Interferon-β1b Increases Th2 Response in Neuromyelitis Optica

Abstract
:1. Introduction
2. Results
2.1. Comparison of Th1/Th2-Related Chemokine Receptor Expression at Baseline
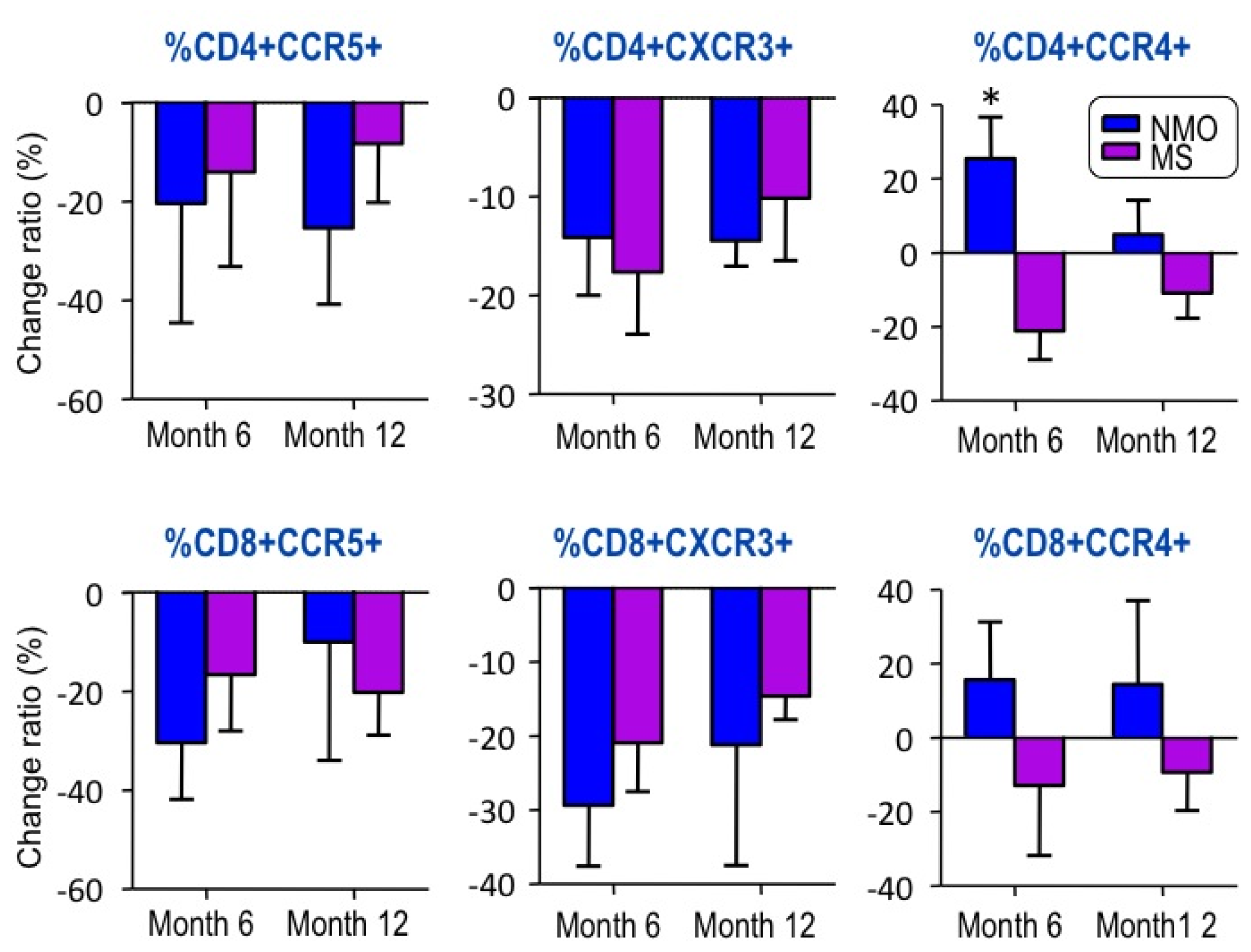
2.2. Changes in Chemokine Receptor Expression by IFN-â1b Treatment
3. Discussion
4. Experimental Section
4.1. Subjects
4.2. Flow Cytometry
4.3. Statistical Analysis
5. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Sørensen, T.L.; Sellebjerg, F. Selective suppression of chemokine receptor CXCR3 expression by interferon-beta1a in multiple sclerosis. Mult. Scler 2002, 8, 104–107. [Google Scholar]
- Sega, S.; Wraber, B.; Mesec, A.; Horvat, A.; Ihan, A. IFN-beta1a and IFN-beta1b have different patterns of influence on cytokines. Clin. Neurol. Neurosurg 2004, 106, 255–258. [Google Scholar]
- Ersoy, E.; Kuş, C.N.; Sener, U.; Coker, I.; Zorlu, Y. The effects of interferon-beta on interleukin-10 in multiple sclerosis patients. Eur. J. Neurol 2005, 12, 208–211. [Google Scholar]
- Krakauer, M.; Sorensen, P.S.; Khademi, M.; Olsson, T.; Sellebjerg, F. Dynamic T-lymphocyte chemokine receptor expression induced by interferon-beta therapy in multiple sclerosis. Scand. J. Immunol 2006, 64, 155–163. [Google Scholar]
- Shimizu, Y.; Ota, K.; Kubo, S.; Kabasawa, C.; Kobayashi, M.; Ohashi, T.; Uchiyama, S. Association of Th1/Th2-related chemokine receptors in peripheral T cells with disease activity in patients with multiple sclerosis and neuromyelitis optica. Eur. Neurol 2011, 66, 91–97. [Google Scholar]
- Wingerchuk, D.M.; Lennon, V.A.; Lucchinetti, C.F.; Pittock, S.J.; Weinshenker, B.G. The spectrum of neuromyelitis optica. Lancet Neurol 2007, 6, 805–815. [Google Scholar]
- Lucchinetti, C.F.; Mandler, R.N.; McGavern, D.; Bruck, W.; Gleich, G.; Ransohoff, R.M.; Trebst, C.; Weinshenker, B.; Wingerchuk, D.; Parisi, J.E.; et al. A role for humoral mechanisms in the pathogenesis of Devic’s neuromyelitis optica. Brain 2002, 125, 1450–1461. [Google Scholar]
- Hosokawa, T.; Nakajima, H.; Doi, Y.; Sugino, M.; Kimura, F.; Hanafusa, T.; Takahashi, T. Increased serum matrix metalloproteinase-9 in neuromyelitis optica: Implication of disruption of blood-brain barrier. J. Neuroimmunol 2011, 236, 81–86. [Google Scholar]
- Nakajima, H.; Hosokawa, T.; Sugino, M.; Kimura, F.; Sugasawa, J.; Hanafusa, T.; Takahashi, T. Visual field defects of optic neuritis in neuromyelitis optica compared with multiple sclerosis. BMC Neurol 2010, 10, 45. [Google Scholar]
- Lennon, V.A.; Wingerchuk, D.M.; Kryzer, T.J.; Pittock, S.J.; Lucchinetti, C.F.; Fujihara, K.; Nakashima, I.; Weinshenker, B.G. A serum autoantibody marker of neuromyelitis optica: Distinction from multiple sclerosis. Lancet 2004, 364, 2106–2112. [Google Scholar]
- Jarius, S.; Aboul-Enein, F.; Waters, P.; Kuenz, B.; Hauser, A.; Berger, T.; Lang, W.; Reindl, M.; Vincent, A.; Kristoferitsch, W. Antibody to aquaporin-4 in the long-term course of neuromyelitis optica. Brain 2008, 131, 3072–3080. [Google Scholar]
- Saida, T.; Tashiro, K.; Itoyama, Y.; Sato, T.; Ohashi, Y.; Zhao, Z. Interferon Beta-1b Multiple Sclerosis Study Group of Japan. Interferon beta-1b is effective in Japanese RRMS patients: A randomized multicenter study. Neurology 2005, 64, 621–630. [Google Scholar]
- Tanaka, M.; Tanaka, K.; Komori, M. Interferon-beta(1b) treatment in neuromyelitis optica. Eur. Neurol 2009, 62, 167–170. [Google Scholar]
- Uzawa, A.; Mori, M.; Hayakawa, S.; Masuda, S.; Kuwabara, S. Different responses to interferon beta-1b treatment in patients with neuromyelitis optica and multiple sclerosis. Eur. J. Neurol 2010, 17, 672–676. [Google Scholar]
- Simpson, J.E.; Newcombe, J.; Cuzner, M.L.; Woodroofe, M.N. Expression of monocyte chemoattractant protein-1 and other beta-chemokines by resident glia and inflammatory cells in multiple sclerosis lesions. J. Neuroimmunol 1998, 84, 238–249. [Google Scholar]
- Sorensen, T.L.; Tani, M.; Jensen, J.; Pierce, V.; Lucchinetti, C.; Folcik, V.A.; Qin, S.; Rottman, J.; Sellebjerg, F.; Strieter, R.M.; et al. Expression of specific chemokines and chemokine receptors in the central nervous system of multiple sclerosis patients. J. Clin. Invest 1999, 103, 807–815. [Google Scholar]
- Balashov, K.E.; Rottman, J.B.; Weiner, H.L.; Hancock, W.W. CCR5(+) and CXCR3(+) T cells are increased in multiple sclerosis and their ligands MIP-1alpha and IP-10 are expressed in demyelinating brain lesions. Proc. Natl. Acad. Sci. USA 1999, 96, 6873–6878. [Google Scholar]
- Nakajima, H.; Fukuda, K.; Doi, Y.; Sugino, M.; Kimura, F.; Hanafusa, T.; Ikemoto, T.; Shimizu, A. Expression of TH1/TH2-related chemokine receptors on peripheral T cells and correlation with clinical disease activity in patients with multiple sclerosis. Eur. Neurol 2004, 52, 162–168. [Google Scholar]
- Uzawa, A.; Mori, M.; Hayakawa, S.; Masuda, S.; Nomura, F.; Kuwabara, S. Expression of chemokine receptors on peripheral blood lymphocytes in multiple sclerosis and neuromyelitis optica. BMC Neurol 2010, 10, 113. [Google Scholar]
- Warabi, Y.; Matsumoto, Y.; Hayashi, H. Interferon beta-1b exacerbates multiple sclerosis with severe optic nerve and spinal cord demyelination. J. Neurol. Sci 2007, 252, 57–61. [Google Scholar]
- Shimizu, Y.; Yokoyama, K.; Misu, T.; Takahashi, T.; Fujihara, K.; Kikuchi, S.; Itoyama, Y.; Iwata, M. Development of extensive brain lesions following interferon beta therapy in relapsing neuromyelitis optica and longitudinally extensive myelitis. J. Neurol 2008, 255, 305–307. [Google Scholar]
- Shimizu, J.; Hatanaka, Y.; Hasegawa, M.; Iwata, A.; Sugimoto, I.; Date, H.; Goto, J.; Shimizu, T.; Takatsu, M.; Sakurai, Y.; et al. IFNβ-1b may severely exacerbate Japanese optic-spinal MS in neuromyelitis optica spectrum. Neurology 2010, 75, 1423–1427. [Google Scholar]
- Takahashi, T.; Fujihara, K.; Nakashima, I.; Misu, T.; Miyazawa, I.; Nakamura, M.; Watanabe, S.; Shiga, Y.; Kanaoka, C.; Fujimori, J.; et al. Anti-aquaporin-4 antibody is involved in the pathogenesis of NMO: A study on antibody titre. Brain 2007, 130, 1235–1243. [Google Scholar]
- Misu, T.; Fujihara, K.; Kakita, A.; Konno, H.; Nakamura, M.; Watanabe, S.; Takahashi, T.; Nakashima, I.; Takahashi, H.; Itoyama, Y. Loss of aquaporin 4 in lesions of neuromyelitis optica: Distinction from multiple sclerosis. Brain 2007, 130, 1224–1234. [Google Scholar]
- Rudick, R.A.; Goelz, S.E. Beta-interferon for multiple sclerosis. Exp Cell Res 2011, 317, 1301–1311. [Google Scholar]
- Kieseier, B.C. The mechanism of action of interferon-â in relapsing multiple sclerosis. CNS Drugs 2011, 25, 491–502. [Google Scholar]
- Wandinger, K.P.; Stürzebecher, C.S.; Bielekova, B.; Detore, G.; Rosenwald, A.; Staudt, L.M.; McFarland, H.F.; Martin, R. Complex immunomodulatory effects of interferon-beta in multiple sclerosis include the upregulation of T helper 1-associated marker genes. Ann. Neurol 2001, 50, 349–357. [Google Scholar]
- Krumbholz, M.; Faber, H.; Steinmeyer, F.; Hoffmann, L.A.; Kümpfel, T.; Pellkofer, H.; Derfuss, T.; Ionescu, C.; Starck, M.; Hafner, C.; et al. Interferon-beta increases BAFF levels in multiple sclerosis: implications for B cell autoimmunity. Brain 2008, 131, 1455–1463. [Google Scholar]
- Jonsson, M.V.; Szodoray, P.; Jellestad, S.; Jonsson, R.; Skarstein, K. Association between circulating levels of the novel TNF family members APRIL and BAFF and lymphoid organization in primary Sjögren’s syndrome. J. Clin. Immunol 2005, 25, 189–201. [Google Scholar]
- Stohl, W.; Metyas, S.; Tan, S.M.; Cheema, G.S.; Oamar, B.; Xu, D.; Roschke, V.; Wu, Y.; Baker, K.P.; Hilbert, D.M. B lymphocyte stimulator overexpression in patients with systemic lupus erythematosus: Longitudinal observations. Arthritis. Rheum 2003, 48, 3475–3486. [Google Scholar]
- Krumbholz, M.; Specks, U.; Wick, M.; Kalled, S.L.; Jenne, D.; Meinl, E. BAFF is elevated in serum of patients with Wegener’s granulomatosis. J. Autoimmun 2005, 25, 298–302. [Google Scholar]
- Palace, J.; Leite, M.I.; Nairne, A.; Vincent, A. Interferon Beta treatment in neuromyelitis optica: Increase in relapses and aquaporin 4 antibody titers. Arch. Neurol 2010, 67, 1016–1017. [Google Scholar]
- Karni, A.; Balashov, K.; Hancock, W.W.; Bharanidharan, P.; Abraham, M.; Khoury, S.J.; Weiner, H.L. Cyclophosphamide modulates CD4+ T cells into a T helper type 2 phenotype and reverses increased IFN-gamma production of CD8+ T cells in secondary progressive multiple sclerosis. J. Neuroimmunol 2004, 146, 189–198. [Google Scholar]
- Acosta-Rodriguez, E.V.; Rivino, L.; Geginat, J.; Jarrossay, D.; Gattorno, M.; Lanzavecchia, A.; Sallusto, F.; Napolitani, G. Surface phenotype and antigenic specificity of human interleukin 17-producing T helper memory cells. Nat. Immunol 2007, 8, 639–646. [Google Scholar]
- Lim, H.W.; Lee, J.; Hillsamer, P.; Kim, C.H. Human Th17 cells share major trafficking receptors with both polarized effector T cells and FOXP3+ regulatory T cells. J. Immunol 2008, 180, 122–129. [Google Scholar]
- Axtell, R.C.; Raman, C.; Steinman, L. Interferon-β exacerbates Th17-mediated inflammatory disease. Trends. Immunol 2011, 32, 272–277. [Google Scholar]
- Ishizu, T.; Osoegawa, M.; Mei, F.J.; Kikuchi, H.; Tanaka, M.; Takakura, Y.; Minohara, M.; Murai, H.; Mihara, F.; Taniwaki, T.; et al. Intrathecal activation of the IL-17/IL-8 axis in opticospinal multiple sclerosis. Brain 2005, 128, 988–1002. [Google Scholar]
- Wingerchuk, D.M.; Lennon, V.A.; Pittock, S.J.; Lucchinetti, C.F.; Weinshenker, B.G. Revised diagnostic criteria for neuromyelitis optica. Neurology 2006, 66, 1485–1489. [Google Scholar]
- McDonald, W.I.; Compston, A.; Edan, G.; Goodkin, D.; Hartung, H.P.; Lublin, F.D.; McFarland, H.F.; Paty, D.W.; Polman, C.H.; Reingold, S.C.; et al. Recommended diagnostic criteria for multiple sclerosis: Guidelines from the International Panel on the diagnosis of multiple sclerosis. Ann. Neurol 2001, 50, 121–127. [Google Scholar]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar]
- Nakajima, H.; Sugino, M.; Kimura, F.; Hanafusa, T.; Ikemoto, T.; Shimizu, A. Increased Intrathecal Chemokine Receptor CCR2 Expression in Multiple Sclerosis. Biomark. Insights 2007, 2, 463–468. [Google Scholar]
- Nakajima, H.; Sugino, M.; Kimura, F.; Hanafusa, T.; Ikemoto, T.; Shimizu, A. Decreased CD14+CCR2+ monocytes in active multiple sclerosis. Neurosci. Lett 2004, 363, 187–189. [Google Scholar]


{kind=link}
{kind=link}
| NMO (n = 10) | MS (n = 16) | HC (n = 14) | |
|---|---|---|---|
| CD4+CCR5+ | 4.71 ± 1.44 | 6.17 ± 2.74 | 5.97 ± 0.99 |
| CD8+CCR5+ | 9.53 ± 3.69 | 16.14 ± 8.33 | 15.40 ± 4.02 * |
| CD4+CXCR3+ | 24.58 ± 3.44 | 18.81 ± 5.05 ** | 15.75 ± 2.52 *** |
| CD8+CXCR3+ | 24.55 ± 8.74 | 26.58 ± 9.52 | 23.0 ± 3.66 |
| CD4+CCR4+ | 13.48 ± 5.15 | 10.94 ± 3.24 | 11.34 ± 1.95 |
| CD8+CCR4+ | 2.44 ± 2.43 | 1.06 ± 0.65 | 0.71 ± 0.37 |
| NMO (n = 5) | MS (n = 6) | p Value | |||
|---|---|---|---|---|---|
| Age | 55.3 | 42.2 | <0.05 | ||
| Disease duration (years) | 9.1 | 6.8 | ns | ||
| EDSS score | 5.1 | 3.3 | ns | ||
| Annualized relapse rate before IFN-â1b | 2 |  ns ns | 2.3 | <0.05 | ns |
| Annualized relapse rate after start of IFN-â1b | 2 | 0.3 | ns |
| NMO (n = 10) | MS (n = 16) | HC (n = 10) | p Value (NMO vs. MS) | |
|---|---|---|---|---|
| Sex (M:F) | 0:10 | 5:11 | 4:6 | ns |
| Age | 45.1 ± 12.6 | 35.9 ± 12.9 | 33.4 ± 7.2 | ns |
| Age at onset (years) | 37.2 ± 11.5 | 32.9 ± 12.8 | ns | |
| Disease duration (years) | 7.5 ± 8.8 | 3.5 ± 4.8 | ns | |
| Number of relapse | 5.6 ± 4.5 | 2.4 ± 1.8 | <0.05 | |
| EDSS score | 6.5 ± 2.1 | 2.9 ± 0.8 | <0.0001 | |
| Treatment | 5: IFN-â1b 5: treated with PSL and/or AZT | 12: IFN-â1b | ||
| Anti-AQP4 antibody | 100% | 0% | <0.0001 | |
| LESCL | 80% | 0% | <0.0001 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nakajima, H.; Hosokawa, T.; Doi, Y.; Ikemoto, T.; Ishida, S.; Kimura, F.; Hanafusa, T. Interferon-β1b Increases Th2 Response in Neuromyelitis Optica. Int. J. Mol. Sci. 2012, 13, 12213-12223. https://doi.org/10.3390/ijms131012213
Nakajima H, Hosokawa T, Doi Y, Ikemoto T, Ishida S, Kimura F, Hanafusa T. Interferon-β1b Increases Th2 Response in Neuromyelitis Optica. International Journal of Molecular Sciences. 2012; 13(10):12213-12223. https://doi.org/10.3390/ijms131012213
Chicago/Turabian StyleNakajima, Hideto, Takafumi Hosokawa, Yoshimitu Doi, Toshiyuki Ikemoto, Shimon Ishida, Fumiharu Kimura, and Toshiaki Hanafusa. 2012. "Interferon-β1b Increases Th2 Response in Neuromyelitis Optica" International Journal of Molecular Sciences 13, no. 10: 12213-12223. https://doi.org/10.3390/ijms131012213