest 2010, 120, 1400–1414. [Google Scholar]
- Cooper, TG. Role of the epididymis in mediating changes in the male gamete during maturation. Adv. Exp. Med. Biol 1995, 377, 87–101. [Google Scholar]
- Rejraji, H; Sion, B; Prensier, G; Carreras, M; Motta, C; Frenoux, JM; Vericel, E; Grizard, G; Vernet, P; Drevet, JR. Lipid remodeling of murine epididymosomes and spermatozoa during epididymal maturation. Biol. Reprod 2006, 74, 1104–1113. [Google Scholar]
- Lenzi, A; Picardo, M; Gandini, L; Dondero, F. Lipids of the sperm plasma membrane: from polyunsaturated fatty acids considered as markers of sperm function to possible scavenger therapy. Hum. Reprod 1996, 2, 246–256. [Google Scholar]
- Lenzi, A; Gandini, L; Maresca, V; Rago, R; Sgrò, P; Dondero, F; Picardo, M. Fatty acid composition of spermatozoa and immature germ cells. Mol. Hum. Reprod 2000, 6, 226–231. [Google Scholar]
- Furimsky, A; Vuong, N; Xu, H; Kumarathasan, P; Xu, M; Weerachatyanukul, W; Bou, KM; Kates, M; Tanphaichitr, N. Percoll gradient-centrifuged capacitated mouse sperm have increased fertilizing ability and higher contents of sulfogalactosylglycerolipid and docosahexaenoic acid-containing phosphatidylcholine compared to washed capacitated mouse sperm. Biol. Reprod 2005, 72, 574–583. [Google Scholar]
- Aksoy, Y; Aksoy, H; Altinkaynak, K; Aydin, HR; Ozkan, A. Sperm fatty acid composition in subfertile men. Prostag. Leuk. Essent. Fatty 2006, 75, 75–79. [Google Scholar]
- Hall, JC; Hadley, J; Doman, T. Correlation between changes in rat sperm membrane lipids, protein, and the membrane physical state during epididymal maturation. J. Androl 1991, 12, 76–87. [Google Scholar]
- Haidl, G; Opper, C. Changes in lipids and membrane anisotropy in human spermatozoa during epididymal maturation. Hum. Reprod 1997, 12, 2720–2723. [Google Scholar]
- Fischer, KA; Van Leyen, K; Lovercamp, KW; Manandhar, G; Sutovsky, M; Feng, D; Safranski, T; Sutovsky, P. 15-Lipoxygenase is a component of the mammalian sperm cytoplasmic droplet. Reproduction 2005, 130, 213–222. [Google Scholar]
- Law, GL; McGuinness, MP; Linder, CC; Griswold, MD. Expression of apolipoprotein E mRNA in the epithelium and interstitium of the testis and the epididymis. J. Androl 1997, 18, 32–42. [Google Scholar]
- Hermo, L; Wright, J; Oko, R; Morales, CR. Role of epithelial cells of the male excurrent duct system of the rat in the endocytosis or secretion of sulfated glycoprotein-2 (clusterin). Biol. Reprod 1991, 44, 1113–1131. [Google Scholar]
- Huang, LS; Voyiaziakis, E; Chen, HL; Rubin, EM; Gordon, JW. A novel functional role for apolipoprotein B in male infertility in heterozygous apolipoprotein B knockout mice. Proc. Natl. Acad. Sci. USA 1996, 93, 10903–10907. [Google Scholar]
- Moghadasian, MH; Nguyen, LB; Shefer, S; McManus, BM; Frohlich, JJ. Histologic, hematologic, and biochemical characteristics of apo E-deficient mice: effects of dietary cholesterol and phytosterols. Lab. Invest 1999, 79, 355–364. [Google Scholar]
- Andersen, OM; Yeung, CH; Vorum, H; Wellner, M; Andreassen, TK; Erdmann, B; Mueller, EC; Herz, J; Otto, A; Cooper, TG; Willnow, TE. Essential role of the apolipoprotein E receptor-2 in sperm development. J. Biol. Chem 2003, 278, 23989–23995. [Google Scholar]
- Saez, F; Frenette, G; Sullivan, R. Epididymosomes and prostasomes: their roles in posttesticular maturation of the sperm cells. J. Androl 2003, 24, 149–154. [Google Scholar]
- Masuda, S; Murakami, M; Matsumoto, S; Eguchi, N; Urade, Y; Lambeau, G; Gelb, MH; Ishikawa, Y; Ishii, T; Kudo, I. Localization of various secretory phospholipase A2 enzymes in male reproductive organs. Biochim. Biophys. Acta 2004, 1686, 61–76. [Google Scholar]
- Seilhamer, JJ; Randall, TL; Yamanaka, M; Johnson, LK. Pancreatic phospholipase A2: Isolation of the human gene and cDNAs from porcine pancreas and human lung. DNA 1986, 5, 519–527. [Google Scholar]
- Labonte, ED; Kirby, RJ; Schildmeyer, NM; Cannon, AM; Huggins, KW; Hui, DY. Group IB phospholipase A2-mediated lysophospholipid absorption directly contributes to postprandial hyperglycemia. Diabetes 2006, 55, 935–941. [Google Scholar]
- Hui, DY; Cope, MJ; Labonte, ED; Chang, HT; Shao, J; Goka, E; Abousalham, A; Charmot, D; Buysse, J. The phospholipase A2 inhibitor methyl indoxam suppresses diet-induced obesity and glucose intolerance in mice. Br. J. Pharmacol 2009, 157, 1263–1269. [Google Scholar]
- Wilson, SG; Adam, G; Langdown, M; Reneland, R; Braun, A; Andrew, T; Surdulescu, GL; Norberg, M; Dudbridge, F; Reed, PW; Sambrook, PN; Kleyn, PW; Spector, TD. Linkage and potential association of obesity-related phenotypes with two genes on chromosome 12q24 in a female dizygous twin cohort. Eur. J. Hum Genet 2006, 14, 340–348. [Google Scholar]
- Pruzanski, W; Vadas, P. Phospholipase A2—A mediator between proximal and distal effectors of inflammation. Immunol. Today 1991, 12, 143–146. [Google Scholar]
- Murakami, M; Kambe, T; Shimbara, S; Yamamoto, S; Kuwata, H; Kudo, I. Functional association of type IIA secretory phospholipase A2 with the glycosylphosphatidylinositol-anchored heparan sulfate proteoglycan in the cyclooxygenase-2-mediated delayed prostanoid-biosynthetic pathway. J. Biol. Chem 1999, 274, 29927–29936. [Google Scholar]
- MacPhee, M; Chepenik, PK; Liddel, AR; Nelson, KK; Siracusa, DL; Buchberg, MA. The secretory phospholipase A2 gene is a candidate for the Mom1 locus, a major modifier of Apcmin-induced intestinal neoplasia. Cell 1995, 81, 957–966. [Google Scholar]
- Cormier, RT; Hong, KH; Halberg, RB; Hawkins, TL; Richardson, P; Mulherkar, R; Dove, WF; Lander, ES. Secretory phospholipase Pla2g2a confers resistance to intestinal tumorigenesis. Nat. Genet 1997, 17, 88–91. [Google Scholar]
- Weinrauch, Y; Elsbach, P; Madsen, LM; Foreman, A; Weiss, J. The potent anti-Staphylococcus aureus activity of a sterile rabbit inflammatory fluid is due to a 14-kD phospholipase A2. J. Clin. Invest 1996, 97, 250–257. [Google Scholar]
- Weinrauch, Y; Abad, C; Liang, NS; Lowry, SF; Weiss, J. Mobilization of potent plasma bactericidal activity during systemic bacterial challenge. Role of group IIA phospholipase A2. J. Clin. Invest 1998, 102, 633–638. [Google Scholar]
- Laine, VJ; Grass, DS; Nevalainen, TJ. Protection by group II phospholipase A2 against Staphylococcus aureus. J. Immunol 1999, 162, 7402–7408. [Google Scholar]
- Koduri, RS; Grönroos, JO; Laine, VJ; Le Calvez, C; Lambeau, G; Nevalainen, TJ; Gelb, MH. Bactericidal properties of human and murine groups I, II, V, X, and XII secreted phospholipases A2. J. Biol. Chem 2002, 277, 5849–5857. [Google Scholar]
- Kugiyama, K; Ota, Y; Takazoe, K; Moriyama, Y; Kawano, H; Miyao, Y; Sakamoto, T; Soejima, H; Ogawa, H; Doi, H; Sugiyama, S; Yasue, H. Circulating levels of secretory type II phospholipase A2 predict coronary events in patients with coronary artery disease. Circulation 1999, 100, 1280–1284. [Google Scholar]
- Ivandic, B; Castellani, LW; Wang, XP; Qiao, JH; Mehrabian, M; Navab, M; Fogelman, AM; Grass, DS; Swanson, ME; De Beer, MC; De Beer, F; Lusis, AJ. Role of group II secretory phospholipase A2 in atherosclerosis: 1. Increased atherogenesis and altered lipoproteins in transgenic mice expressing group IIa phospholipase A2. Arterioscler. Thromb. Vasc. Biol 1999, 19, 1284–1290. [Google Scholar]
- Webb, NR; Bostrom, MA; Szilvassy, SJ; Van der Westhuyzen, DR; Daugherty, A; De Beer, FC. Macrophage-expressed group IIA secretory phospholipase A2 increases atherosclerotic lesion formation in LDL receptor-deficient mice. Arterioscler. Thromb. Vasc. Biol 2003, 23, 263–268. [Google Scholar]
- Chen, J; Shao, C; Lazar, V; Srivastava, CH; Lee, WH; Tischfield, JA. Localization of group IIc low molecular weight phospholipase A2 mRNA to meiotic cells in the mouse. J. Cell Biochem 1997, 64, 369–375. [Google Scholar]
- Shakhov, AN; Rubtsov, AV; Lyakhov, IG; Tumanov, AV; Nedospasov, SA. SPLASH (PLA2IID), a novel member of phospholipase A2 family, is associated with lymphotoxin deficiency. Genes Immun 2000, 1, 191–199. [Google Scholar]
- Von Allmen, CE; Schmitz, N; Bauer, M; Hinton, HJ; Kurrer, MO; Buser, RB; Gwerder, M; Muntwiler, S; Sparwasser, T; Beerli, RR; Bachmann, MF. Secretory phospholipase A2-IID is an effector molecule of CD4+CD25+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 11673–11678. [Google Scholar]
- Suzuki, N; Ishizaki, J; Yokota, Y; Higashino, K; Ono, T; Ikeda, M; Fujii, N; Kawamoto, K; Hanasaki, K. Structures, enzymatic properties, and expression of novel human and mouse secretory phospholipases A2. J. Biol. Chem 2000, 275, 5785–5793. [Google Scholar]
- Valentin, E; Ghomashchi, F; Gelb, MH; Lazdunski, M; Lambeau, G. On the diversity of secreted phospholipases A2. Cloning, tissue distribution, and functional expression of two novel mouse group II enzymes. J. Biol. Chem 1999, 274, 31195–31202. [Google Scholar]
- Murakami, M; Yoshihara, K; Shimbara, S; Lambeau, G; Gelb, MH; Singer, AG; Sawada, M; Inagaki, N; Nagai, H; Ishihara, M; Ishikawa, Y; Ishii, T; Kudo, I. Cellular arachidonate-releasing function and inflammation-associated expression of group IIF secretory phospholipase A2. J. Biol. Chem 2002, 277, 19145–19155. [Google Scholar]
- Gelb, MH; Valentin, E; Ghomashchi, F; Lazdunski, M; Lambeau, G. Cloning and recombinant expression of a structurally novel human secreted phospholipase A2. J. Biol. Chem 2000, 275, 39823–39826. [Google Scholar]
- Munoz-Sanjuan, I; Brivanlou, AH. Induction of ectopic olfactory structures and bone morphogenetic protein inhibition by Rossy, a group XII secreted phospholipase A2. Mol. Cell Biol 2005, 25, 3608–3619. [Google Scholar]
- Rouault, M; Bollinger, JG; Lazdunski, M; Gelb, MH; Lambeau, G. Novel mammalian group XII secreted phospholipase A2 lacking enzymatic activity. Biochemistry 2003, 42, 11494–11503. [Google Scholar]
- Guan, M; Qu, L; Tan, W; Chen, L; Wong, CW. Hepatocyte nuclear factor-4. regulates liver triglyceride metabolism in part through secreted phospholipase A2 GXIIB. Hepatology 2011, in press. [Google Scholar]
- Rosenson, RS. Phospholipase A2 inhibition and atherosclerotic vascular disease: prospects for targeting secretory and lipoprotein-associated phospholipase A2 enzymes. Curr. Opin. Lipidol 2010, 21, 473–480. [Google Scholar]
- Yamamoto, K; Takemomi, Y; Miki, Y; Shimo, K; Nakanishi, H; Ikeda, K; Taguchi, R; Gelb, MH; Murakami, M. The Tokyo Metropolitan Institute of Medical Science; Tokyo, Japan, 2010; unpublished results. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sPLA2s | resting cell membrane | activated cell membrane | lipoprotein (PC) | surfactant (PC) | Gram-positive bacteria | Gram-negative bacteria | |
|---|---|---|---|---|---|---|---|
| conventional sPLA2s | IB | weak | moderate | weak | weak | none | none |
| IIA | none | moderate | weak | weak | very high | weak* | |
| IID | none | weak | n.d. | weak | high | none | |
| IIE | none | weak | n.d. | n.d. | moderate | none | |
| IIF | moderate | moderate | moderate | n.d. | none | none | |
| V | high | very high | very high | very high | high | none | |
| X | very high | very high | very high | high | moderate | none | |
| atypical sPLA2s | III | moderate | moderate | high | n.d. | n.d. | none |
| XIIA | none | none | n.d. | n.d. | high | moderate |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Murakami, M.; Sato, H.; Taketomi, Y.; Yamamoto, K. Integrated Lipidomics in the Secreted Phospholipase A2 Biology. Int. J. Mol. Sci. 2011, 12, 1474-1495. https://doi.org/10.3390/ijms12031474
Murakami M, Sato H, Taketomi Y, Yamamoto K. Integrated Lipidomics in the Secreted Phospholipase A2 Biology. International Journal of Molecular Sciences. 2011; 12(3):1474-1495. https://doi.org/10.3390/ijms12031474
Chicago/Turabian StyleMurakami, Makoto, Hiroyasu Sato, Yoshitaka Taketomi, and Kei Yamamoto. 2011. "Integrated Lipidomics in the Secreted Phospholipase A2 Biology" International Journal of Molecular Sciences 12, no. 3: 1474-1495. https://doi.org/10.3390/ijms12031474