Esculetin Ameliorates Carbon Tetrachloride-Mediated Hepatic Apoptosis in Rats

Abstract
:1. Introduction
2. Results and Discussion
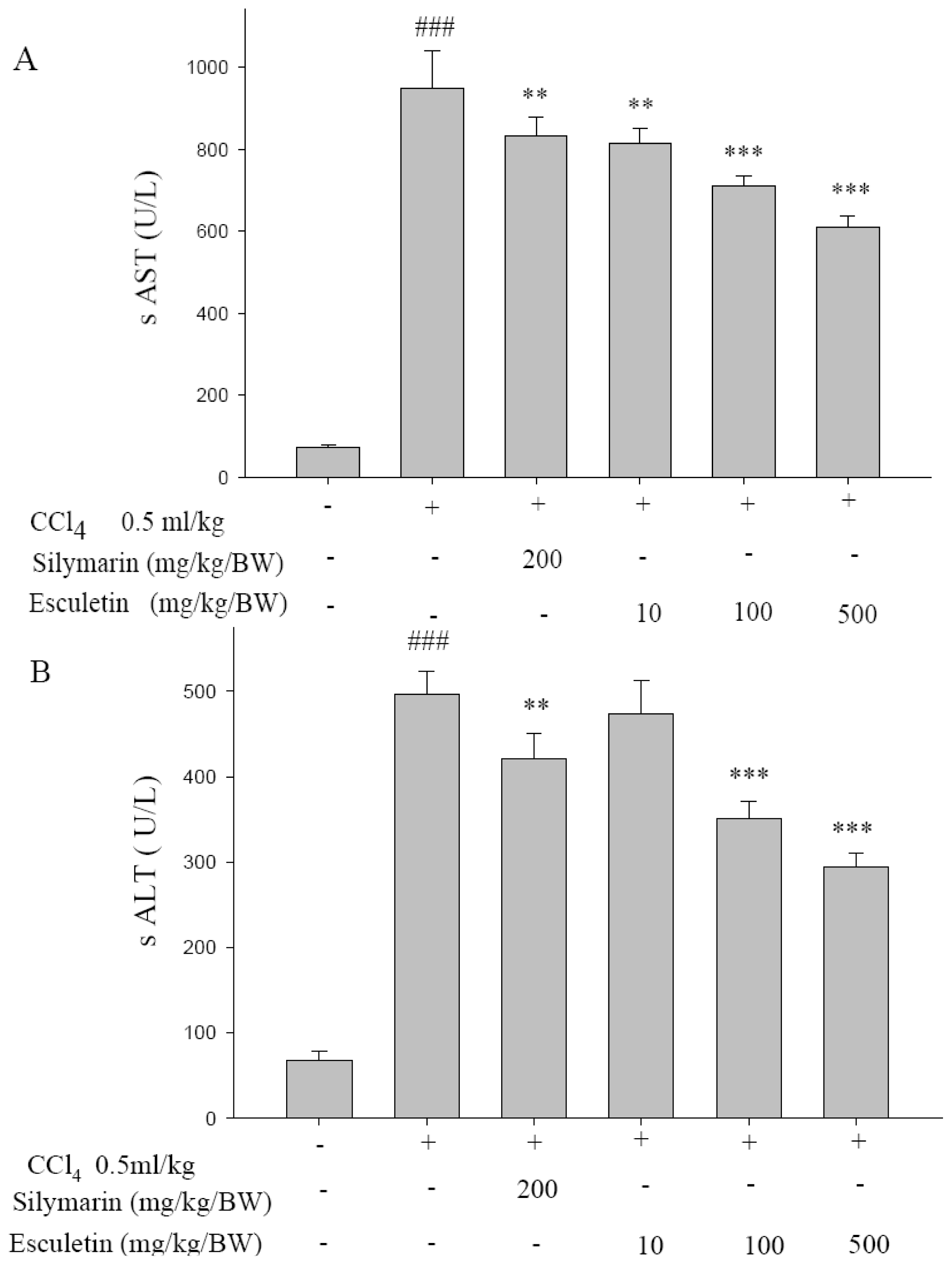
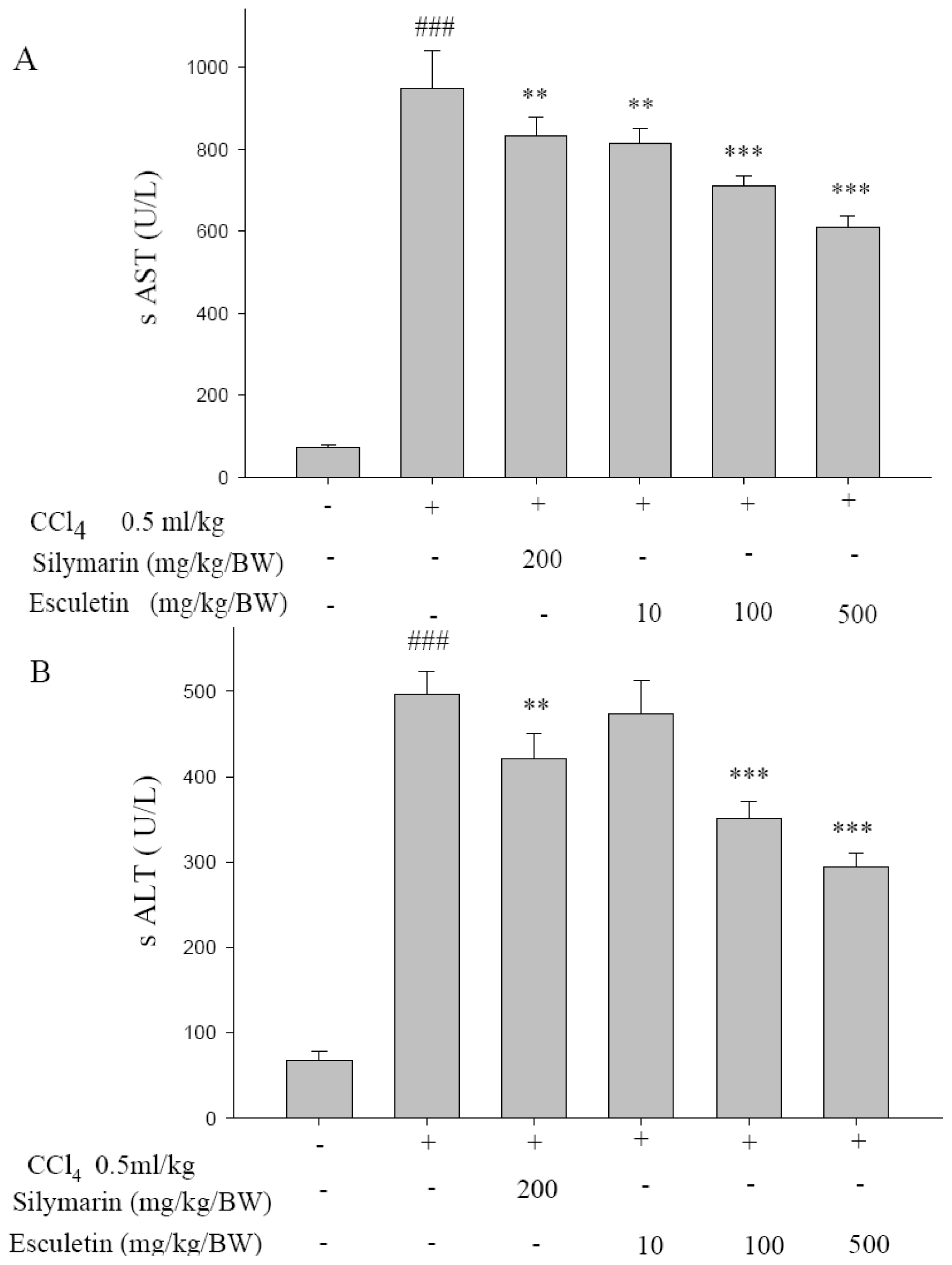
2.1. Serum ALT and AST Activities
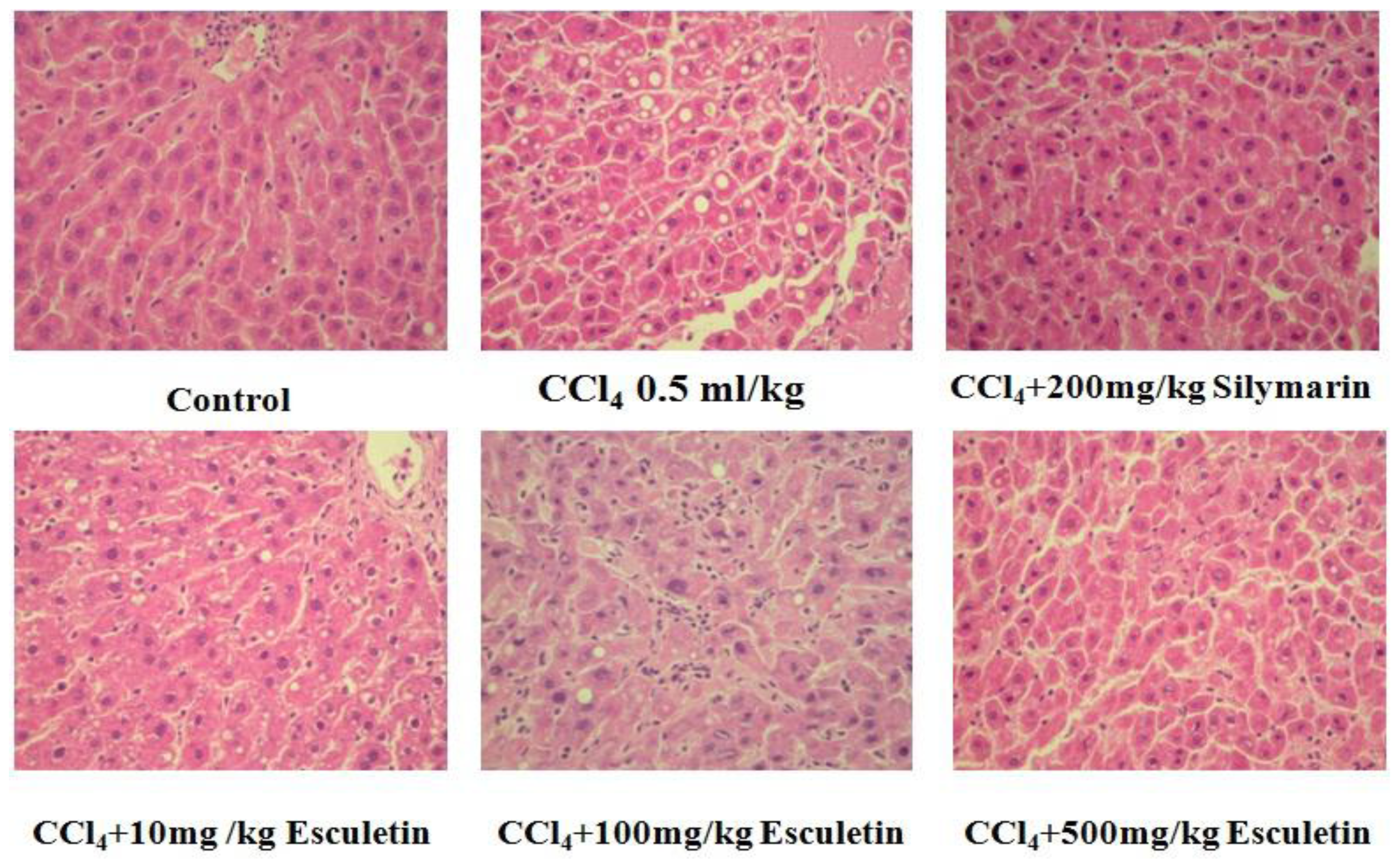
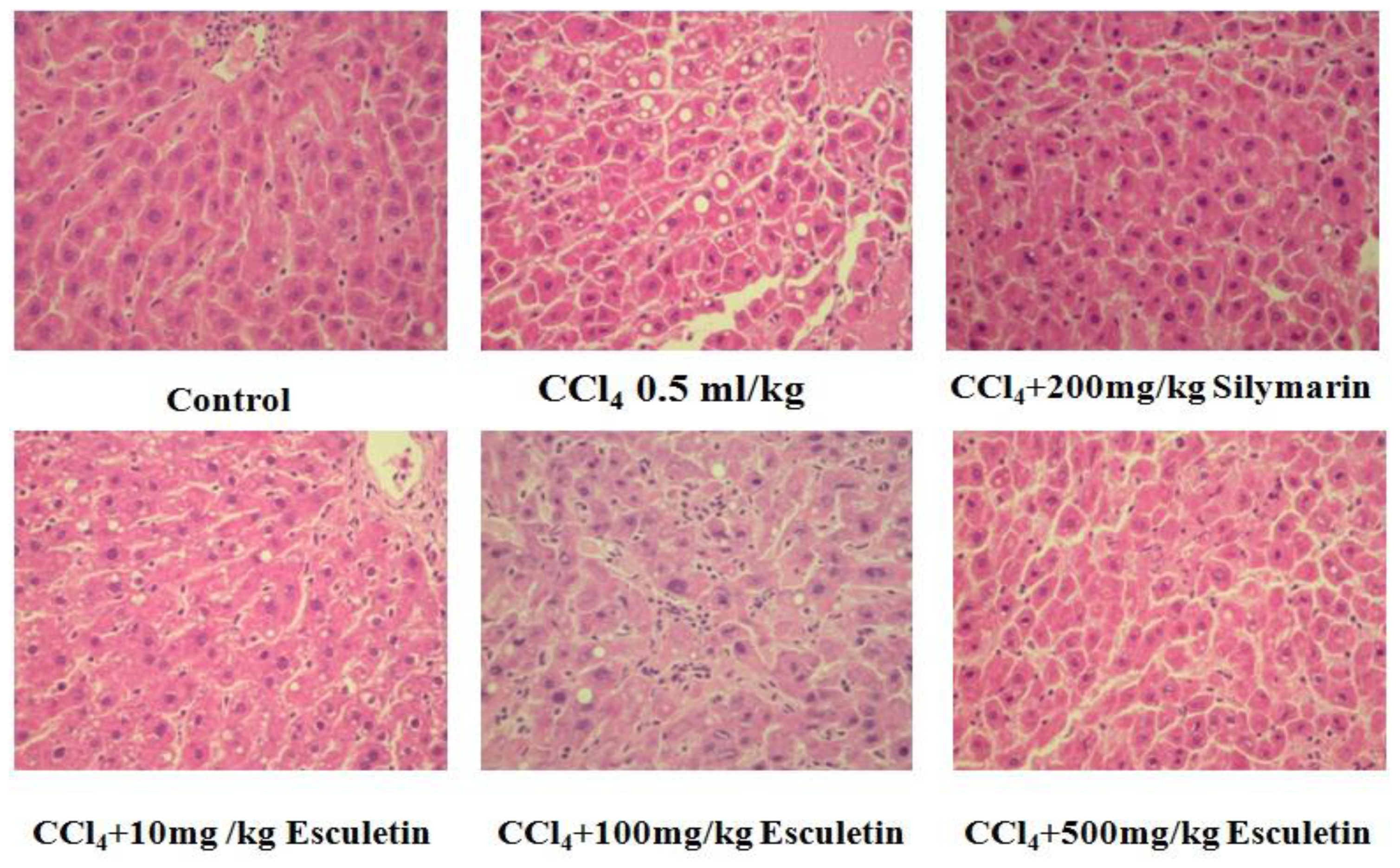
2.2. Changes in Liver Histopathology
2.3. Hepatic Catalase, SOD and GPx Activities
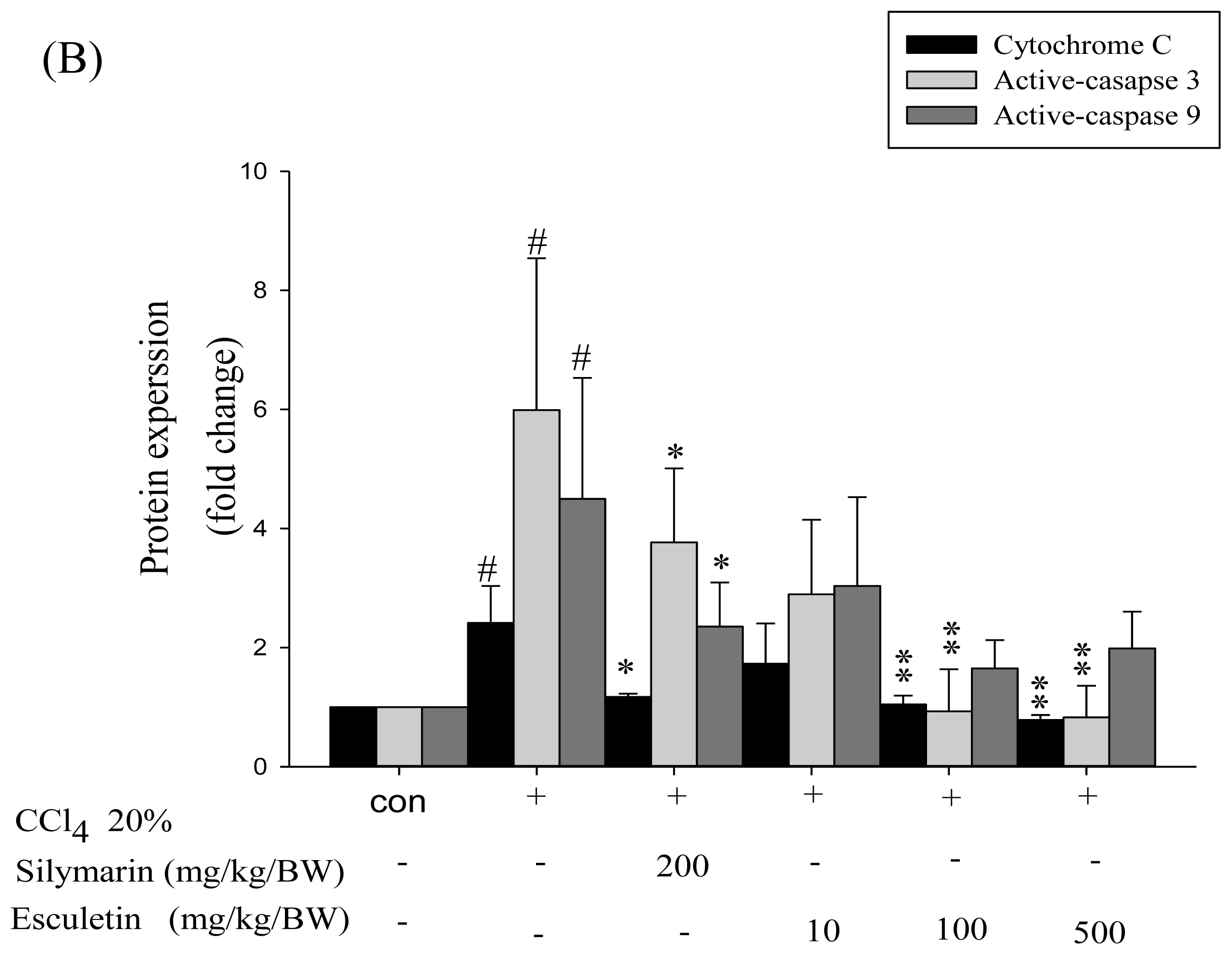
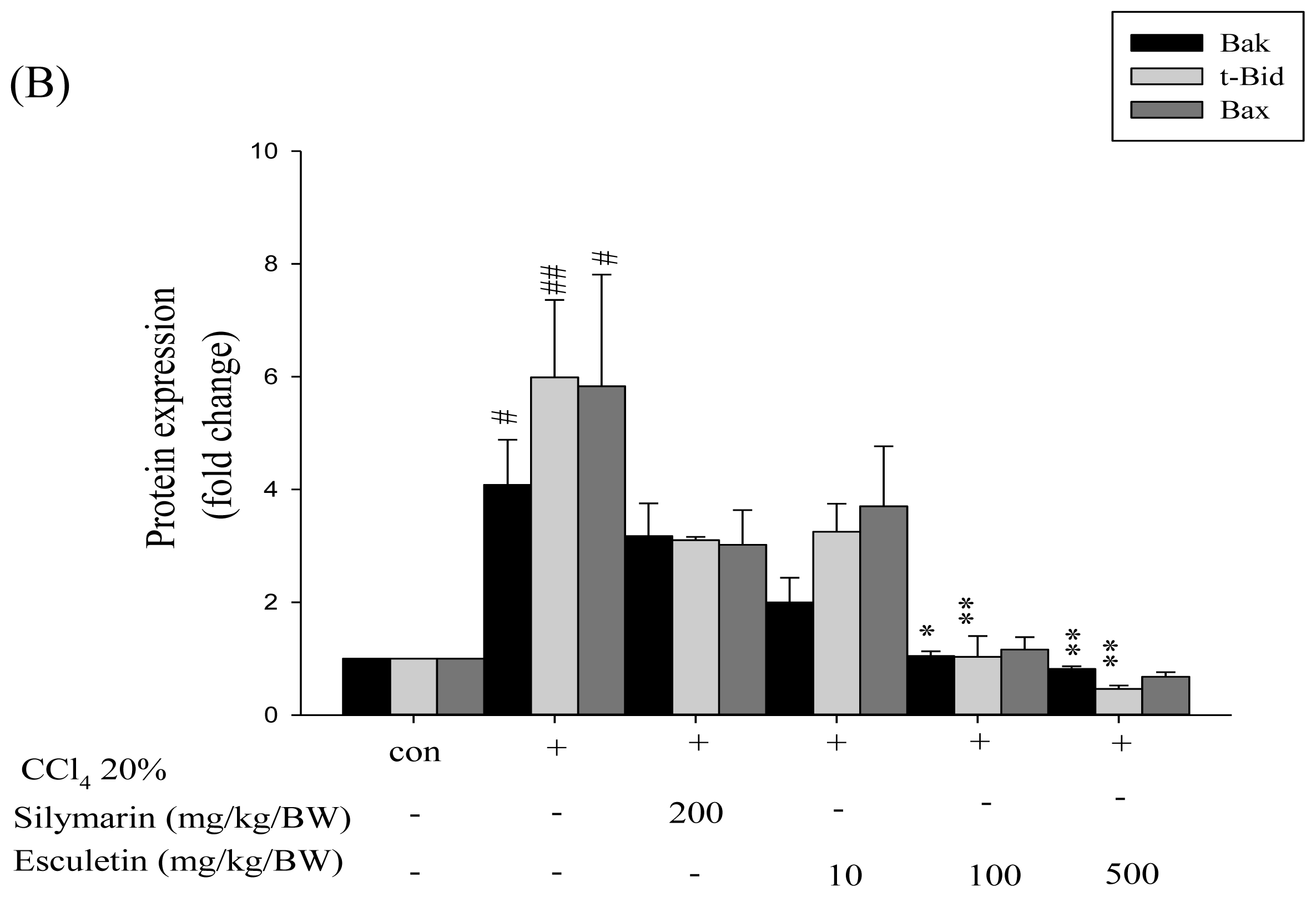
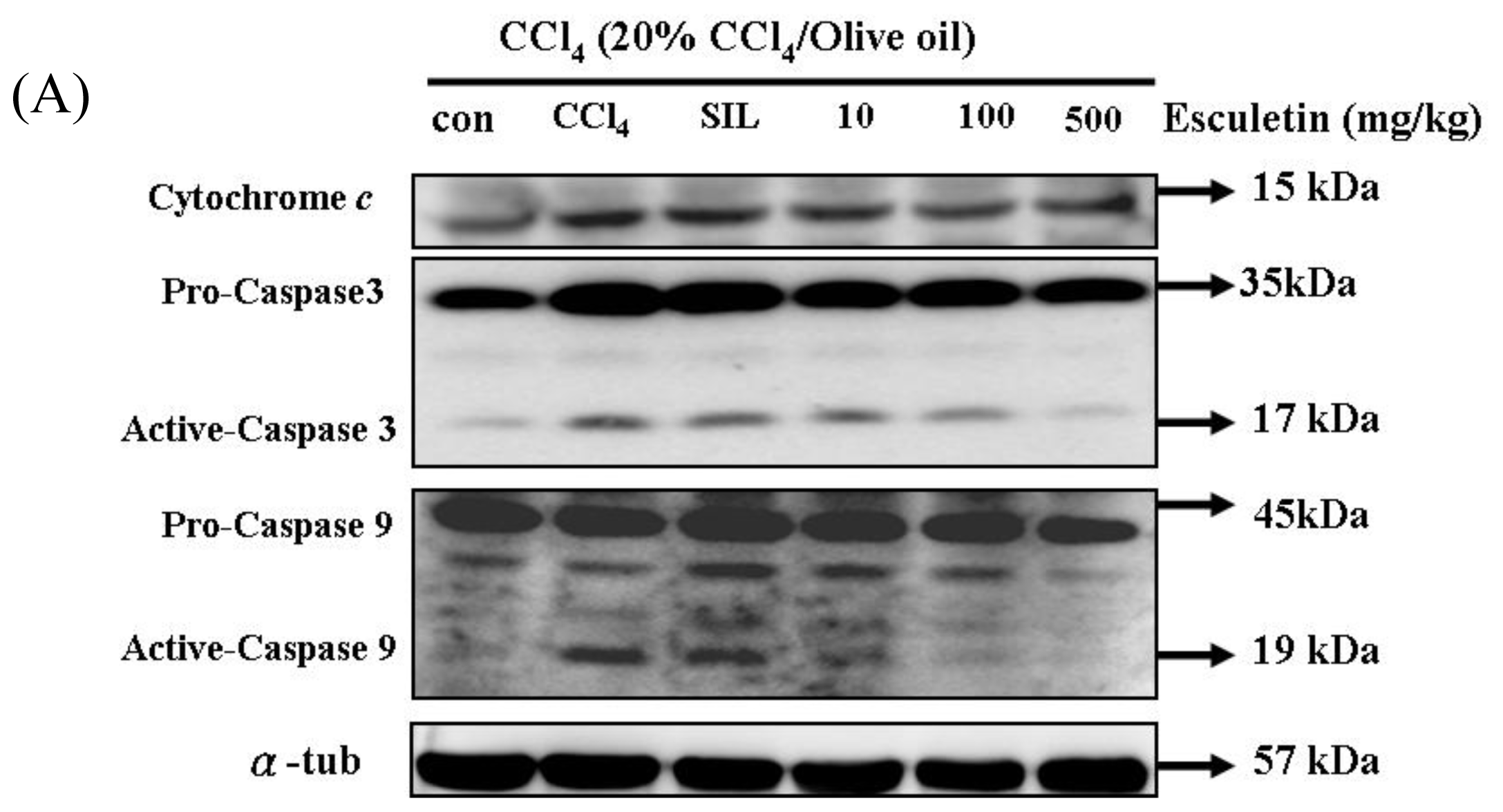
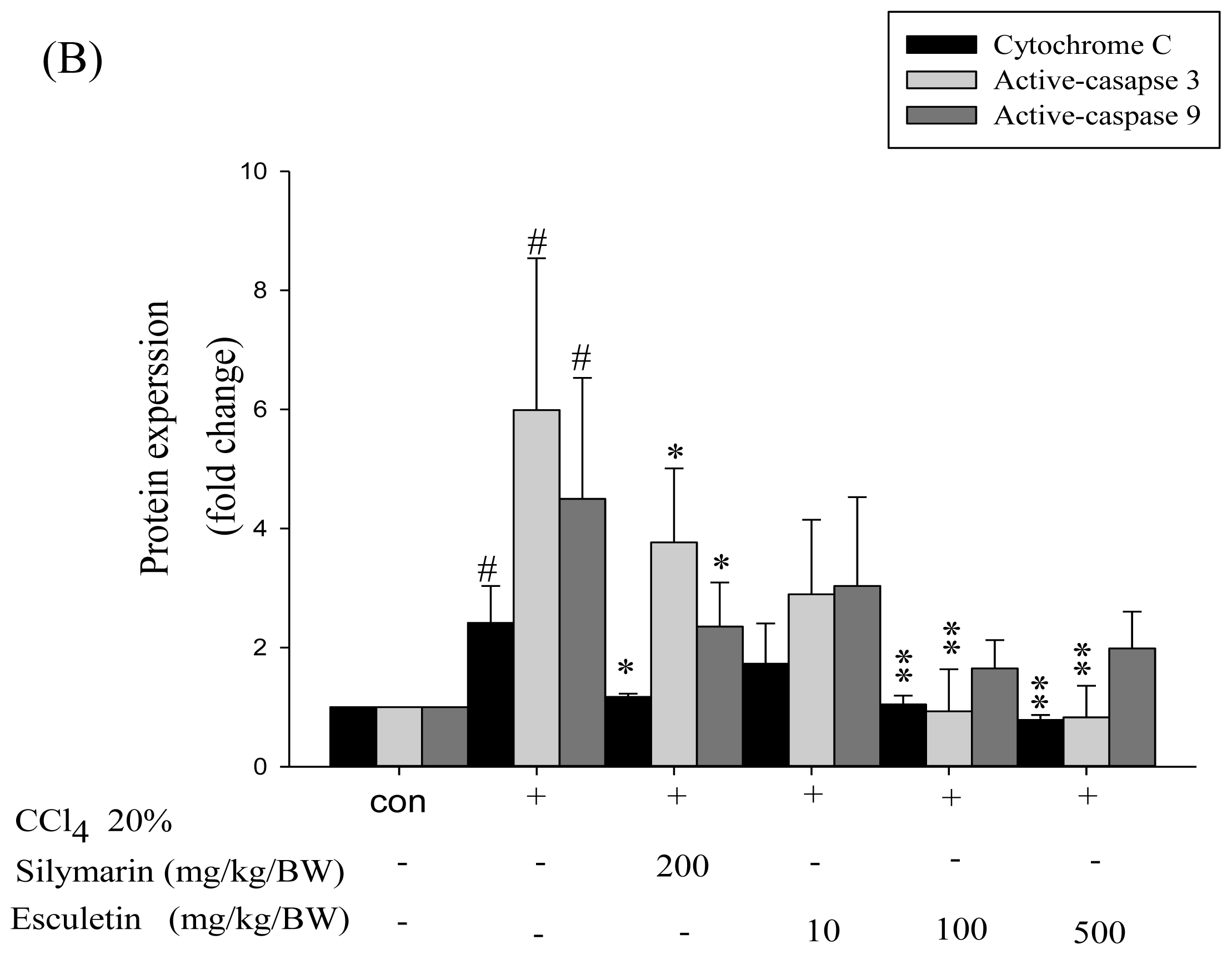
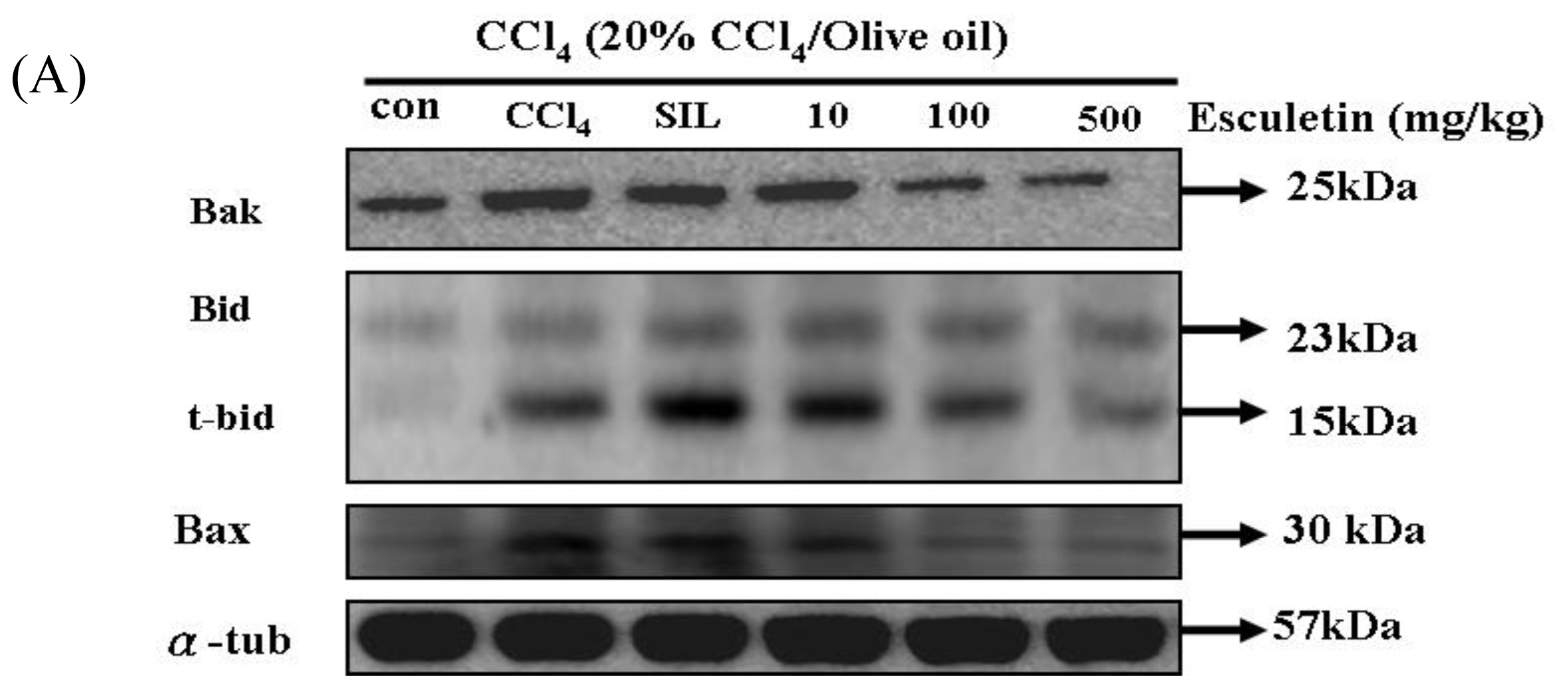
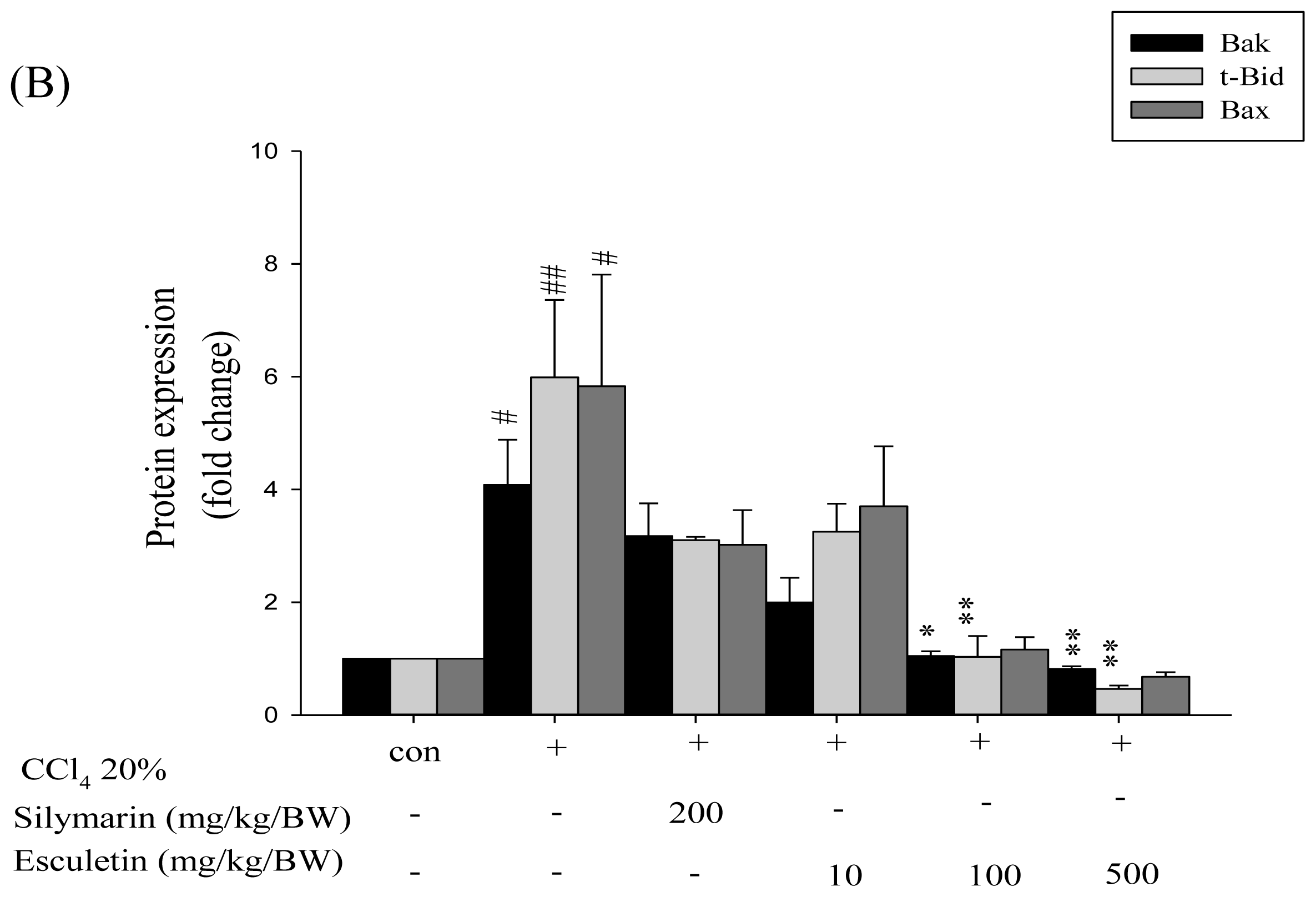
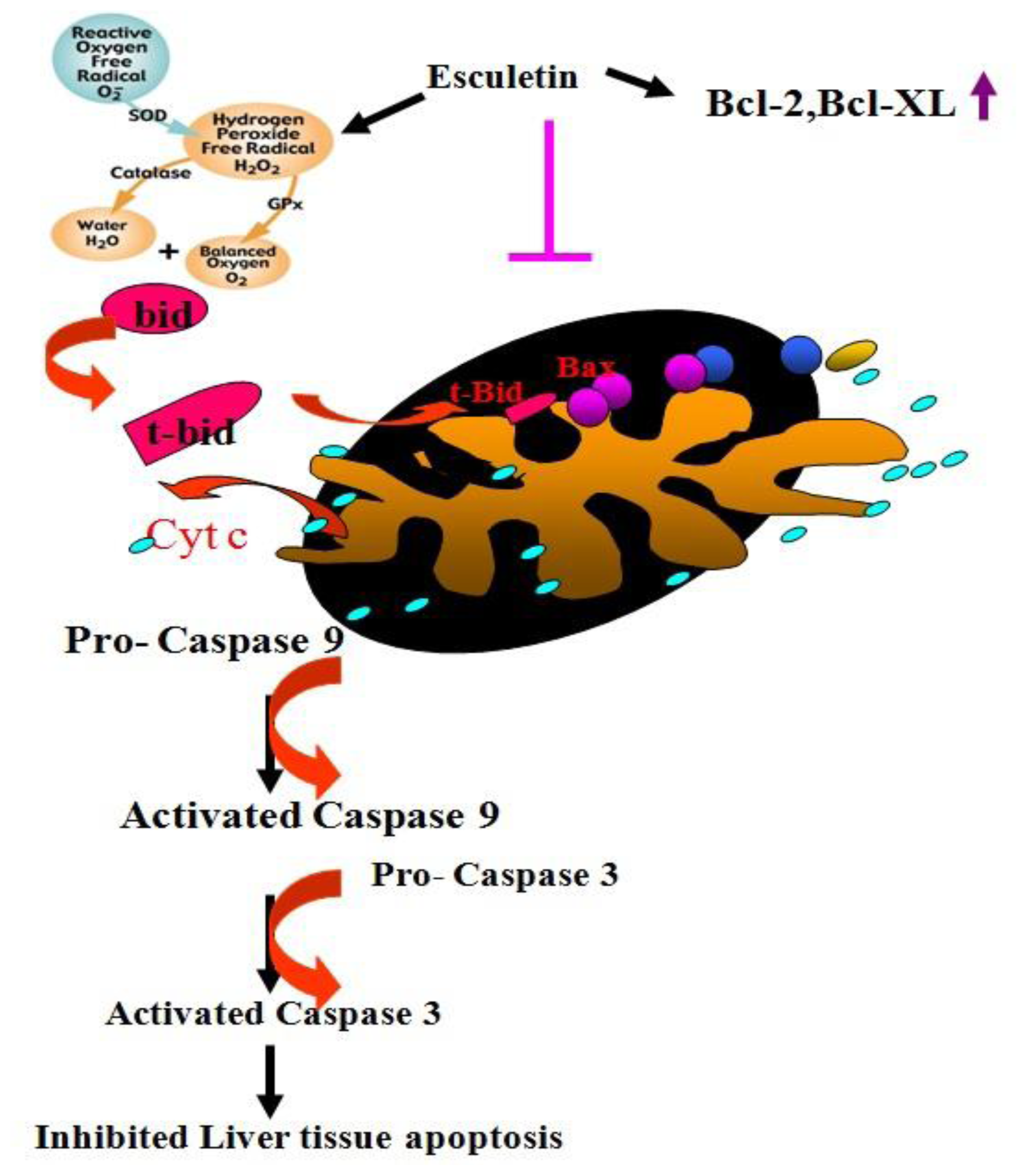
2.4. Changes in Caspase-9, Caspase-3 and Cytochrome c Protein Levels in CCl4 -Treated Rats
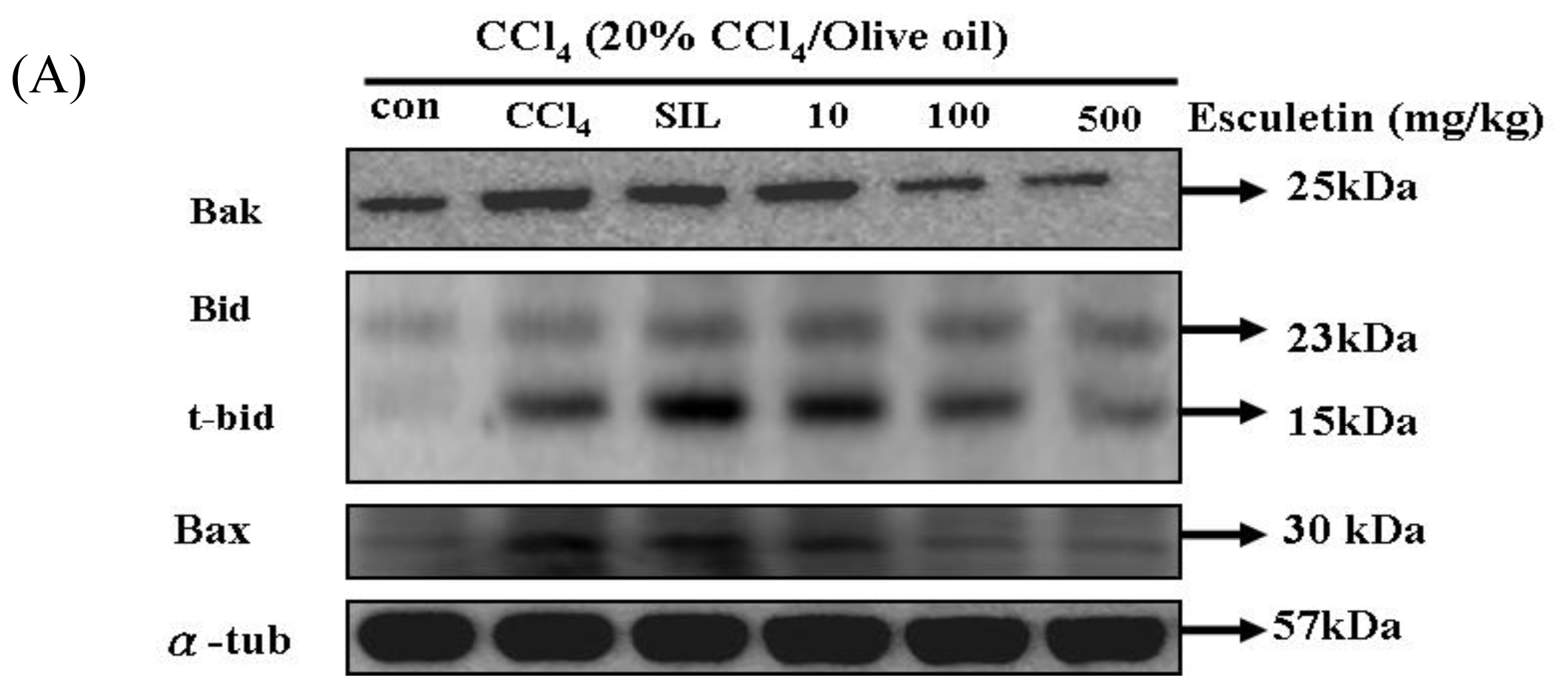
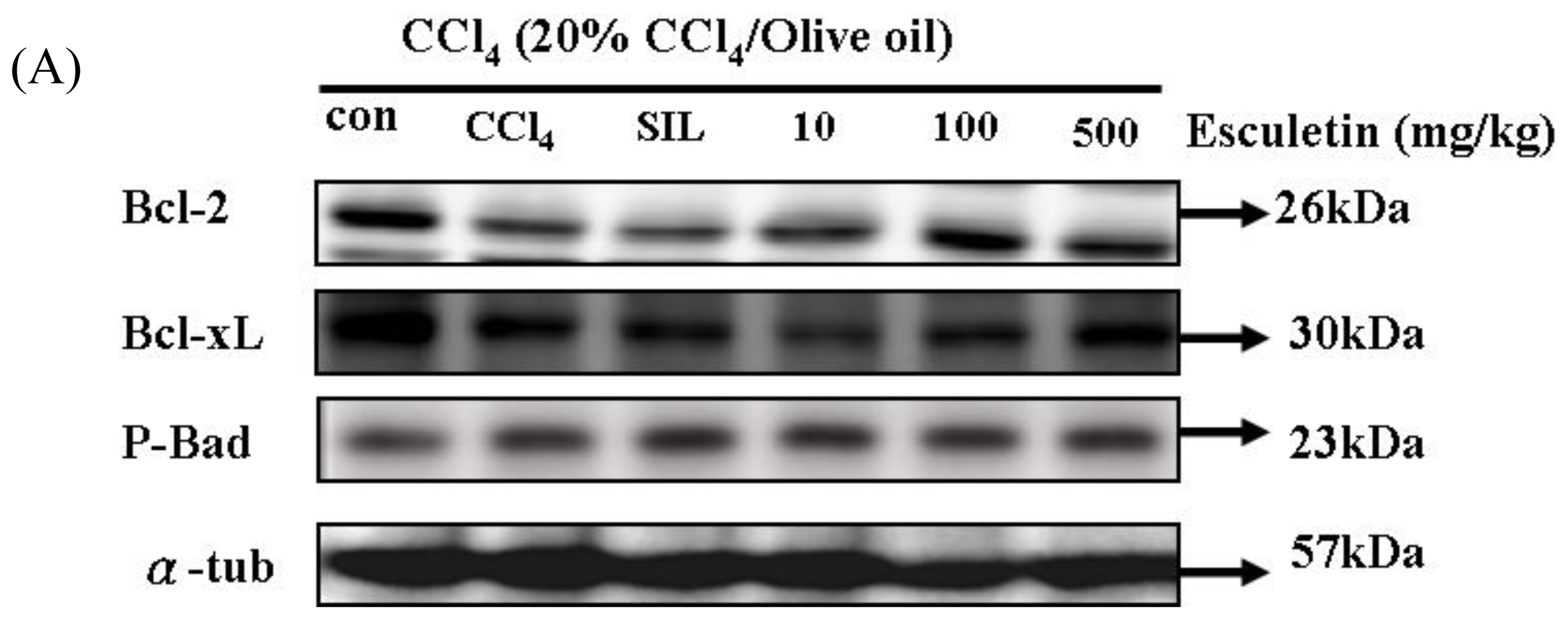
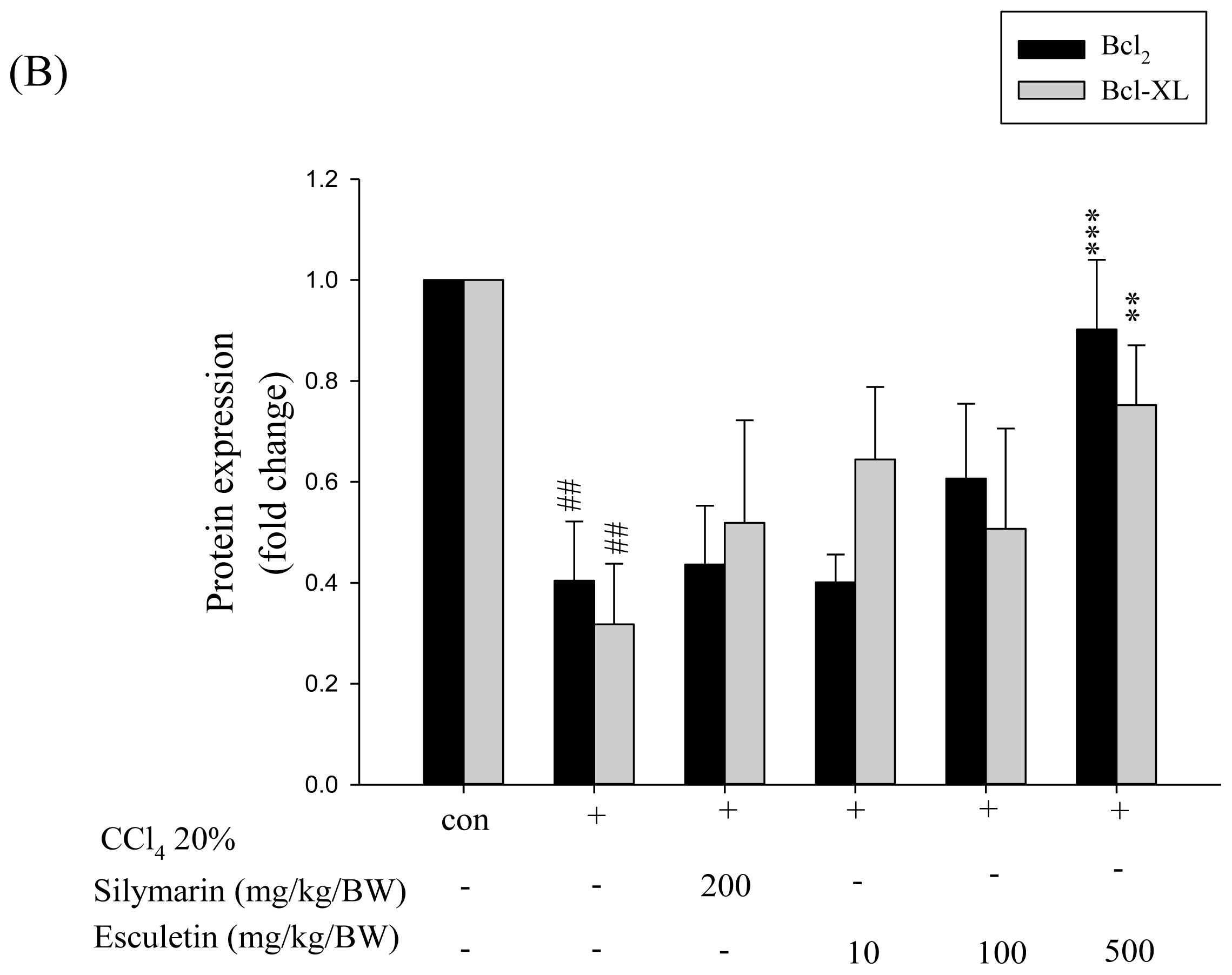
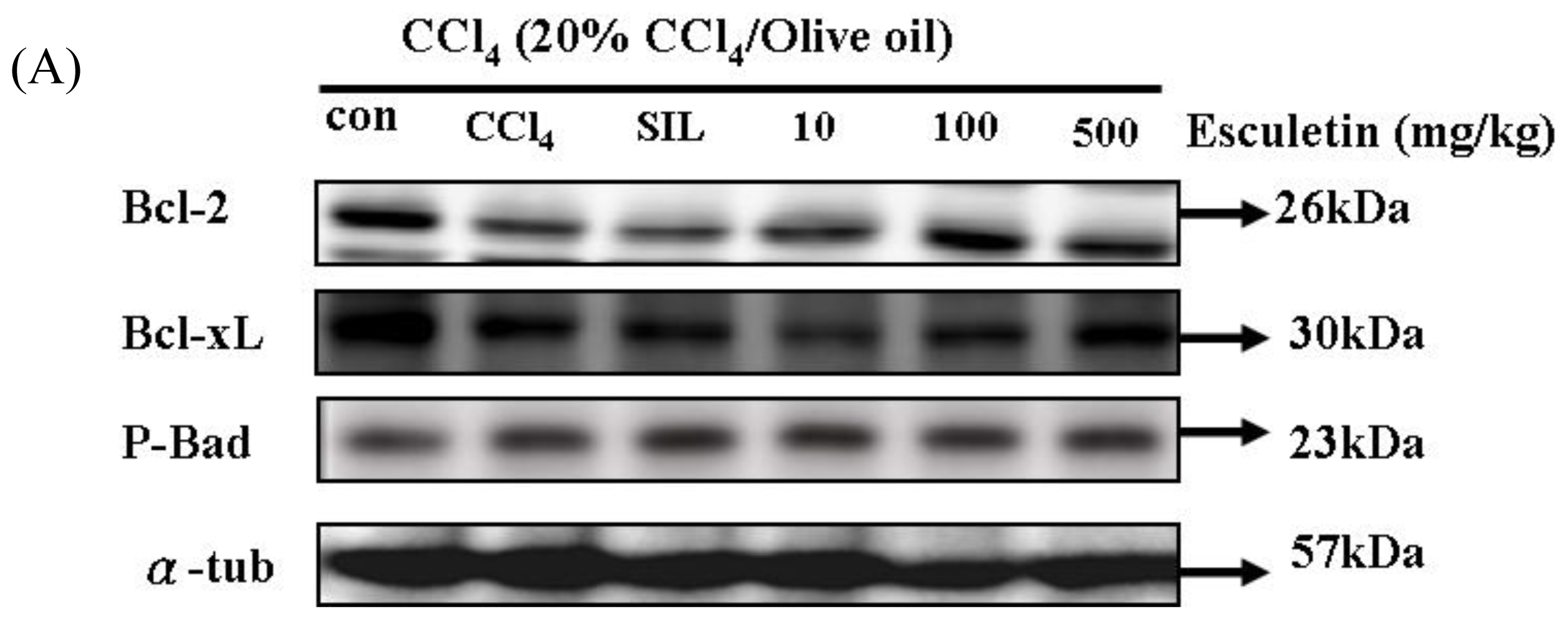
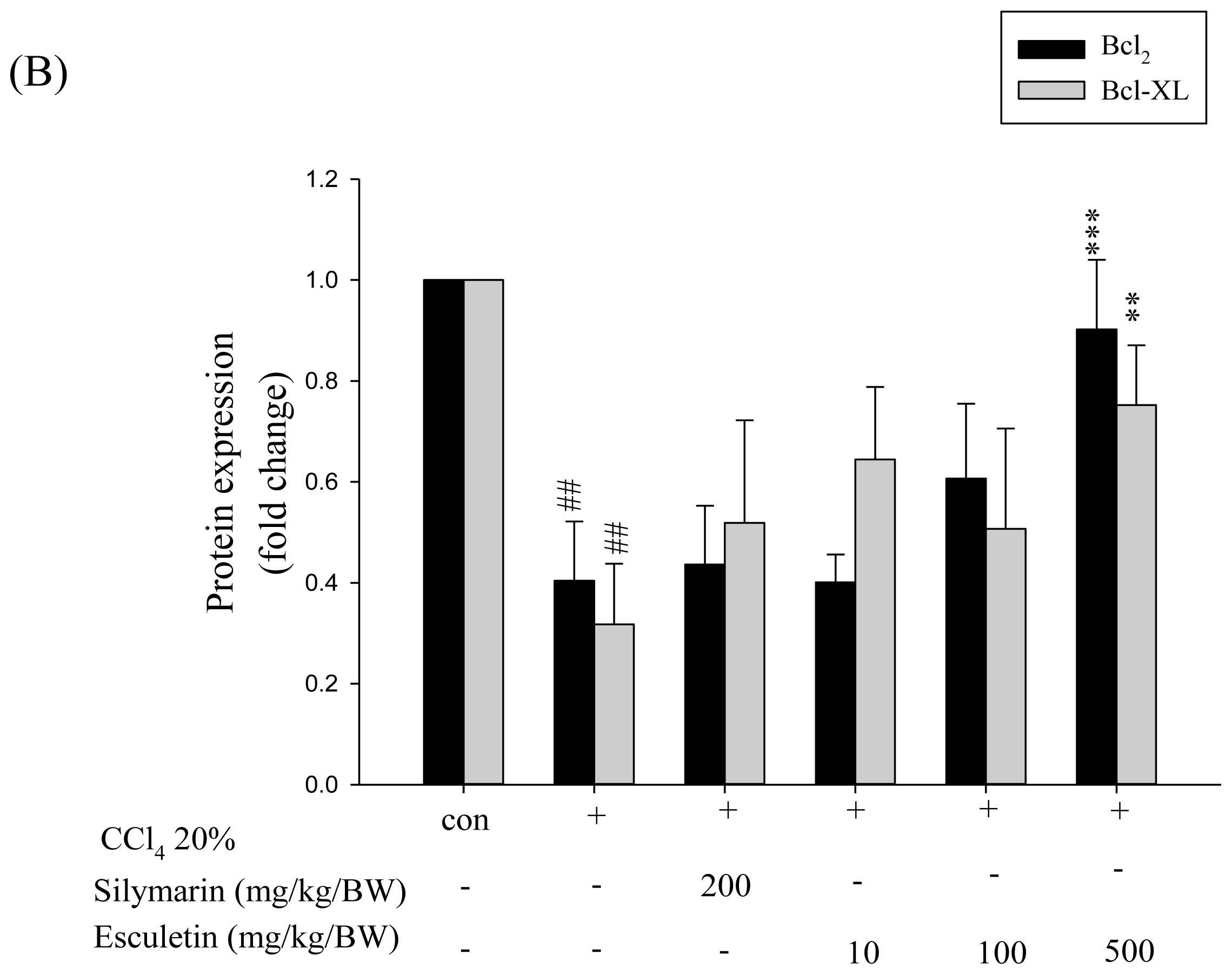
2.5. Changes in Bcl-2 Family Components in CCl4-Treated Rats
3. Experimental Section
3.1. Animals
3.2. CCl4-Induced Liver Apoptosis
3.3. Assessment of Liver Functions
3.4. Antioxidant Enzymes Activity Measurements
3.5. Pathological Examinations
3.6. Assessment of Liver Apoptosis Proteins and Anti-Apoptic Proteins
3.7. Statistical Analysis
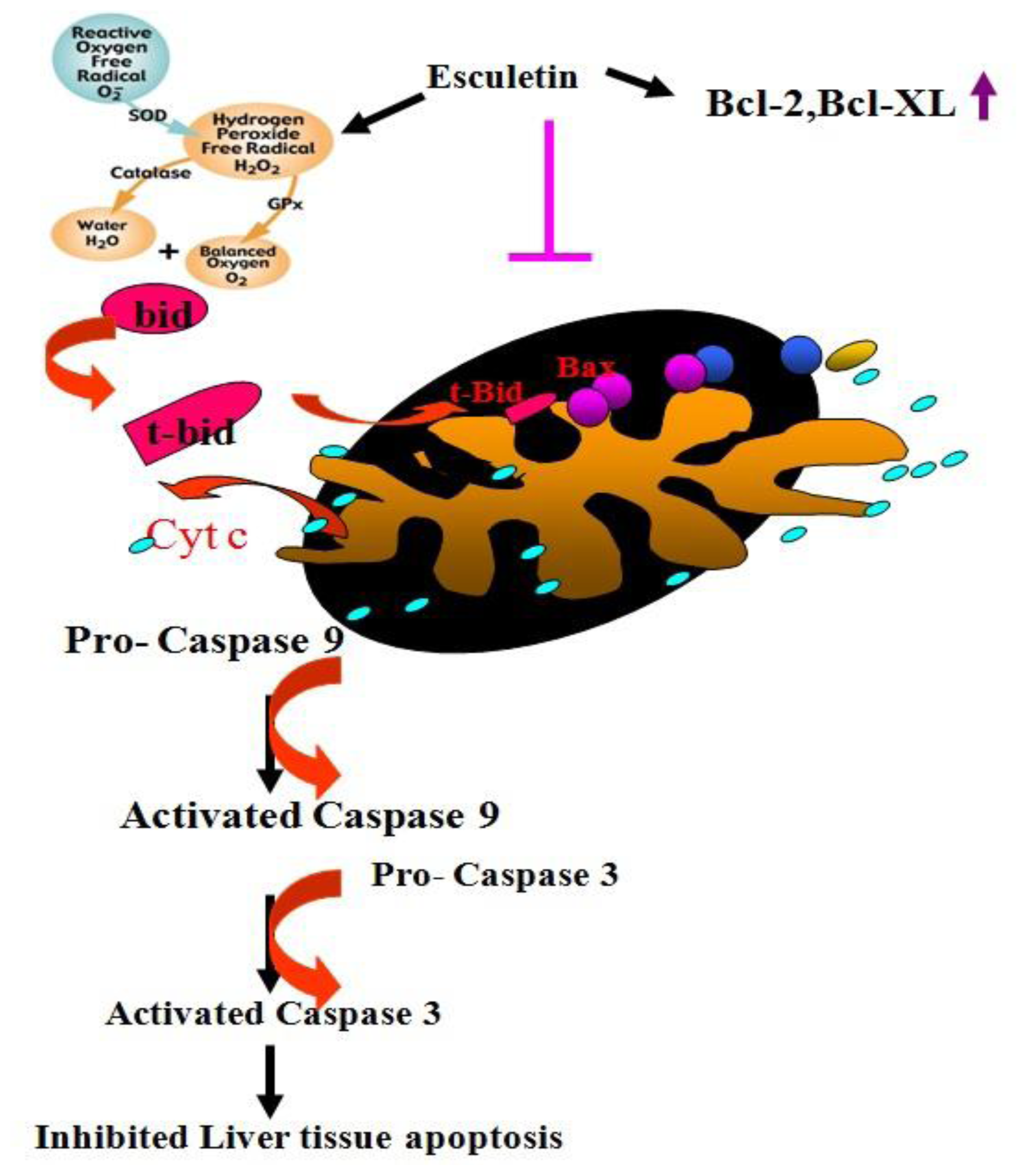
4. Conclusions
Acknowledgments
References
- Wu, HC; Wang, Q; Wang, LW; Yang, HI; Ahsan, H; Tsai, WY; Wang, LY; Chen, SY; Chen, CJ; Santella, RM. Urinary 8-oxodeoxyguanosine, aflatoxin B1 exposure and hepatitis B virus infection and hepatocellular carcinoma in Taiwan. Carcinogenesis 2007, 28, 995–999. [Google Scholar]
- McCaughan, GW. Asian perspectives on viral hepatitis: Hepatitis C virus infection. J. Gastroenterol. Hepatol 2000, 15, G90–G93. [Google Scholar]
- Fylaktakidou, KC; Hadjipavlou-Litina, DJ; Litinas, KE; Nicolaides, DN. Natural and synthetic coumarin derivatives with anti-inflammatory/antioxidant activities. Curr. Pharm. Des 2004, 10, 3813–3833. [Google Scholar]
- Kim, SH; Kang, KA; Zhang, R; Piao, MJ; Ko, DO; Wang, ZH; Chae, SW; Kang, SS; Lee, KH; Kang, HK; et al. Protective effect of esculetin against oxidative stress-induced cell damage via scavenging reactive oxygen species. Acta Pharmacol. Sin 2008, 29, 1319–1326. [Google Scholar]
- Lee, BC; Lee, SY; Lee, HJ; Sim, GS; Kim, JH; Cho, YH; Lee, DH; Pyo, HB; Choe, TB; Moon, DC; et al. Anti-oxidative and photo-protective effects of coumarins isolated from Fraxinus chinensis. Arch. Pharm. Res 2007, 30, 1293–1301. [Google Scholar]
- Martin-Aragon, S; Benedi, JM; Villar, AM. Effects of the antioxidant (6,7-dihydroxycoumarin) esculetin on the glutathione system and lipid peroxidation in mice. Gerontology 1998, 44, 21–25. [Google Scholar]
- Tubaro, A; Del Negro, P; Ragazzi, E; Zampiron, S; Loggia Della, R. Anti-inflammatory and peripheral analgesic activity of esculetinin vivo. Pharmacol Res Commun 1988, 20(Suppl 5), 83–85. [Google Scholar]
- Huang, HC; Lai, MW; Wang, HR; Chung, YL; Hsieh, LM; Chen, CC. Antiproliferative effect of esculetin on vascular smooth muscle cells: Possible roles of signal transduction pathways. Eur. J. Pharmacol 1993, 237, 39–44. [Google Scholar]
- Lin, WL; Wang, CJ; Tsai, YY; Liu, CL; Hwang, JM; Tseng, TH. Inhibitory effect of esculetin on oxidative damage induced by t-butyl hydroperoxide in rat liver. Arch. Toxicol 2000, 74, 467–472. [Google Scholar]
- Von Minden, HM; Brandenburg, K; Seydel, U; Koch, MH; Garamus, V; Willumeit, R; Vill, V. Thermotropic and lyotropic properties of long chain alkyl glycopyranosides. Part II. Disaccharide headgroups. Chem. Phys. Lipids 2000, 106, 157–179. [Google Scholar]
- James, JL; Moody, DE; Chan, CH; Smuckler, EA. The phospholipids of the hepatic endoplasmic reticulum. Structural change in liver injury. Biochem. J 1982, 206, 203–210. [Google Scholar]
- Gines, P; Cardenas, A; Arroyo, V; Rodes, J. Management of cirrhosis and ascites. N. Engl. J. Med 2004, 350, 1646–1654. [Google Scholar]
- Iredale, JP. Cirrhosis: New research provides a basis for rational and targeted treatments. Br. Med. J. Int. Ed 2003, 327, 143–147. [Google Scholar]
- Kim, SH; Cheon, HJ; Yun, N; Oh, ST; Shin, E; Shim, KS; Lee, SM. Protective effect of a mixture of Aloe vera and Silybum marianum against carbon tetrachloride-induced acute hepatotoxicity and liver fibrosis. J. Pharmacol. Sci 2009, 109, 119–127. [Google Scholar]
- Lee, CH; Park, SW; Kim, YS; Kang, SS; Kim, JA; Lee, SH; Lee, SM. Protective mechanism of glycyrrhizin on acute liver injury induced by carbon tetrachloride in mice. Biol. Pharm. Bull 2007, 30, 1898–1904. [Google Scholar]
- Hjelle, JJ; Klaassen, CD. Glucuronidation and biliary excretion of acetaminophen in rats. J. Pharmacol. Exp. Ther 1984, 228, 407–413. [Google Scholar]
- Ravagnan, L; Roumier, T; Kroemer, G. Mitochondria, the killer organelles and their weapons. J. Cell Physiol 2002, 192, 131–137. [Google Scholar]
- Zhang, J; Dong, M; Li, L; Fan, Y; Pathre, P; Dong, J; Lou, D; Wells, JM; Olivares-Villagomez, D; van Kaer, L; et al. Endonuclease G is required for early embryogenesis and normal apoptosis in mice. Proc. Natl. Acad. Sci. USA 2003, 100, 15782–15787. [Google Scholar]
- Lorenzo, HK; Susin, SA. Mitochondrial effectors in caspase-independent cell death. FEBS Lett 2004, 557, 14–20. [Google Scholar]
- Susin, SA; Zamzami, N; Kroemer, G. Mitochondria as regulators of apoptosis: Doubt no more. Biochim. Biophys. Acta 1998, 1366, 151–165. [Google Scholar]
- Li, P; Nijhawan, D; Budihardjo, I; Srinivasula, SM; Ahmad, M; Alnemri, ES; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar]
- Chipman, D; Barak, Z; Schloss, JV. Biosynthesis of 2-aceto-2-hydroxy acids: Acetolactate synthases and acetohydroxyacid synthases. Biochim. Biophys. Acta 1998, 1385, 401–419. [Google Scholar]
- Ko, JH; Lim, KT. Glycoprotein isolated from Ulmus davidiana NAKAI protects against carbon tetrachloride-induced liver injury in the mouse. J. Pharmacol. Sci 2006, 101, 205–213. [Google Scholar]
- Yamada, Y; Fausto, N. Deficient liver regeneration after carbon tetrachloride injury in mice lacking type 1 but not type 2 tumor necrosis factor receptor. Am. J. Pathol 1998, 152, 1577–1589. [Google Scholar]
- Knodell, RG; Ishak, KG; Black, WC; Chen, TS; Craig, R; Kaplowitz, N; Kiernan, TW; Wollman, J. Formulation and application of a numerical scoring system for assessing histological activity in asymptomatic chronic active hepatitis. Hepatology 1981, 1, 431–435. [Google Scholar]
- Margaill, I; Plotkine, M; Lerouet, D. Antioxidant strategies in the treatment of stroke. Free Radic. Biol. Med 2005, 39, 429–443. [Google Scholar]
- Park, SW; Lee, CH; Kim, YS; Kang, SS; Jeon, SJ; Son, KH; Lee, SM. Protective effect of baicalin against carbon tetrachloride-induced acute hepatic injury in mice. J. Pharmacol. Sci 2008, 106, 136–143. [Google Scholar]
- Poon, MK; Chiu, PY; Mak, DH; Ko, KM. Metformin protects against carbon tetrachloride hepatotoxicity in mice. J. Pharmacol. Sci 2003, 93, 501–504. [Google Scholar]
- Reiter, RJ; Tan, DX; Osuna, C; Gitto, E. Actions of melatonin in the reduction of oxidative stress. A review. J. Biomed. Sci 2000, 7, 444–458. [Google Scholar]
- Kumaravelu, P; Dakshinamoorthy, DP; Subramaniam, S; Devaraj, H; Devaraj, NS. Effect of eugenol on drug-metabolizing enzymes of carbon tetrachloride-intoxicated rat liver. Biochem. Pharmacol 1995, 49, 1703–1707. [Google Scholar]
- Araragi, S; Kondoh, M; Kawase, M; Saito, S; Higashimoto, M; Sato, M. Mercuric chloride induces apoptosis via a mitochondrial-dependent pathway in human leukemia cells. Toxicology 2003, 184, 1–9. [Google Scholar]
- Sun, F; Hamagawa, E; Tsutsui, C; Ono, Y; Ogiri, Y; Kojo, S. Evaluation of oxidative stress during apoptosis and necrosis caused by carbon tetrachloride in rat liver. Biochim. Biophys. Acta 2001, 1535, 186–191. [Google Scholar]
- Brown, GC; Borutaite, V. Nitric oxide, cytochrome c and mitochondria. Biochem. Soc. Symp 1999, 66, 17–25. [Google Scholar]
- Lee, TY; Chang, HH; Wang, GJ; Chiu, JH; Yang, YY; Lin, HC. Water-soluble extract of Salvia miltiorrhiza ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. J. Pharm. Pharmacol 2006, 58, 659–665. [Google Scholar]
- Ding, WX; Nam, OC. Role of oxidative stress and mitochondrial changes in cyanobacteria-induced apoptosis and hepatotoxicity. FEMS Microbiol. Lett 2003, 220, 1–7. [Google Scholar]
- Aebi, H. Catalase in vitro. Meth. Enzymol 1984, 105, 121–126. [Google Scholar]
- Misra, HP; Fridovich, I. The generation of superoxide radical during the autoxidation of hemoglobin. J. Biol. Chem 1972, 247, 6960–6962. [Google Scholar]
- Flohe, L; Gunzler, WA. Assays of glutathione peroxidase. Meth. Enzymol 1984, 105, 114–121. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Activity (U/mg protein) | ||
|---|---|---|---|
| Catalase | SOD | GPx | |
| Control | 55.82 ± 0.3184 | 47.26 ± 0.77 | 64.12 ± 0.72 |
| CCl4 | 28.9 ± 0.484### | 24.84 ± 0.36### | 24.94 ± 0.46### |
| CCl4 + Silymarin200 mg/kg | 33.74 ± 0.47*** | 31.38 ± 0.71*** | 30.42 ± 0.81*** |
| CCl4 + Esculetin 10 mg/kg | 37.76 ± 0.79* | 27.4 ± 0.46 | 28.1 ± 0.55 |
| Cl4 + Esculetin 100 mg/g | 38.13 ± 1.65* | 32.62 ± 0.724*** | 36.06 ± 0.73*** |
| CCl4 + Esculetin 500 mg/kg | 41.16 ± 0.64*** | 32.8 ± 0.55*** | 40.18 ± 0.44*** |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tien, Y.-C.; Liao, J.-C.; Chiu, C.-S.; Huang, T.-H.; Huang, C.-Y.; Chang, W.-T.; Peng, W.-H. Esculetin Ameliorates Carbon Tetrachloride-Mediated Hepatic Apoptosis in Rats. Int. J. Mol. Sci. 2011, 12, 4053-4067. https://doi.org/10.3390/ijms12064053
Tien Y-C, Liao J-C, Chiu C-S, Huang T-H, Huang C-Y, Chang W-T, Peng W-H. Esculetin Ameliorates Carbon Tetrachloride-Mediated Hepatic Apoptosis in Rats. International Journal of Molecular Sciences. 2011; 12(6):4053-4067. https://doi.org/10.3390/ijms12064053
Chicago/Turabian StyleTien, Yun-Chen, Jung-Chun Liao, Chuan-Sung Chiu, Tai-Hung Huang, Chih-Yang Huang, Wen-Te Chang, and Wen-Huang Peng. 2011. "Esculetin Ameliorates Carbon Tetrachloride-Mediated Hepatic Apoptosis in Rats" International Journal of Molecular Sciences 12, no. 6: 4053-4067. https://doi.org/10.3390/ijms12064053