Coenzyme Q Metabolism Is Disturbed in High Fat Diet-Induced Non Alcoholic Fatty Liver Disease in Rats

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
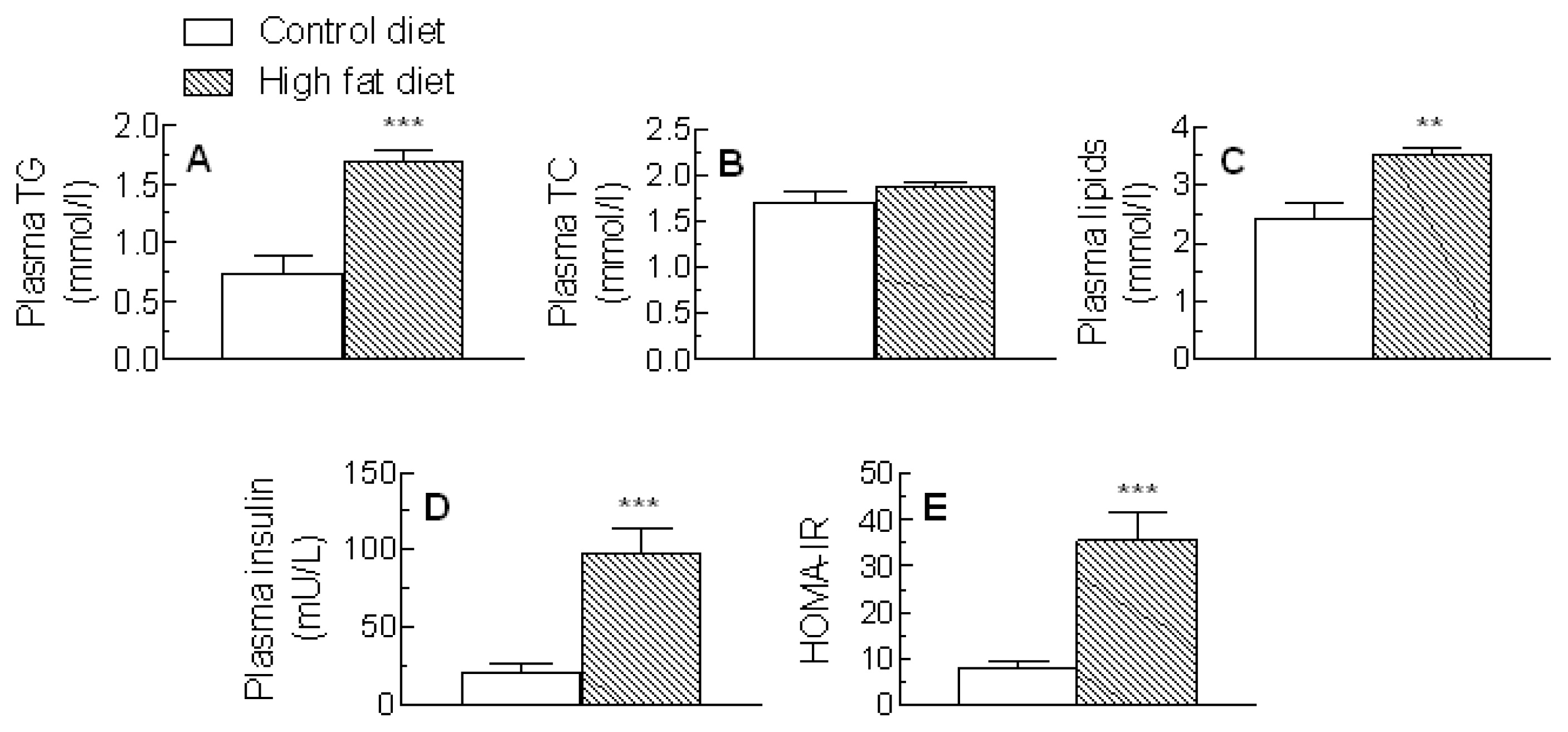
2.1. Plasma Lipids, Enzymes, Insulin and Homeostatic Model Assessment of Insulin Resistance (HOMA-IR)
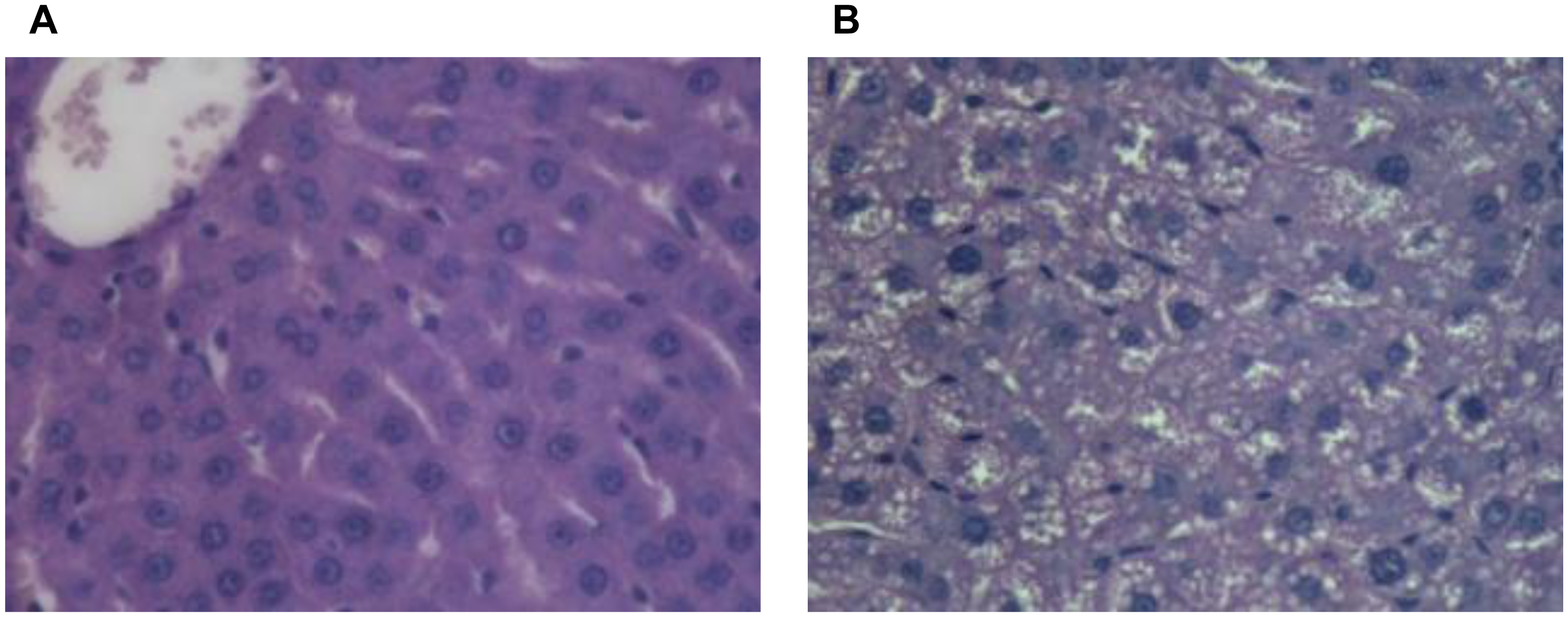
2.2. Liver Lipids and Histology
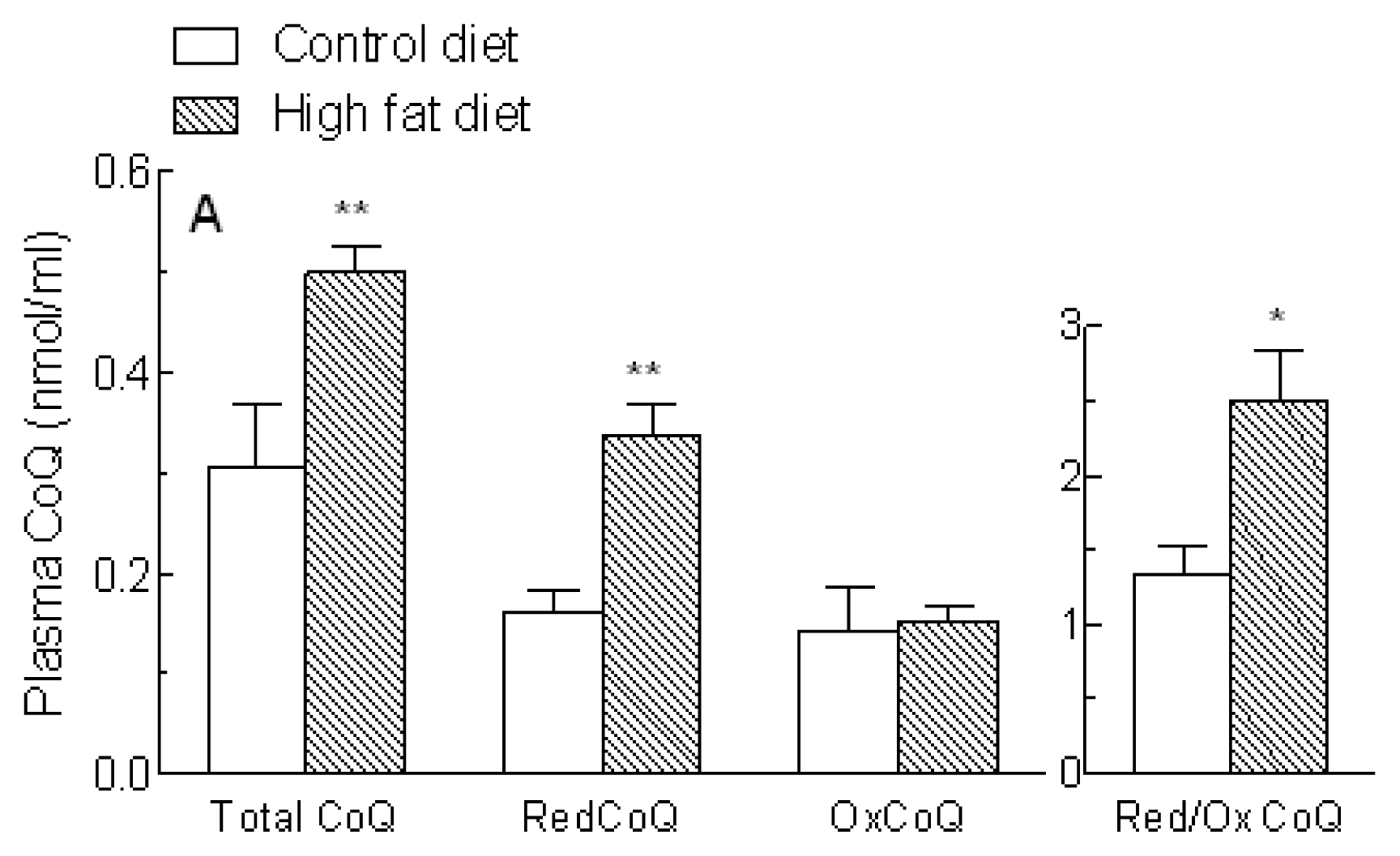
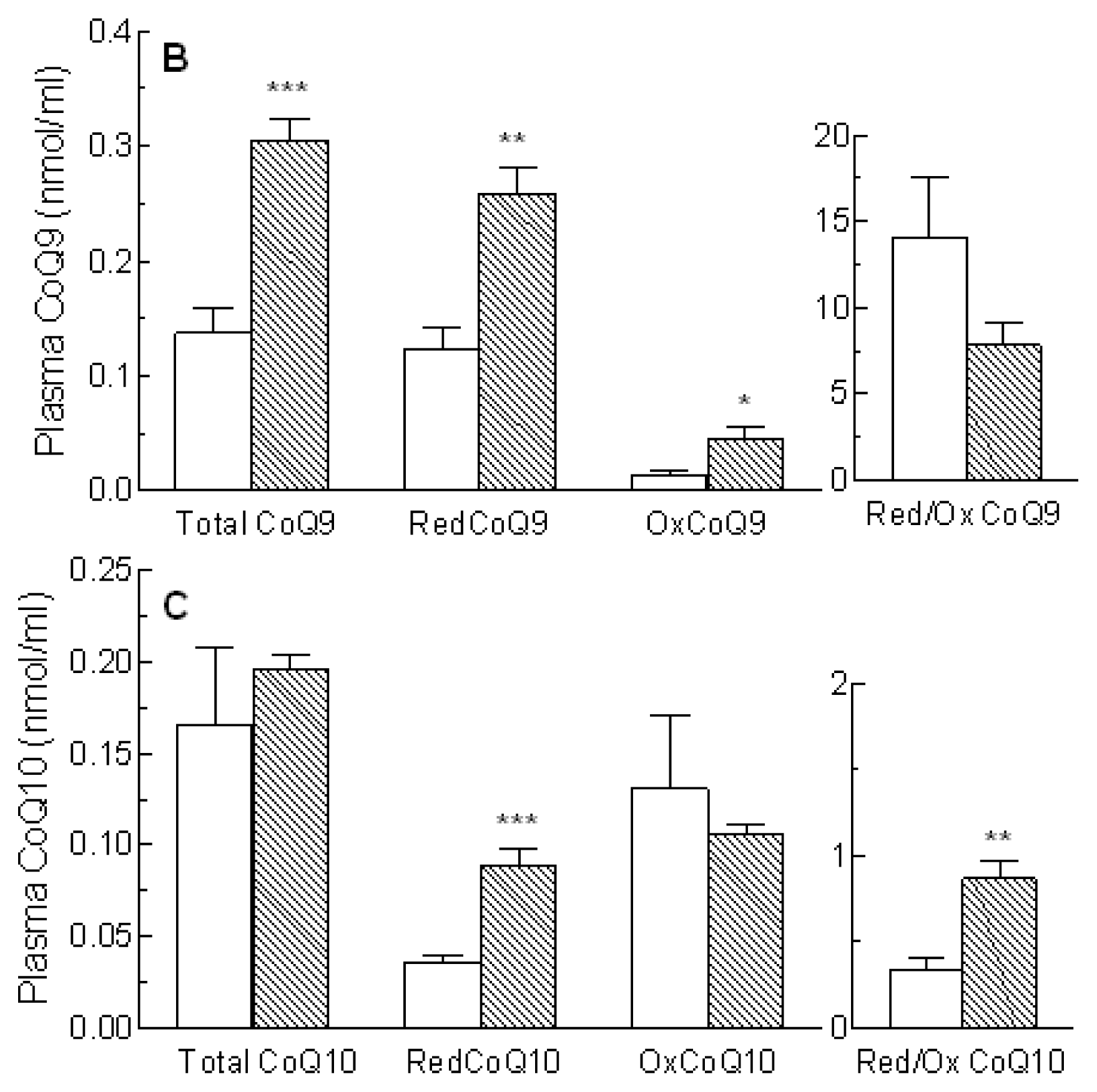
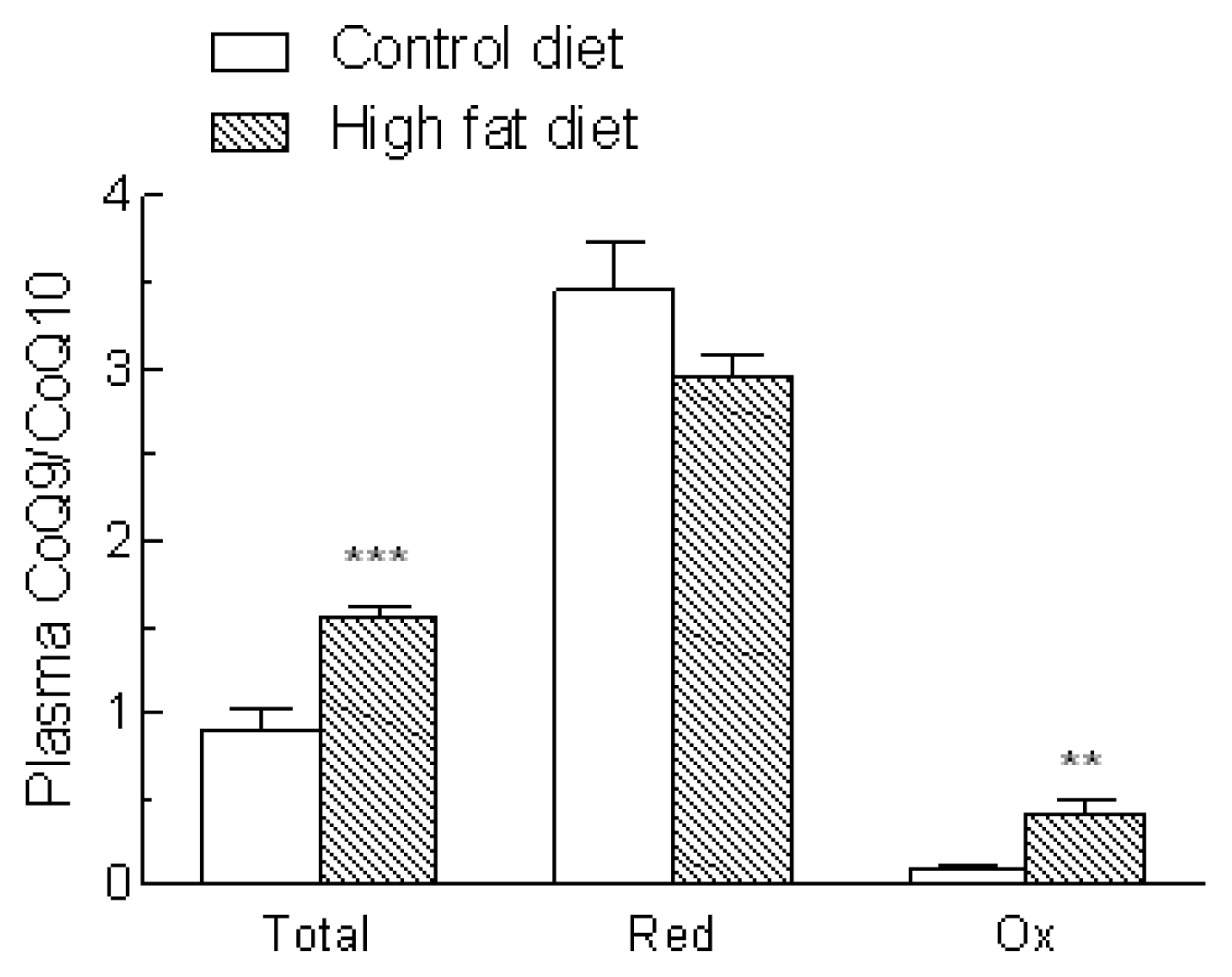
2.3. Coenzyme Q
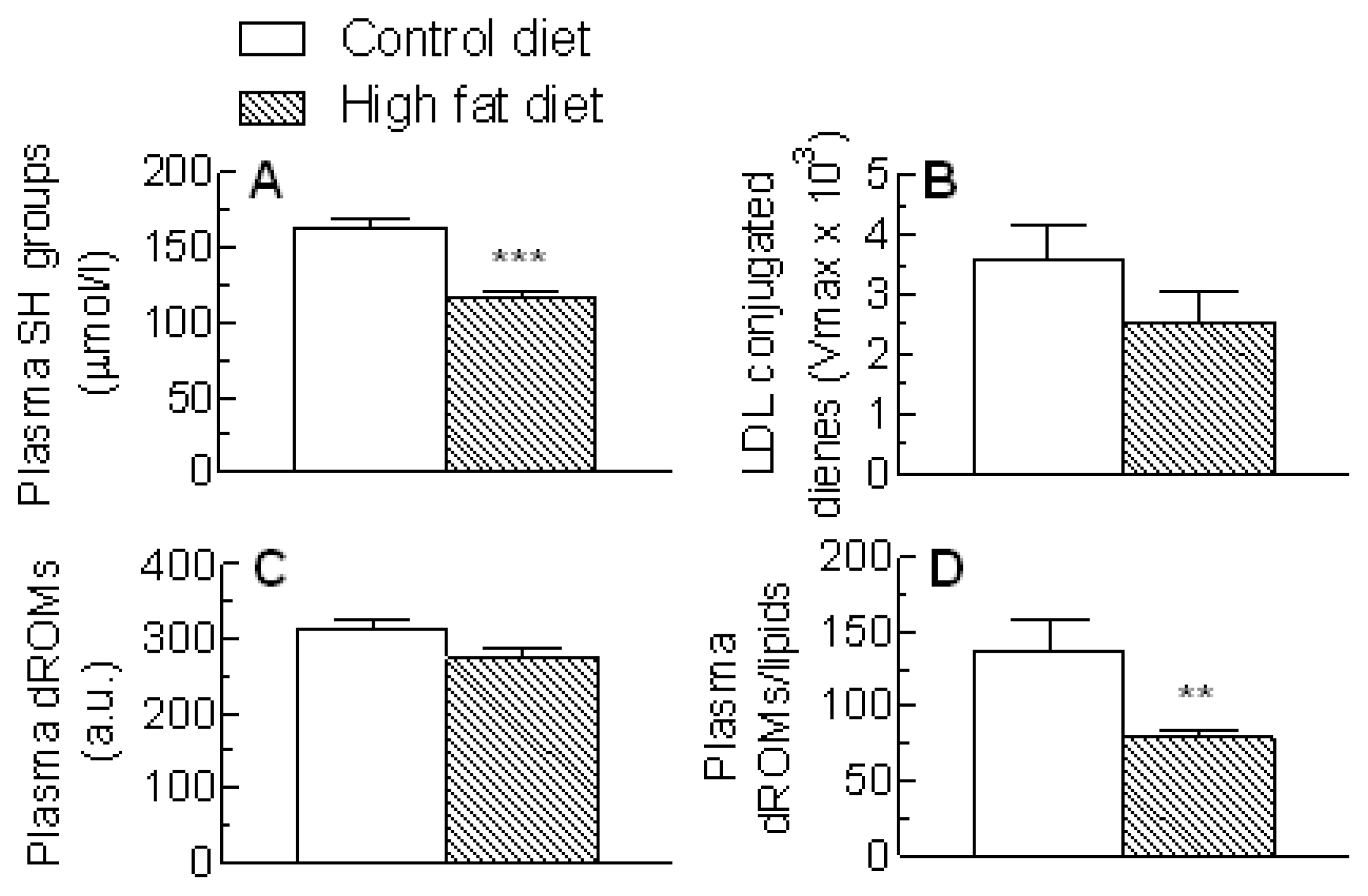
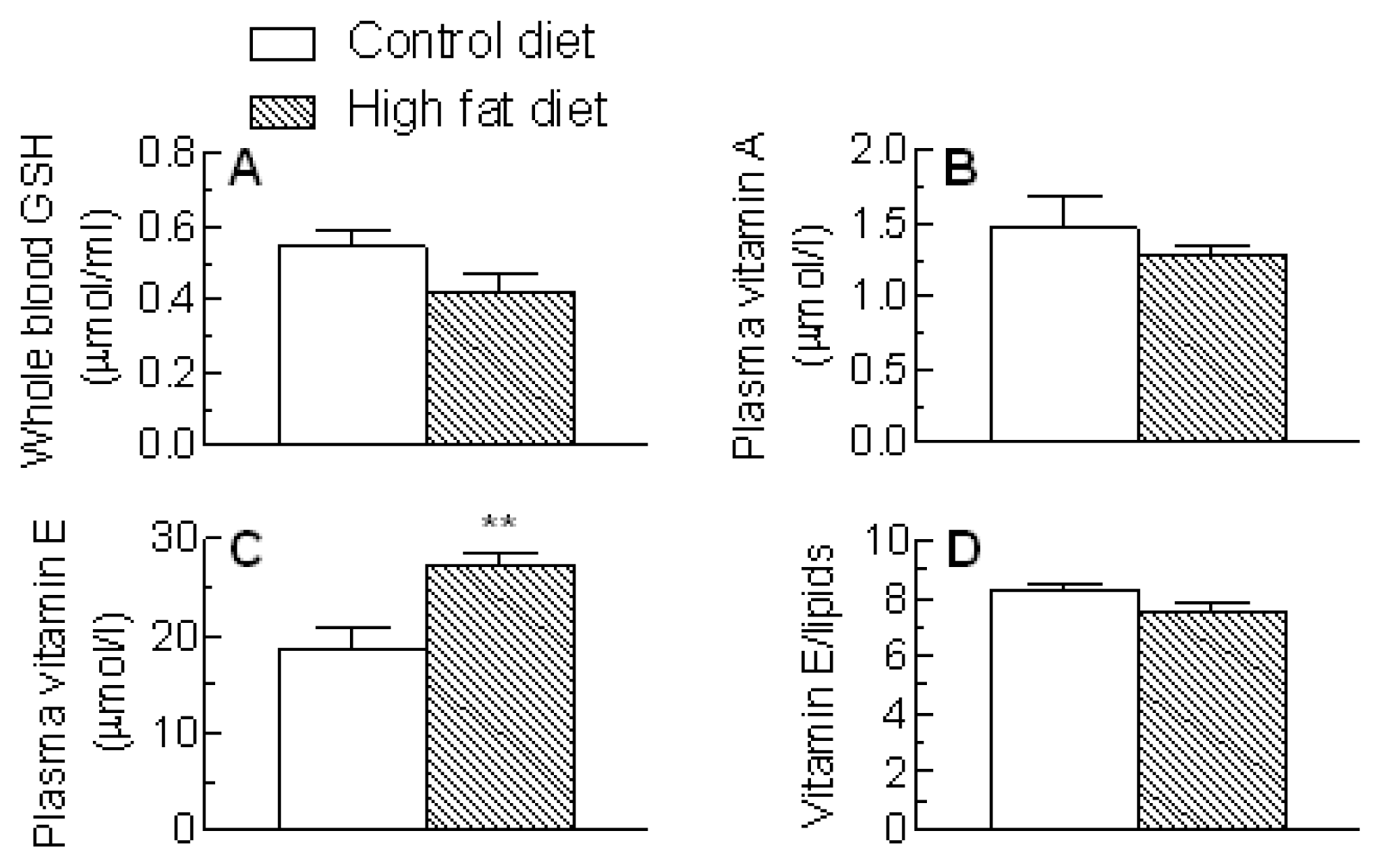
2.4. Blood Antioxidants and Oxidative Stress Markers
3. Experimental Section
3.1. Animals and Diets
3.2. Determination of Conjugated Dienes
3.3. Other Analytical Methods
4. Conclusions
Acknowledgments
References
- Adams, L.A.; Angulo, P. Recent concepts in non-alcoholic fatty liver disease. Diabet. Med 2005, 22, 1129–1133. [Google Scholar]
- Svegliati-Baroni, G.; Candelaresi, C.; Saccomanni, S.; Ferretti, G.; Bachetti, T.; Marzioni, M.; Minicis, S.D.; Nobili, L.; Salzano, R.; Omenetti, A.; et al. A model of insulin resistance and nonalcoholic steatohepatitis in rats. Am. J. Pathol 2006, 169, 846–860. [Google Scholar]
- Videla, L.A.; Rodrigo, R.; Araya, J.; Poniachik, J. Insulin resistance and oxidative stress interdependency in non-alcoholic fatty liver disease. Trends Mol. Med 2006, 12, 555–558. [Google Scholar]
- Day, C.P.; James, O.F.W. Steatohepatitis: A tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar]
- Anstee, Q.M.; Goldin, R.D. Mouse models in non-alcoholic fatty liver disease and steatohepatitis research. Int. J. Exp. Pathol 2006, 87, 1–16. [Google Scholar]
- Grattagliano, I.; Palmieri, V.O.; Portincasa, P.; Moschetta, A.; Palasciano, G. Oxidative stress-induced risk factors associated with the metabolic syndrome: A unifying hypothesis. J. Nutr. Biochem 2008, 19, 491–504. [Google Scholar]
- Videla, L.A.; Rodrigo, R.; Araya, J.; Poniachik, J. Oxidative stress and depletion of hepatic long-chain polyunsaturated fatty acids may contribute to nonalcoholic fatty liver disease. Free Radic. Biol. Med 2004, 37, 1499–1507. [Google Scholar]
- Videla, L.A.; Rodrigo, R.N.; Orellana, M.; Fernandez, V.; Tapia, G.; Quinones, L.; Varela, N.; Contreras, J.; Lazarte, R.; Csendes, A.; et al. Oxidative stress-related parameters in the liver of non-alcoholic fatty liver disease patients. Clin. Sci 2004, 106, 261–268. [Google Scholar]
- Day, C.P. Non-alcoholic fatty liver disease: Current concepts and management strategies. Clin. Med 2006, 6, 19–25. [Google Scholar]
- Araya, J.; Rodrigo, R.N.; Videla, L.A.; Thielemann, L.; Orellana, M.; Pettinelli, P.; Poniachik, J. Increase in long-chain polyunsaturated fatty acid n-6/n-3 ratio in relation to hepatic steatosis in patients with non-alcoholic fatty liver disease. Clin. Sci 2004, 106, 635–643. [Google Scholar]
- Tessari, P.; Coracina, A.; Cosma, A.; Tiengo, A. Hepatic lipid metabolism and non-alcoholic fatty liver disease. Nutr. Metab. Cardiovasc. Dis 2009, 19, 291–302. [Google Scholar]
- Buqué, X.; Martínez, M.J.; Cano, A; Miquilena-Colina, M.E.; García-Monzón, C.; Aspichueta, P.; Ochoa, B. A subset of dysregulated metabolic and survival genes is associated with severity of hepatic steatosis in obese Zucker rats. J. Lipid Res. 2010, 51, 500–513. [Google Scholar]
- Bentinger, M.; Brismar, K.; Dallner, G. The antioxidant role of coenzyme Q. Mitochondrion 2007, 7, S41–S50. [Google Scholar]
- Petrosillo, G.; Portincasa, P.; Grattagliano, I.; Casanova, G.; Matera, M.; Ruggiero, F.M.; Ferri, D.; Paradies, G. Mitochondrial dysfunction in rat with nonalcoholic fatty liver: Involvement of complex I, reactive oxygen species and cardiolipin. Biochim. Biophys. Acta 2007, 1767, 1260–1267. [Google Scholar]
- Safwat, G.M.; Pisanò, S.; D’Amore, E.; Borioni, G.; Napolitano, M.; Kamal, A.A.; Ballanti, P.; Botham, K.M.; Bravo, E. Induction of non-alcoholic fatty liver disease and insulin resistance by feeding a high-fat diet in rats: Does coenzyme Q monomethyl ether have a modulatory effect? Nutrition 2009, 25, 1157–1168. [Google Scholar]
- Koteish, A.; Diehl, A. Animal models of steatosis. Semin. Liver Dis 2001, 21, 89–104. [Google Scholar]
- Lieber, C.S.; Leo, M.A.; Mak, K.M.; Xu, Y.; Cao, Q.; Ren, C.; Ponomarenko, A.; de Carli, L.M. Model of nonalcoholic steatohepatitis. Am. J. Clin. Nutr 2004, 79, 502–509. [Google Scholar]
- Cano, A.; Ciaffoni, F.; Safwat, G.M.; Aspichueta, P.; Ochoa, B.; Bravo, E.; Botham, K.M. Hepatic very low density lipoprotein assembly is disturbed in a rat model of non alcoholic fatty liver disease: Is there a role for dietary Coenzyme Q? J. Appl. Physiol 2009, 107, 707–717. [Google Scholar]
- Adams, L.A.; Angulo, P.; Lindor, K.D. Nonalcoholic fatty liver disease. CMAJ 2005, 172, 899–905. [Google Scholar]
- Sorbi, D.; Boyton, J.; Lindor, K.D. The ratio of aspartate aminotransferase to alanine aminotransferase: Potential value in differentiating nonalcoholic steatohepatitis from alcoholic liver disease. Am. J. Gastroenterol 1999, 94, 1018–1022. [Google Scholar]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar]
- Hermans, N.; Cos, P.; de Meyer, G.R.Y.; Maes, L.; Pieters, L.; Vanden Berghe, D.; Vlietinck, A.J.; de Bruyne, T. Study of potential systemic oxidative stress animal models for the evaluation of antioxidant activity: Status of lipid peroxidation and fat-soluble antioxidants. J. Pharm. Pharmacol 2007, 59, 131–136. [Google Scholar]
- Ferreira, F.M.; Seiça, R.; Oliveira, P.J.; Coxito, P.M.; Moreno, A.J.; Palmeira, C.M.; Santos, M.S. Diabetes induces metabolic adaptations in rat liver mitochondria: Role of coenzyme Q and cardiolipin contents. Biochim. Biophys. Acta 2003, 1639, 113–120. [Google Scholar]
- Kohli, R.; Kirby, M.; Xanthakos, S.A.; Softic, S.; Feldstein, A.E.; Saxena, V.; Tang, P.H.; Miles, L.; Miles, M.V.; Balistreri, W.F.; et al. High-fructose, medium chain trans fat diet induces liver fibrosis and elevates plasma coenzyme Q9 in a novel murine model of obesity and nonalcoholic steatohepatitis. Hepatology 2010, 52, 934–944. [Google Scholar]
- Ernster, L.; Dallner, G. Biochemical, physiological and medical aspects of ubiquinone formation. Biochim. Biophys. Acta 1995, 1271, 195–204. [Google Scholar]
- Turunen, M.; Olsson, J.; Dallner, G. Metabolism and function of coenzyme Q. Biochim. Biophys. Acta 2004, 1660, 171–199. [Google Scholar]
- Quiles, J.A.; Huertas, J.R.; Manas, M.; Battino, M.; Cassinello, M.; Littarru, G.P.; Lenaz, G.; Mataix, F.J. Peroxidative extent and coenzyme Q levels in the rat: Influence of physical training and dietary fats. Mol. Aspects Med 1994, 15, 89–95. [Google Scholar]
- Ahmed, U.; Redgrave, T.G.; Oates, P.S. Effect of dietary fat to produce non-alcoholic fatty liver in the rat. J. Gastroenterol. Hepatol 2009, 24, 1463–1471. [Google Scholar]
- Zhu, M.J.; Sun, L.J.; Liu, Y.Q.; Feng, Y.L.; Tong, H.T.; Hu, Y.H.; Zhao, Z. Blood F2-isoprostanes are significantly associated with abnormalities of lipid status in rats with steatosis. World Gastroenterol. J 2008, 14, 4677–4683. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem 1957, 226, 497–509. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem 1972, 18, 499–502. [Google Scholar]
- Pinchuk, I.; Lichtenberg, D. The mechanism of action of antioxidants against lipoprotein peroxidation, evaluation based on kinetic experiments. Prog. Lipid Res 2002, 41, 279–314. [Google Scholar]
- Bradford, M. A sensitive method for quantitation of microgram quantities of protein utilizing the principle of dye-binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Balercia, G.; Mosca, F.; Mantero, F.; Boscaro, M.; Mancini, A.; Ricciardo-Lamonica, G.; Littarru, G. Coenzyme Q10 supplementation in infertile men with idiopathic asthenozoospermia: An open, uncontrolled pilot study. Fertil. Steril 2004, 81, 93–98. [Google Scholar]
- Nierenberg, D.W.; Lester, D.C. Determination of vitamins A and E in serum and plasma using a simplified clarification method and high-performance liquid chromatography. J. Chromatogr 1985, 345, 275–284. [Google Scholar]
- Pfeiffer, C.M.; Huff, D.L.; Gunter, E.W. Rapid and accurate HPLC assay for plasma total homocysteine and cysteine in a clinical laboratory setting. Clin. Chem 1999, 45, 290–292. [Google Scholar]
- Vassalle, C.; Boni, C.; di Cecco, P.; Ndreu, R.; Zucchelli, G.C. Automation and validation of a fast method for the assessment of in vivo oxidative stress levels. Clin. Chem. Lab. Med 2006, 44, 1372–1375. [Google Scholar]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol 1985, 113, 548–555. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control diet | High fat diet |
|---|---|---|
| Glucose (mmol/L) | 8.75 ± 0.74 | 8.07 ± 0.25 |
| ALT (U/L) | 33.5 ± 4.6 | 111.4+19.4 ** |
| AST (U/L) | 52.5 ± 5.1 | 61.1 ± 3.9 |
| AST/ALT | 1.65 ± 0.17 | 0.66 ± 0.07 *** |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bravo, E.; Palleschi, S.; Rossi, B.; Napolitano, M.; Tiano, L.; D’Amore, E.; Botham, K.M. Coenzyme Q Metabolism Is Disturbed in High Fat Diet-Induced Non Alcoholic Fatty Liver Disease in Rats. Int. J. Mol. Sci. 2012, 13, 1644-1657. https://doi.org/10.3390/ijms13021644
Bravo E, Palleschi S, Rossi B, Napolitano M, Tiano L, D’Amore E, Botham KM. Coenzyme Q Metabolism Is Disturbed in High Fat Diet-Induced Non Alcoholic Fatty Liver Disease in Rats. International Journal of Molecular Sciences. 2012; 13(2):1644-1657. https://doi.org/10.3390/ijms13021644
Chicago/Turabian StyleBravo, Elena, Simonetta Palleschi, Barbara Rossi, Mariarosaria Napolitano, Luca Tiano, Emanuela D’Amore, and Kathleen M Botham. 2012. "Coenzyme Q Metabolism Is Disturbed in High Fat Diet-Induced Non Alcoholic Fatty Liver Disease in Rats" International Journal of Molecular Sciences 13, no. 2: 1644-1657. https://doi.org/10.3390/ijms13021644