Cancer Stem-Like Cells Enriched in Panc-1 Spheres Possess Increased Migration Ability and Resistance to Gemcitabine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. CSC Population of Panc-1 Cells Is Enriched under Sphere-Forming Conditions
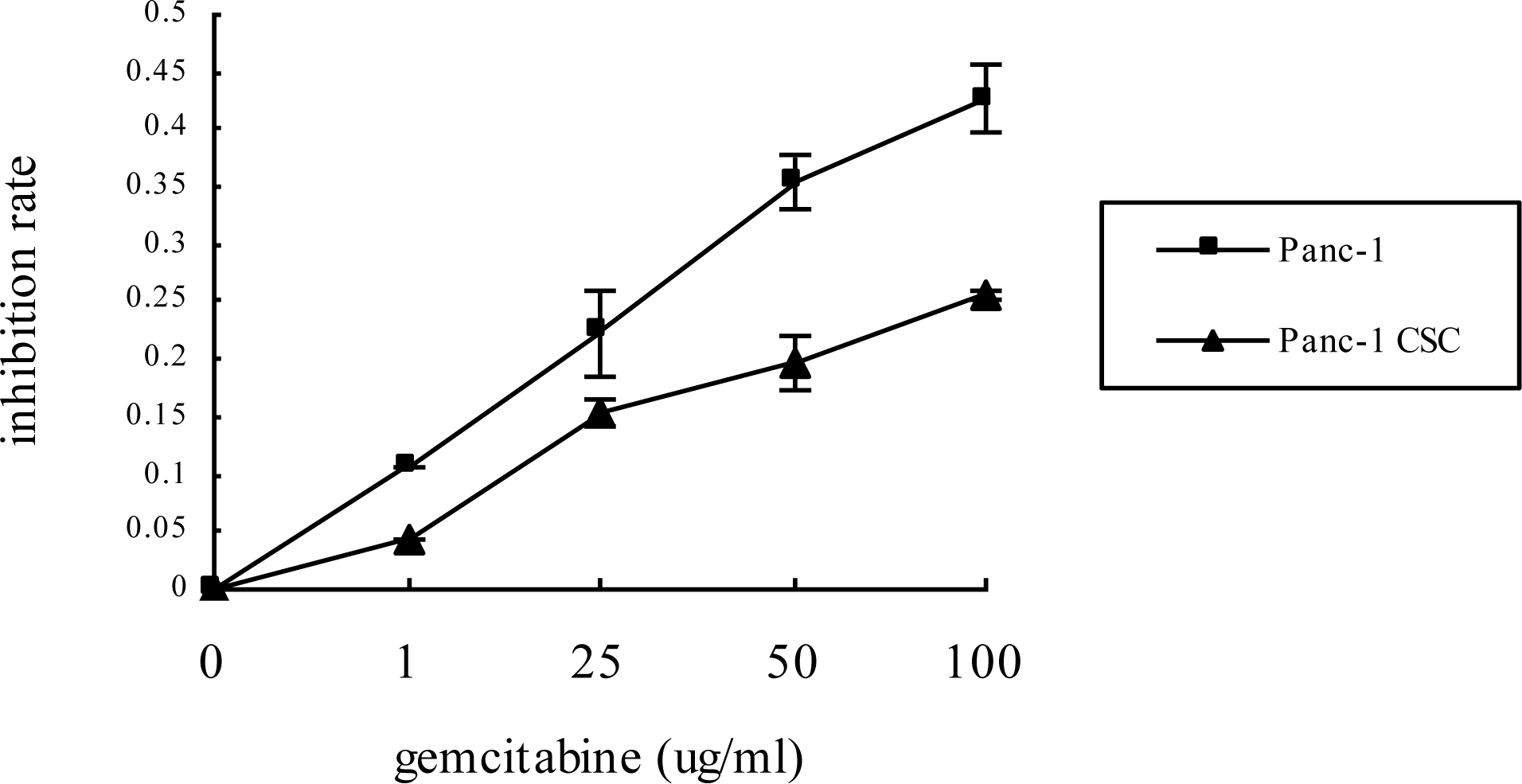
2.2. Panc-1 CSCs Exhibit Increased Chemoresistance to Gemcitabine
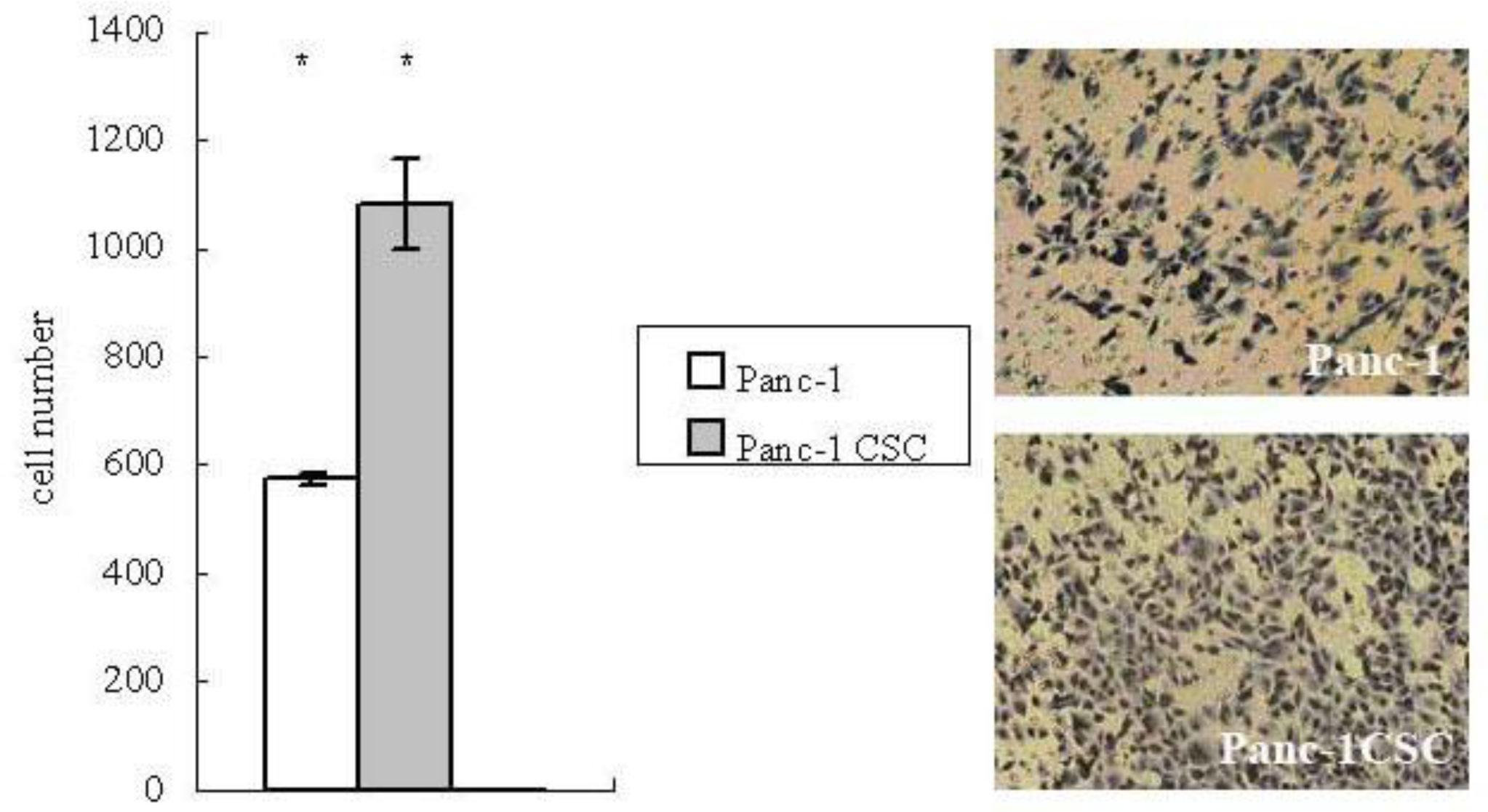
2.3. Panc-1 CSCs Exhibit Increased Migration Ability
2.4. Panc-1 CSCs Exhibit Epithelial to Mesenchymal Transition (EMT)
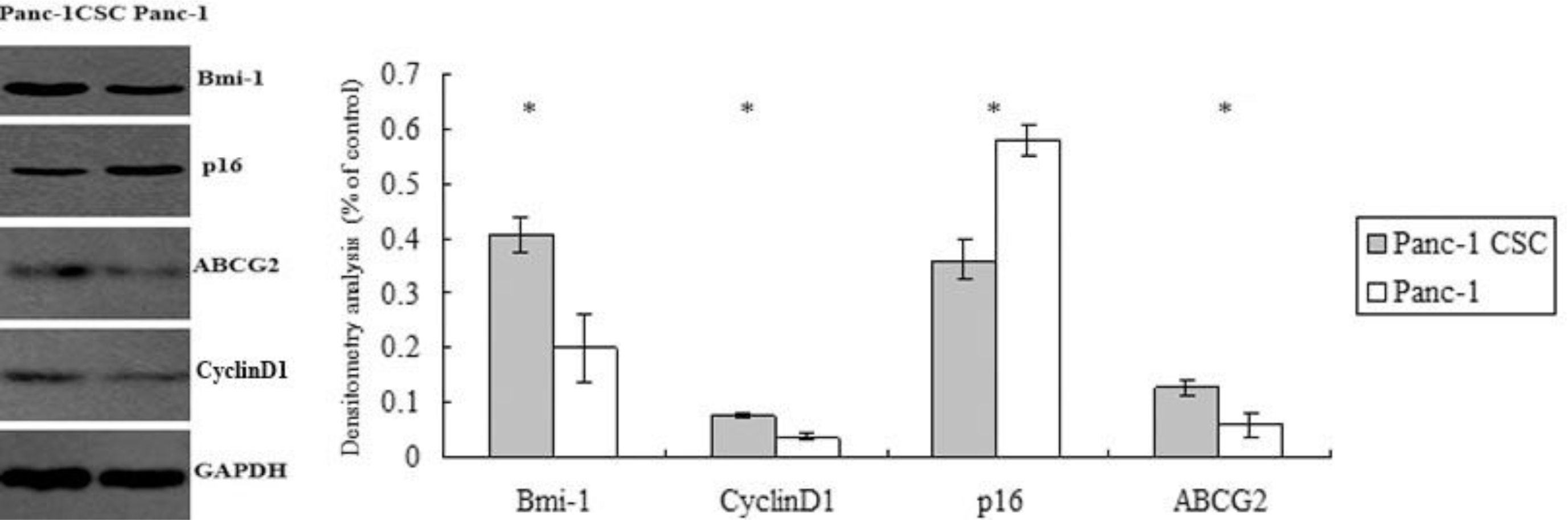
2.5. Abnormal Expression of Bmi-1, ABCG2, Cyclin D1 and p16 in Panc-1 CSCs
2.6. Discussion
3. Experimental Section
3.1. Cell Lines and Cell Culture
3.2. Flow Cytometry Assay
3.3. MTT Cell Proliferation Assay
3.4. Transwell Migration Assay
3.5. Western Blot Analysis
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Jemal, A; Tiwari, RC; Murray, T. Cancer statistics, 2004. CA Cancer J. Clin 2004, 54, 8–29. [Google Scholar]
- Polyak, K; Hahn, WC. Roots and stems, stem cells in cancer. Nat. Med 2006, 12, 296–300. [Google Scholar]
- Li, F; Tiede, B; Massagué, J; Kang, Y. Beyond tumorigenesis, cancer stem cells in metastasis. Cell Res 2007, 17, 3–14. [Google Scholar]
- Kakarala, M; Wicha, MS. Implications of the cancer stem-cell hypothesis for breast cancer prevention and therapy. J. Clin. Oncol 2008, 26, 2813–2820. [Google Scholar]
- Singh, SK; Clarke, ID; Terasaki, M; Bonn, VE; Hawkins, C; Squire, J; Dirks, PB. Identification of a cancer stem cell in human brain tumors. Cancer Res 2003, 63, 5821–5828. [Google Scholar]
- Ricci-Vitiani, L; Lombardi, DG; Pilozzi, E; Biffoni1, M; Todaro, M; Peschle1, C; De Maria1, R. Identification and expansion of human colon-cancer-initiating cells. Nature 2007, 445, 111–115. [Google Scholar]
- Ponti, D; Costa, A; Zaffaroni, N; Pratesi, G; Petrangolini, G; Coradini, D; Pilotti, S; Pierotti, MA; Daidone, MG. Isolation and in vitro propagation of tumorigenic breast cancer cells with stem/progenitor cell properties. Cancer Res 2005, 65, 5506–5511. [Google Scholar]
- Bonnet, D; Dick, JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat. Med 1997, 3, 730–737. [Google Scholar]
- Li, C; Heidt, DG; Dalerba, P; Burant, CF; Zhang, L; Adsay, V; Wicha, M; Clarke, MF; Simeone, DM. Identification of pancreatic cancer stem cells. Cancer Res 2007, 67, 1030–1037. [Google Scholar]
- Gou, S; Liu, T; Wang, C; Yin, T; Li, K; Yang, M; Zhou, J. Establishment of clonal colony-forming assay for propagation of pancreatic cancer cells with stem cell properties. Pancreas 2007, 34, 429–435. [Google Scholar]
- Huang, P; Wang, CY; Gou, SM; Wu, H-S; Liu, T; Xiong, J-X. Isolation and biological analysis of tumor stem cells from pancreatic adenocarcinoma. World J. Gastroenterol 2008, 14, 3903–3907. [Google Scholar]
- Balic, M; Lin, H; Young, L; Hawes, D; Giuliano, A; McNamara, G; Datar, RH; Cote1, RJ. Most early disseminated cancer cells detected in bone marrow of breast cancer patients have a putative breast cancer stem cell phenotype. Clin. Cancer Res 2006, 12, 5615–5621. [Google Scholar]
- Thiery, JP. Epithelial-mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar]
- Polyak, K; Hahn, WC. Roots and stems, stem cells in cancer. Nat. Med 2006, 12, 296–300. [Google Scholar]
- Ischenko, I; Seeliger, H; Schaffer, M. Cancer stem cells, how can we target them? Curr. Med. Chem 2008, 15, 3171–3184. [Google Scholar]
- Dylla, SJ; Beviglia, L; Park, IK; Chartier, C; Raval, J; Ngan, L; Pickell, K; Aguilar, J; Lazetic, S; Smith-Berdan, S; et al. Colorectal cancer stem cells are enriched in xenogeneic tumors following chemotherapy. PLoS One 2008, 3, e2428:1–e2428:13. [Google Scholar]
- Rizo, A; Olthof, S; Han, L; Vellenga, E; de Haan, G; Schuringa, JJ. Repression of BMI1 in normal and leukemic human CD34(+) cells impairs self-renewal and induces apoptosis. Blood 2009, 114, 1498–1505. [Google Scholar]
- Datta, S; Hoenerhoff, MJ; Bommi, P; Sainger, R; Guo, WJ; Dimri, M; Band, H; Band, V; Green, JE; Dimri, GP. Bmi-1 cooperates with H-Ras to transform human mammary epithelial cells via dysregulation of multiple growth-regulatory pathways. Cancer Res 2007, 67, 10286–10295. [Google Scholar]
- Song, W; Tao, K; Li, H; Jin, C; Song, Z; Li, J; Shi, H; Li, X; Dang, Z; Dou, K. Bmi-1 is related to proliferation, survival and poor prognosis in pancreatic cancer. Cancer Sci 2010, 101, 1754–1760. [Google Scholar]
- Ejendal, KF; Hrycyna, CA. Multidrug resistance and cancer, the role of the human ABC transporter ABCG2. Curr. Protein Pept. Sci 2002, 3, 503–511. [Google Scholar]
- Kim, M; Turnquist, H; Jackson, J; Sgagias, M; Yan, Y; Gong, M; Dean, M; Sharp, JG; Cowan, K. The multidrug resistance transporter ABCG2 (breast cancer resistance protein 1effluxes Hoechst 33342 and is overexpressed in hematopoietic stem cells. Clin. Cancer Res 2002, 8, 22–28. [Google Scholar]
- Attri, J; Srinivasan, R; Majumdar, S; Radotra, BD; Wig, J. Alterations of tumor suppressor gene p16INK4a in pancreatic ductal carcinoma. BMC Gastroenterol 2005, 5, 22:1–22:10. [Google Scholar]
- Halloran, CM; Ghaneh, P; Shore, S; Greenhalf, W; Zumstein, L; Wilson, D; Neoptolemos, JP; Costello, E. 5-Fluorouracil or gemcitabine combined with adenoviral-mediated reintroduction of p16INK4A greatly enhanced cytotoxicity in Panc-1 pancreatic adenocarcinoma cells. J. Gene Med 2004, 6, 514–525. [Google Scholar]
- Gansauge, S; Gansauge, F; Ramadani, M; Stobbe, H; Rau, B; Harada, N; Beger, HG. Overexpression of cyclin D1 in human pancreatic carcinoma is associated with poor prognosis. Cancer Res 1997, 57, 1634–1637. [Google Scholar]
- Biliran, H, Jr; Wang, Y; Banerjee, S; Xu, H; Heng, H; Thakur, A; Bollig, A; Sarkar, FH; Liao, JD. Overexpression of cyclin D1 promotes tumor cell growth and confers resistance to cisplatin-mediated apoptosis in an elastase-myc transgene-expressing pancreatic tumor cell line. Clin. Cancer Res 2005, 11, 6075–6086. [Google Scholar]
- Jeselsohn, R; Brown, NE; Arendt, L; Klebba, I; Hu, MG; Kuperwasser, C; Hinds, PW. Cyclin D1 kinase activity is required for the self-renewal of mammary stem and progenitor cells that are targets of MMTV-ErbB2 tumorigenesis. Cancer Cell 2010, 17(1), 65–76. [Google Scholar]
- Mani, SA; Guo, W; Liao, MJ; Eaton, EN; Ayyanan, A; Zhou1, AY; Brooks, M; Reinhard, F; Zhang, CC; Shipitsin, M; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar]
- Larue, L; Bellacosa, A. Epithelial-mesenchymal transition in development and cancer, role of phosphatidylinositol 3′ kinase/AKT pathways. Oncogene 2005, 24, 7443–7454. [Google Scholar]
- Robson, EJ; Khaled, WT; Abell, K; Watson, CJ. Epithelial-to-mesenchymal transition confers resistance to apoptosis in three murine mammary epithelial cell lines. Differentiation 2006, 74, 254–264. [Google Scholar]
- Lee, JM; Dedhar, S; Kalluri, R; Thompson, EW. The epithelial-mesenchymal transition, new insights in signaling, development, and disease. J. Cell Biol 2006, 172, 973–981. [Google Scholar]





© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yin, T.; Wei, H.; Gou, S.; Shi, P.; Yang, Z.; Zhao, G.; Wang, C. Cancer Stem-Like Cells Enriched in Panc-1 Spheres Possess Increased Migration Ability and Resistance to Gemcitabine. Int. J. Mol. Sci. 2011, 12, 1595-1604. https://doi.org/10.3390/ijms12031595
Yin T, Wei H, Gou S, Shi P, Yang Z, Zhao G, Wang C. Cancer Stem-Like Cells Enriched in Panc-1 Spheres Possess Increased Migration Ability and Resistance to Gemcitabine. International Journal of Molecular Sciences. 2011; 12(3):1595-1604. https://doi.org/10.3390/ijms12031595
Chicago/Turabian StyleYin, Tao, Hongji Wei, Shanmiao Gou, Pengfei Shi, Zhiyong Yang, Gang Zhao, and Chunyou Wang. 2011. "Cancer Stem-Like Cells Enriched in Panc-1 Spheres Possess Increased Migration Ability and Resistance to Gemcitabine" International Journal of Molecular Sciences 12, no. 3: 1595-1604. https://doi.org/10.3390/ijms12031595