The Intriguing Life of Autophagosomes

Abstract
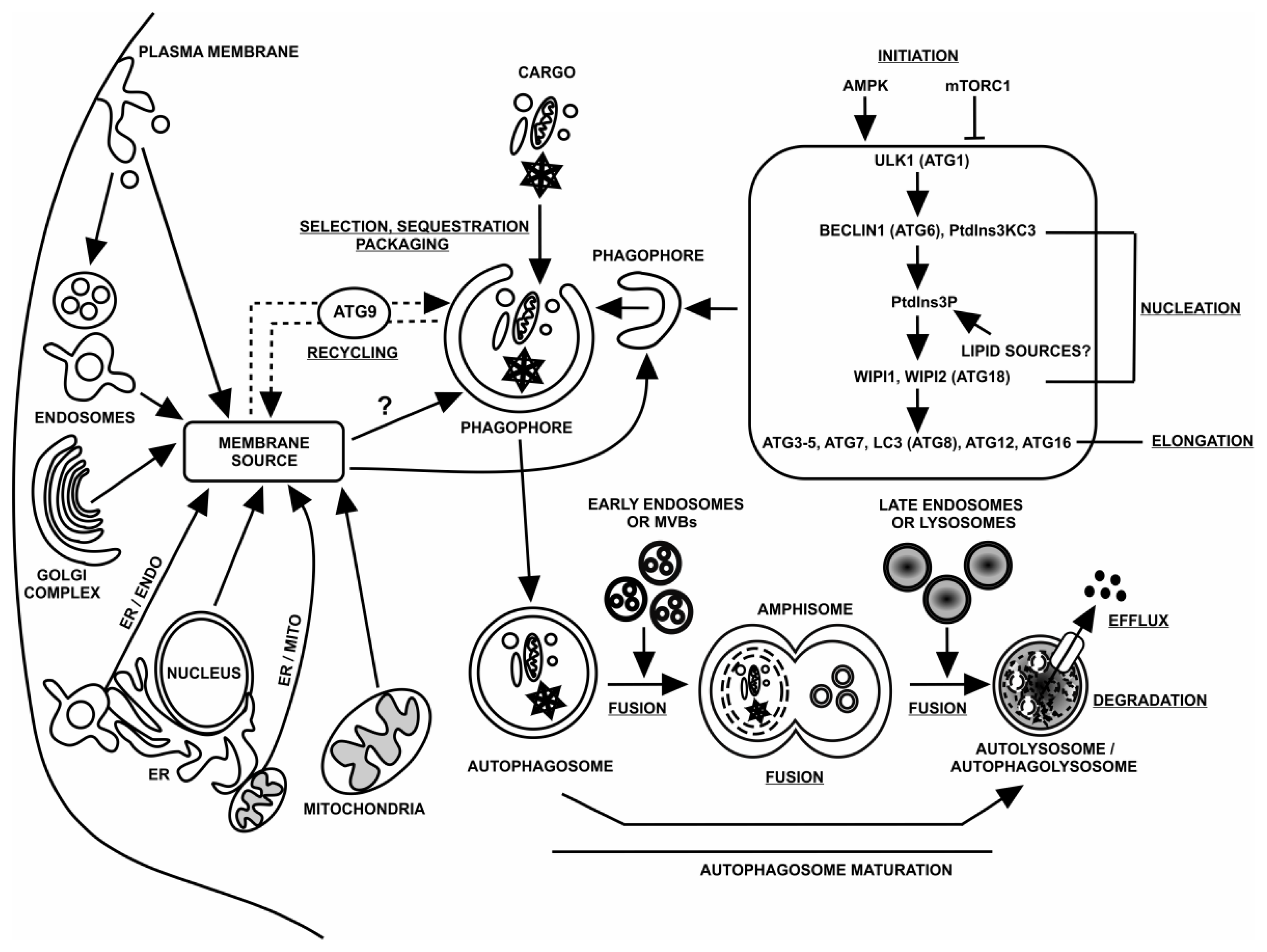
:1. Autophagy
2. Macroautophagy
3. Autophagosomes (APs)
3.1. Site(s) of Initiation
3.2. Structure
3.3. Source of Membrane Lipids
3.4. Trafficking and Fusion Events
4. Autophagosome Origin and Birth
4.1. ER and Mitochondrial Membranes
4.2. ER and Nuclear Membranes
4.3. Plasma Membrane
4.4. Golgi Complex and Endosomes
5. Phagophore Expansion and Autophagosome Development
6. Autophagosome Fusion Events
7. Outstanding Questions
7.1. AP Origin
7.2. AP Development
7.3. AP Maturation and Death
References
- Legakis, J.E.; Klionsky, D.J. Overview of Autophagy. In Autophagy in Immunity and Infection; Deretic, V., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germnay, 2006; Volume 1, Chapter 1; p. 3. [Google Scholar]
- Deretic, V.; Klionsky, D.J. How cells clean house. Sci. Am 2008, 298, 74–81. [Google Scholar]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075. [Google Scholar]
- Ravikumar, B.; Sarkar, S.; Davies, J.E.; Futter, M.; Garcia-Arencibia, M.; Green-Thompson, Z.W.; Jimenez-Sanchez, M.; Korolchuk, V.I.; Lichtenberg, M.; Luo, S.; et al. Regulation of mammalian autophagy in physiology and pathophysiology. Physiol. Rev 2010, 90, 1383–1435. [Google Scholar]
- Sridhar, S.; Botbol, Y.; Macian, F.; Cuervo, A.M. Autophagy and disease: Always two sides to a problem. J. Pathol 2011, 226, 255–273. [Google Scholar]
- Kaushik, S.; Bandyopadhyay, U.; Sridhar, S.; Kiffin, R.; Martinez-Vicente, M.; Kon, M.; Orenstein, S.J.; Wong, E.; Cuervo, A.M. Chaperone-mediated autophagy at a glance. J. Cell Sci 2011, 124, 495–499. [Google Scholar]
- Uttenweiler, A.; Mayer, A. Microautophagy in the yeast Saccharomyces cerevisiae. Methods Mol. Biol 2008, 445, 245–259. [Google Scholar]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. Microautophagy in mammalian cells: Revisiting a 40-year-old conundrum. Autophagy 2011, 7, 673–682. [Google Scholar]
- Li, W.W.; Li, J.; Bao, J.K. Microautophagy: Lesser-known self-eating. Cell. Mol. Life Sci 2011. [Google Scholar] [CrossRef]
- Sahu, R.; Kaushik, S.; Clement, C.C.; Cannizzo, E.S.; Scharf, B.; Follenzi, A.; Potolicchio, I.; Nieves, E.; Cuervo, A.M.; Santambrogio, L. Microautophagy of cytosolic proteins by late endosomes. Dev. Cell 2011, 20, 131–139. [Google Scholar]
- Santambrogio, L.; Cuervo, A.M. Chasing the elusive mammalian microautophagy. Autophagy 2011, 7, 652–654. [Google Scholar]
- Yang, Z.; Klionsky, D.J. Mammalian autophagy: Core molecular machinery and signaling regulation. Curr. Opin. Cell Biol 2010, 22, 124–131. [Google Scholar]
- Tooze, S.A.; Yoshimori, T. The origin of autophagosomal membrane. Nat. Cell Biol 2010, 12, 831–835. [Google Scholar]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. The role of atg proteins in autophagosome formation. Annu. Rev. Cell Dev. Biol 2011, 27, 107–132. [Google Scholar]
- Weidberg, H.; Shvets, E.; Elazar, Z. Biogenesis and cargo selectivity of autophagosomes. Annu. Rev. Biochem 2011, 80, 125–156. [Google Scholar]
- Mari, M.; Tooze, S.A.; Reggiori, F. The puzzling origin of the autophagosomal membrane. F1000 Biol. Rep 2011, 3. [Google Scholar] [CrossRef]
- Codogno, P.; Mehrpour, M.; Proikas-Cezanne, T. Canonical and non-canonical autophagy: Variations on a common theme of self-eating? Nat. Rev. Mol. Cell Biol 2011, 13, 7–12. [Google Scholar]
- Rubinsztein, D.C.; Shpilka, T.; Elazar, Z. Mechanisms of autophagosome biogenesis. Curr. Biol 2012, 22, R229–R234. [Google Scholar]
- He, C.; Klionsky, D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet 2009, 43, 67–93. [Google Scholar]
- Kanki, T.; Klionsky, D.J. The molecular mechanism of mitochondria autophagy in yeast. Mol. Microbiol 2010, 75, 795–800. [Google Scholar]
- Yorimitsu, T.; Klionsky, D.J. Autophagy: Molecular machinery of self-eating. Cell Death Differ 2005, 12, 1542–1552. [Google Scholar]
- Klionsky, D.J.; Cuervo, A.M.; Dunn, W.A., Jr; Levine, B.; van der Klei, I.; Seglen, P.O. How shall I eat thee? Autophagy 2007, 3, 413–416. [Google Scholar]
- Johansen, T.; Lamark, T. Selective autophagy mediated by autophagy adapter proteins. Autophagy 2011, 7, 279–296. [Google Scholar]
- Yamamoto, A.; Simonsen, A. The elimination of accumulated and aggregated proteins: A role for aggrephagy in neurodegeneration. Neurobiol. Dis 2011, 43, 17–28. [Google Scholar]
- Lamark, T.; Johansen, T. Aggrephagy: Selective disposal of protein aggregates by macroautophagy. Int. J. Cell Biol 2011, in press. [Google Scholar]
- Lynch-Day, M.A.; Klionsky, D.J. The Cvt pathway as a model for selective autophagy. FEBS Lett 2010, 584, 1359–1366. [Google Scholar]
- Umekawa, M.; Klionsky, D.J. The cytoplasm-to-vacuole targeting pathway: A historical perspective. Int. J. Cell Biol 2011, 2012. [Google Scholar] [CrossRef]
- Cebollero, E.; Reggiori, F.; Kraft, C. Reticulophagy and ribophagy: Regulated degradation of protein production factories. Int. J. Cell Biol 2011, 2012. [Google Scholar] [CrossRef]
- Hamasaki, M.; Noda, T.; Baba, M.; Ohsumi, Y. Starvation triggers the delivery of the endoplasmic reticulum to the vacuole via autophagy in yeast. Traffic 2005, 6, 56–65. [Google Scholar]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. Endoplasmic reticulum and Golgi complex: Contributions to, and turnover by, autophagy. Traffic 2006, 7, 1590–1595. [Google Scholar]
- Bernales, S.; Schuck, S.; Walter, P. ER-phagy: Selective autophagy of the endoplasmic reticulum. Autophagy 2007, 3, 285–287. [Google Scholar]
- Yorimitsu, T.; Klionsky, D.J. Eating the endoplasmic reticulum: Quality control by autophagy. Trends Cell Biol 2007, 17, 279–285. [Google Scholar]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar]
- Weidberg, H.; Shvets, E.; Elazar, Z. Lipophagy: Selective catabolism designated for lipids. Dev. Cell 2009, 16, 628–630. [Google Scholar]
- Rodriguez-Navarro, J.A.; Cuervo, A.M. Autophagy and lipids: Tightening the knot. Semin. Immunopathol 2010, 32, 343–353. [Google Scholar]
- Tolkovsky, A.M. Mitophagy. Biochim. Biophys. Acta 2009, 1793, 1508–1515. [Google Scholar]
- Kanki, T.; Klionsky, D.J.; Okamoto, K. Mitochondria autophagy in yeast. Antioxid. Redox Signal 2011, 14, 1989–2001. [Google Scholar]
- Youle, R.J.; Narendra, D.P. Mechanisms of mitophagy. Nat. Rev. Mol. Cell Biol 2011, 12, 9–14. [Google Scholar]
- May, A.I.; Rodney, J.D.; Prescott, M. The many faces of mitochondrial autophagy: Making sense of contrasting observations in recent research. Int. J. Cell Biol 2011, in press. [Google Scholar]
- Park, Y.E.; Hayashi, Y.K.; Bonne, G.; Arimura, T.; Noguchi, S.; Nonaka, I.; Nishino, I. Autophagic degradation of nuclear components in mammalian cells. Autophagy 2009, 5, 795–804. [Google Scholar]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. The intricacy of nuclear membrane dynamics during nucleophagy. Nucleus 2010, 1, 213–223. [Google Scholar]
- Dunn, W.A., Jr; Cregg, J.M.; Kiel, J.A.; van der Klei, I.J.; Oku, M.; Sakai, Y.; Sibirny, A.A.; Stasyk, O.V.; Veenhuis, M. Pexophagy: The selective autophagy of peroxisomes. Autophagy 2005, 1, 75–83. [Google Scholar]
- Sakai, Y.; Oku, M.; van der Klei, I.J.; Kiel, J.A. Pexophagy: Autophagic degradation of peroxisomes. Biochim. Biophys. Acta 2006, 1763, 1767–1775. [Google Scholar]
- Manjithaya, R.; Nazarko, T.Y.; Farré, J.C.; Subramani, S. Molecular mechanism and physiological role of pexophagy. FEBS Lett 2010, 584, 1367–1373. [Google Scholar]
- Oku, M.; Sakai, Y. Peroxisomes as dynamic organelles: Autophagic degradation. FEBS J 2010, 277, 3289–3294. [Google Scholar]
- Till, A.; Lakhani, R.; Burnett, S.F.; Subramani, S. Pexophagy—The selective degradation of peroxisomes. Int. J. Cell Biol 2011, in press. [Google Scholar]
- Roberts, P.; Moshitch-Moshkovitz, S.; Kvam, E.; O’Toole, E.; Winey, M.; Goldfarb, D.S. Piecemeal microautophagy of the nucleus in Saccharomyces cerevisiae. Mol. Biol. Cell 2003, 14, 129–141. [Google Scholar]
- Kvam, E.; Goldfarb, D.S. Nucleus-vacuole junctions and piecemeal microautophagy of the nucleus in S. cerevisiae. Autophagy 2007, 3, 85–92. [Google Scholar]
- Krick, R.; Muehe, Y.; Prick, T.; Bremer, S.; Schlotterhose, P.; Eskelinen, E.L.; Millen, J.; Goldfarb, D.S.; Thumm, M. Piecemeal microautophagy of the nucleus requires the core macroautophagy genes. Mol. Biol. Cell 2008, 19, 4492–4505. [Google Scholar]
- Kraft, C.; Deplazes, A.; Sohrmann, M.; Peter, M. Mature ribosomes are selectively degraded upon starvation by an autophagy pathway requiring the Ubp3p/Bre5p ubiquitin protease. Nat. Cell Biol 2008, 10, 602–610. [Google Scholar]
- Kraft, C.; Peter, M. Is the Rsp5 ubiquitin ligase involved in the regulation of ribophagy? Autophagy 2008, 4, 838–840. [Google Scholar]
- Tang, F.; Watkins, J.W.; Bermudez, M.; Gray, R.; Gaban, A.; Portie, K.; Grace, S.; Kleve, M.; Craciun, G. A life-span extending form of autophagy employs the vacuole-vacuole fusion machinery. Autophagy 2008, 4, 874–886. [Google Scholar]
- Brown, C.R.; Dunton, D.; Chiang, H.L. The vacuole import and degradation pathway utilizes early steps of endocytosis and actin polymerization to deliver cargo proteins to the vacuole for degradation. J. Biol. Chem 2010, 285, 1516–1528. [Google Scholar]
- Knodler, L.A.; Celli, J. Eating the strangers within: Host control of intracellular bacteria via xenophagy. Cell. Microbiol 2010, 13, 1319–1327. [Google Scholar]
- Itakura, E.; Mizushima, N. Characterization of autophagosome formation site by a hierarchical analysis of mammalian Atg proteins. Autophagy 2010, 6, 764–776. [Google Scholar]
- Chen, Y.; Klionsky, D.J. The regulation of autophagy—Unanswered questions. J. Cell Sci 2011, 124, 161–170. [Google Scholar]
- Suzuki, K.; Kirisako, T.; Kamada, Y.; Mizushima, N.; Noda, T.; Ohsumi, Y. The pre-autophagosomal structure organized by concerted functions of APG genes is essential for autophagosome formation. EMBO J 2001, 20, 5971–5981. [Google Scholar]
- Suzuki, K.; Ohsumi, Y. Current knowledge of the pre-autophagosomal structure (PAS). FEBS Lett 2010, 584, 1280–1286. [Google Scholar]
- Baba, M.; Osumi, M.; Ohsumi, Y. Analysis of the membrane structures involved in autophagy in yeast by freeze-replica method. Cell. Struct. Funct 1995, 20, 465–471. [Google Scholar]
- Suzuki, K.; Ohsumi, Y. Molecular machinery of autophagosome formation in yeast, Saccharomyces cerevisiae. FEBS Lett 2007, 581, 2156–2161. [Google Scholar]
- Øverbye, A.; Fengsrud, M.; Seglen, P.O. Proteomic analysis of membrane-associated proteins from rat liver autophagosomes. Autophagy 2007, 3, 300–322. [Google Scholar]
- Mizushima, N. Autophagy: Process and function. Genes Dev 2007, 21, 2861–2873. [Google Scholar]
- Takeshige, K.; Baba, M.; Tsuboi, S.; Noda, T.; Ohsumi, Y. Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. J. Cell Biol 1992, 119, 301–311. [Google Scholar]
- Pfeifer, U. Inhibition by insulin of the formation of autophagic vacuoles in rat liver. A morphometric approach to the kinetics of intracellular degradation by autophagy. J. Cell Biol 1978, 78, 152–167. [Google Scholar]
- Schworer, C.M.; Shiffer, K.A.; Mortimore, G.E. Quantitative relationship between autophagy and proteolysis during graded amino acid deprivation in perfused rat liver. J. Biol. Chem 1981, 256, 7652–7658. [Google Scholar]
- Mizushima, N.; Klionsky, D.J. Protein turnover via autophagy: Implications for metabolism. Annu. Rev. Nutr 2007, 27, 19–40. [Google Scholar]
- Mortimore, G.E.; Pösö, A.R. Intracellular protein catabolism and its control during nutrient deprivation and supply. Annu. Rev. Nutr 1987, 7, 539–564. [Google Scholar]
- Kimura, S.; Noda, T.; Yoshimori, T. Dynein-dependent movement of autophagosomes mediates efficient encounters with lysosomes. Cell. Struct. Funct 2008, 33, 109–122. [Google Scholar]
- Axe, E.L.; Walker, S.A.; Manifava, M.; Chandra, P.; Roderick, H.L.; Habermann, A.; Griffiths, G.; Ktistakis, N.T. Autophagosome formation from membrane compartments enriched in phosphatidylinositol 3-phosphate and dynamically connected to the endoplasmic reticulum. J. Cell Biol 2008, 182, 685–701. [Google Scholar]
- Simonsen, A.; Stenmark, H. Self-eating from an ER-associated cup. J. Cell Biol 2008, 182, 621–622. [Google Scholar]
- Hayashi-Nishino, M.; Fujita, N.; Noda, T.; Yamaguchi, A.; Yoshimori, T.; Yamamoto, A. A subdomain of the endoplasmic reticulum forms a cradle for autophagosome formation. Nat. Cell Biol 2009, 11, 1433–1437. [Google Scholar]
- Ylä-Anttila, P.; Vihinen, H.; Jokitalo, E.; Eskelinen, E.L. 3D tomography reveals connections between the phagophore and endoplasmic reticulum. Autophagy 2009, 5, 1180–1185. [Google Scholar]
- Hailey, D.W.; Rambold, A.S.; Satpute-Krishnan, P.; Mitra, K.; Sougrat, R.; Kim, P.K.; Lippincott-Schwartz, J. Mitochondria supply membranes for autophagosome biogenesis during starvation. Cell 2010, 141, 656–667. [Google Scholar]
- McEwan, D.G.; Dikic, I. Not all autophagy membranes are created equal. Cell 2010, 141, 564–566. [Google Scholar]
- Rambold, A.S.; Lippincott-Schwartz, J. Starved cells use mitochondria for autophagosome biogenesis. Cell Cycle 2010, 9, 3633–3634. [Google Scholar]
- English, L.; Chemali, M.; Duron, J.; Rondeau, C.; Laplante, A.; Gingras, D.; Alexander, D.; Leib, D.; Norbury, C.; Lippé, R.; et al. Autophagy enhances the presentation of endogenous viral antigens on MHC class I molecules during HSV-1 infection. Nat. Immunol 2009, 10, 480–487. [Google Scholar]
- English, L.; Chemali, M.; Desjardins, M. Nuclear membrane-derived autophagy, a novel process that participates in the presentation of endogenous viral antigens during HSV-1 infection. Autophagy 2009, 5, 1026–1029. [Google Scholar]
- Ravikumar, B.; Moreau, K.; Jahreiss, L.; Puri, C.; Rubinsztein, D.C. Plasma membrane contributes to the formation of pre-autophagosomal structures. Nat. Cell Biol 2010, 12, 747–757. [Google Scholar]
- Ravikumar, B.; Moreau, K.; Rubinsztein, D.C. Plasma membrane helps autophagosomes grow. Autophagy 2010, 6, 1184–1186. [Google Scholar]
- Cuervo, A.M. The plasma membrane brings autophagosomes to life. Nat. Cell Biol 2010, 12, 735–737. [Google Scholar]
- Geng, J.; Klionsky, D.J. The Golgi as a potential membrane source for autophagy. Autophagy 2010, 6, 950–951. [Google Scholar]
- Mari, M.; Reggiori, F. Atg9 reservoirs, a new organelle of the yeast endomembrane system? Autophagy 2010, 6, 1221–1223. [Google Scholar]
- Mari, M.; Griffith, J.; Rieter, E.; Krishnappa, L.; Klionsky, D.J.; Reggiori, F. An Atg9-containing compartment that functions in the early steps of autophagosome biogenesis. J. Cell Biol 2010, 190, 1005–1022. [Google Scholar]
- Tooze, S.A.; Jefferies, H.B.; Kalie, E.; Longatti, A.; McAlpine, F.E.; McKnight, N.C.; Orsi, A.; Polson, H.E.; Razi, M.; Robinson, D.J.; Webber, J.L. Trafficking and signaling in mammalian autophagy. IUBMB Life 2010, 62, 503–508. [Google Scholar]
- Takahashi, Y.; Meyerkord, C.L.; Hori, T.; Runkle, K.; Fox, T.E.; Kester, M.; Loughran, T.P.; Wang, H.G. Bif-1 regulates Atg9 trafficking by mediating the fission of Golgi membranes during autophagy. Autophagy 2011, 7, 61–73. [Google Scholar]
- Ohsumi, Y. Molecular dissection of autophagy: Two ubiquitin-like systems. Nat. Rev. Mol. Cell Biol 2001, 2, 211–216. [Google Scholar]
- Mizushima, N.; Noda, T.; Yoshimori, T.; Tanaka, Y.; Ishii, T.; George, M.D.; Klionsky, D.J.; Ohsumi, M.; Ohsumi, Y. A protein conjugation system essential for autophagy. Nature 1998, 395, 395–398. [Google Scholar]
- Kim, J.; Dalton, V.M.; Eggerton, K.P.; Scott, S.V.; Klionsky, D.J. Apg7p/Cvt2p is required for the cytoplasm-to-vacuole targeting, macroautophagy, and peroxisome degradation pathways. Mol. Biol. Cell 1999, 10, 1337–1351. [Google Scholar]
- Shintani, T.; Mizushima, N.; Ogawa, Y.; Matsuura, A.; Noda, T.; Ohsumi, Y. Apg10p, a novel protein-conjugating enzyme essential for autophagy in yeast. EMBO J 1999, 18, 5234–5241. [Google Scholar]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. Role of the Apg12 conjugation system in mammalian autophagy. Int. J. Biochem. Cell Biol 2003, 35, 553–561. [Google Scholar]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; Noda, T.; Ohsumi, Y. A ubiquitin-like system mediates protein lipidation. Nature 2000, 408, 488–492. [Google Scholar]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 conjugation system in mammalian autophagy. Int. J. Biochem. Cell Biol 2004, 36, 2503–2518. [Google Scholar]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar]
- Fujita, N.; Itoh, T.; Omori, H.; Fukuda, M.; Noda, T.; Yoshimori, T. The Atg16L complex specifies the site of LC3 lipidation for membrane biogenesis in autophagy. Mol. Biol. Cell 2008, 19, 2092–2100. [Google Scholar]
- Nemoto, T.; Tanida, I.; Tanida-Miyake, E.; Minematsu-Ikeguchi, N.; Yokota, M.; Ohsumi, M.; Ueno, T.; Kominami, E. The mouse APG10 homologue, an E2-like enzyme for Apg12p conjugation, facilitates MAP-LC3 modification. J. Biol. Chem 2003, 278, 39517–39526. [Google Scholar]
- Jimenez-Sanchez, M.; Thompson, F.; Zavodsky, E.; Rubinsztein, D.C. Autophagy and polyglutamine diseases. Prog. Neurobiol. 2011, in press. [Google Scholar]
- Nakatogawa, H.; Ichimura, Y.; Ohsumi, Y. Atg8, a ubiquitin-like protein required for autophagosome formation, mediates membrane tethering and hemifusion. Cell 2007, 130, 165–178. [Google Scholar]
- Subramani, S.; Farré, J.C. A ubiquitin-like protein involved in membrane fusion. Cell 2007, 130, 18–20. [Google Scholar]
- Xin, Y.; Yu, L.; Chen, Z.; Zheng, L.; Fu, Q.; Jiang, J.; Zhang, P.; Gong, R.; Zhao, S. Cloning, expression patterns, and chromosome localization of three human and two mouse homologues of GABA(A) receptor-associated protein. Genomics 2001, 74, 408–413. [Google Scholar]
- He, H.; Dang, Y.; Dai, F.; Guo, Z.; Wu, J.; She, X.; Pei, Y.; Chen, Y.; Ling, W.; Wu, C.; et al. Post-translational modifications of three members of the human MAP1LC3 family and detection of a novel type of modification for MAP1LC3B. J. Biol. Chem 2003, 278, 29278–29287. [Google Scholar]
- Bjørkøy, G.; Lamark, T.; Brech, A.; Outzen, H.; Perander, M.; Overvatn, A.; Stenmark, H.; Johansen, T. p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J. Cell Biol 2005, 171, 603–614. [Google Scholar]
- Kirkin, V.; Lamark, T.; Sou, Y.S.; Bjørkøy, G.; Nunn, J.L.; Bruun, J.A.; Shvets, E.; McEwan, D.G.; Clausen, T.H.; Wild, P.; et al. A role for NBR1 in autophagosomal degradation of ubiquitinated substrates. Mol. Cell 2009, 33, 505–516. [Google Scholar]
- Sagiv, Y.; Legesse-Miller, A.; Porat, A.; Elazar, Z. GATE-16, a membrane transport modulator, interacts with NSF and the Golgi v-SNARE GOS-28. EMBO J 2000, 19, 1494–1504. [Google Scholar]
- Kittler, J.T.; Rostaing, P.; Schiavo, G.; Fritschy, J.M.; Olsen, R.; Triller, A.; Moss, S.J. The subcellular distribution of GABARAP and its ability to interact with NSF suggest a role for this protein in the intracellular transport of GABA(A) receptors. Mol. Cell Neurosci 2001, 18, 13–25. [Google Scholar]
- Kabeya, Y.; Mizushima, N.; Yamamoto, A.; Oshitani-Okamoto, S.; Ohsumi, Y.; Yoshimori, T. LC3, GABARAP and GATE16 localize to autophagosomal membrane depending on form-II formation. J. Cell Sci 2004, 117, 2805–2812. [Google Scholar]
- Weidberg, H.; Shvets, E.; Shpilka, T.; Shimron, F.; Shinder, V.; Elazar, Z. LC3 and GATE-16/GABARAP subfamilies are both essential yet act differently in autophagosome biogenesis. EMBO J 2010, 29, 1792–1802. [Google Scholar]
- Monastyrska, I.; Rieter, E.; Klionsky, D.J.; Reggiori, F. Multiple roles of the cytoskeleton in autophagy. Biol. Rev. Camb. Philos. Soc 2009, 84, 431–448. [Google Scholar]
- Nakamura, N.; Matsuura, A.; Wada, Y.; Ohsumi, Y. Acidification of vacuoles is required for autophagic degradation in the yeast, Saccharomyces cerevisiae. J. Biochem 1997, 121, 338–344. [Google Scholar]
- Epple, U.D.; Suriapranata, I.; Eskelinen, E.L.; Thumm, M. Aut5/Cvt17p, a putative lipase essential for disintegration of autophagic bodies inside the vacuole. J. Bacteriol 2001, 183, 5942–5955. [Google Scholar]
- Teter, S.A.; Eggerton, K.P.; Scott, S.V.; Kim, J.; Fischer, A.M.; Klionsky, D.J. Degradation of lipid vesicles in the yeast vacuole requires function of Cvt17, a putative lipase. J. Biol. Chem 2001, 276, 2083–2087. [Google Scholar]
- Yang, Z.; Huang, J.; Geng, J.; Nair, U.; Klionsky, D.J. Atg22 recycles amino acids to link the degradative and recycling functions of autophagy. Mol. Biol. Cell 2006, 17, 5094–5104. [Google Scholar]
- Yang, Z.; Klionsky, D.J. Permeases recycle amino acids resulting from autophagy. Autophagy 2007, 3, 149–150. [Google Scholar]
- Jahreiss, L.; Menzies, F.M.; Rubinsztein, D.C. The itinerary of autophagosomes: From peripheral formation to kiss-and-run fusion with lysosomes. Traffic 2008, 9, 574–587. [Google Scholar]
- Ravikumar, B.; Futter, M.; Jahreiss, L.; Korolchuk, V.I.; Lichtenberg, M.; Luo, S.; Massey, D.C.; Menzies, F.M.; Narayanan, U.; Renna, M.; et al. Mammalian macroautophagy at a glance. J. Cell Sci 2009, 122, 1707–1711. [Google Scholar]
- Koga, H.; Kaushik, S.; Cuervo, A.M. Altered lipid content inhibits autophagic vesicular fusion. FASEB J 2010, 24, 3052–3065. [Google Scholar]
- Mehrpour, M.; Esclatine, A.; Beau, I.; Codogno, P. Overview of macroautophagy regulation in mammalian cells. Cell Res 2010, 20, 748–762. [Google Scholar]
- Eskelinen, E.L. Maturation of autophagic vacuoles in mammalian cells. Autophagy 2005, 1, 1–10. [Google Scholar]
- Noda, T.; Fujita, N.; Yoshimori, T. The late stages of autophagy: How does the end begin? Cell Death Differ 2009, 16, 984–990. [Google Scholar]
- Fader, C.M.; Sánchez, D.; Furlán, M.; Colombo, M.I. Induction of autophagy promotes fusion of multivesicular bodies with autophagic vacuoles in K562 cells. Traffic 2008, 9, 230–250. [Google Scholar]
- Liang, C.; Lee, J.S.; Inn, K.S.; Gack, M.U.; Li, Q.; Roberts, E.A.; Vergne, I.; Deretic, V.; Feng, P.; Akazawa, C.; et al. Beclin1-binding UVRAG targets the class C Vps complex to coordinate autophagosome maturation and endocytic trafficking. Nat. Cell Biol 2008, 10, 776–787. [Google Scholar]
- Matsunaga, K.; Saitoh, T.; Tabata, K.; Omori, H.; Satoh, T.; Kurotori, N.; Maejima, I.; Shirahama-Noda, K.; Ichimura, T.; Isobe, T.; et al. Two Beclin 1-binding proteins, Atg14L and Rubicon, reciprocally regulate autophagy at different stages. Nat. Cell Biol 2009, 11, 385–396. [Google Scholar]


{kind=link}
| Selective type of autophagy | Cargo | Organism | References |
|---|---|---|---|
| Aggrephagy | Protein aggregates | Mammals | [22–25] |
| Cytoplasm-to-vacuole targeting (Cvt) pathway | Pro-aminopeptidase 1 (prApe1), pro-α mannosidase 1 (prAms1) and aspartyl aminopeptidase (Ape4) | Yeast | [22,26–28] |
| ER-phagy/reticulophagy | ER | Yeast and Mammals | [22,28–32] |
| Lipophagy | Lipids | Mammals | [33–35] |
| Lysophagy/Lysosomophagy | Vacuole/Lysosomal membrane | Yeast and Mammals | [22] |
| Mitophagy | Mitochondria | Yeast and Mammals | [20,22,36–39] |
| Nucleophagy | Nucleus | Yeast and Mammals | [22,40,41] |
| Pexophagy | Peroxisomes/peroxisome cluster | Yeast and Mammals | [22,42–46] |
| Piecemeal-microautophagy of the nucleus (PMN) | Portions of the nucleus | Yeast | [22,47–49] |
| Ribophagy | Ribosomes | Yeast | [28,50,51] |
| Secretophagy | Atg15 protein | Yeast | [52] |
| Vacuole import and degradation (Vid) pathway | Fructose-1,6-bisphosphatase (FBPase) | Yeast | [22,53] |
| Xenophagy | Pathogens (bacteria and viruses) | Plants and Mammals | [22,23,54] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mijaljica, D.; Prescott, M.; Devenish, R.J. The Intriguing Life of Autophagosomes. Int. J. Mol. Sci. 2012, 13, 3618-3635. https://doi.org/10.3390/ijms13033618
Mijaljica D, Prescott M, Devenish RJ. The Intriguing Life of Autophagosomes. International Journal of Molecular Sciences. 2012; 13(3):3618-3635. https://doi.org/10.3390/ijms13033618
Chicago/Turabian StyleMijaljica, Dalibor, Mark Prescott, and Rodney J. Devenish. 2012. "The Intriguing Life of Autophagosomes" International Journal of Molecular Sciences 13, no. 3: 3618-3635. https://doi.org/10.3390/ijms13033618