Isolation and Purification of a Novel Deca-Antifungal Peptide from Potato (Solanum tuberosum L. cv. Jopung) Against Candida albicans

Abstract
:1. Introduction
2. Results and Discussion
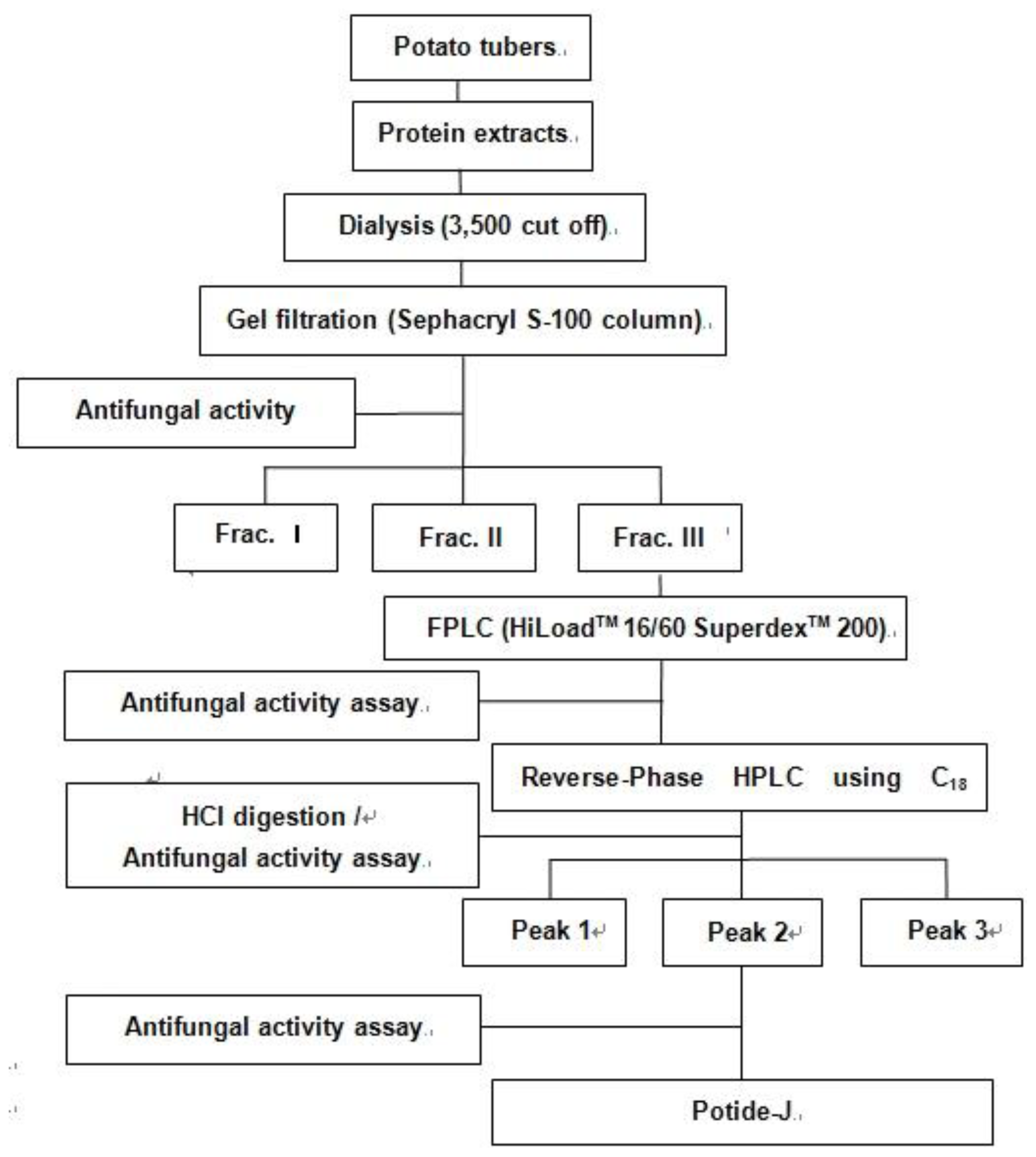
2.1. Purification of the Antifungal Peptide, Potide-J
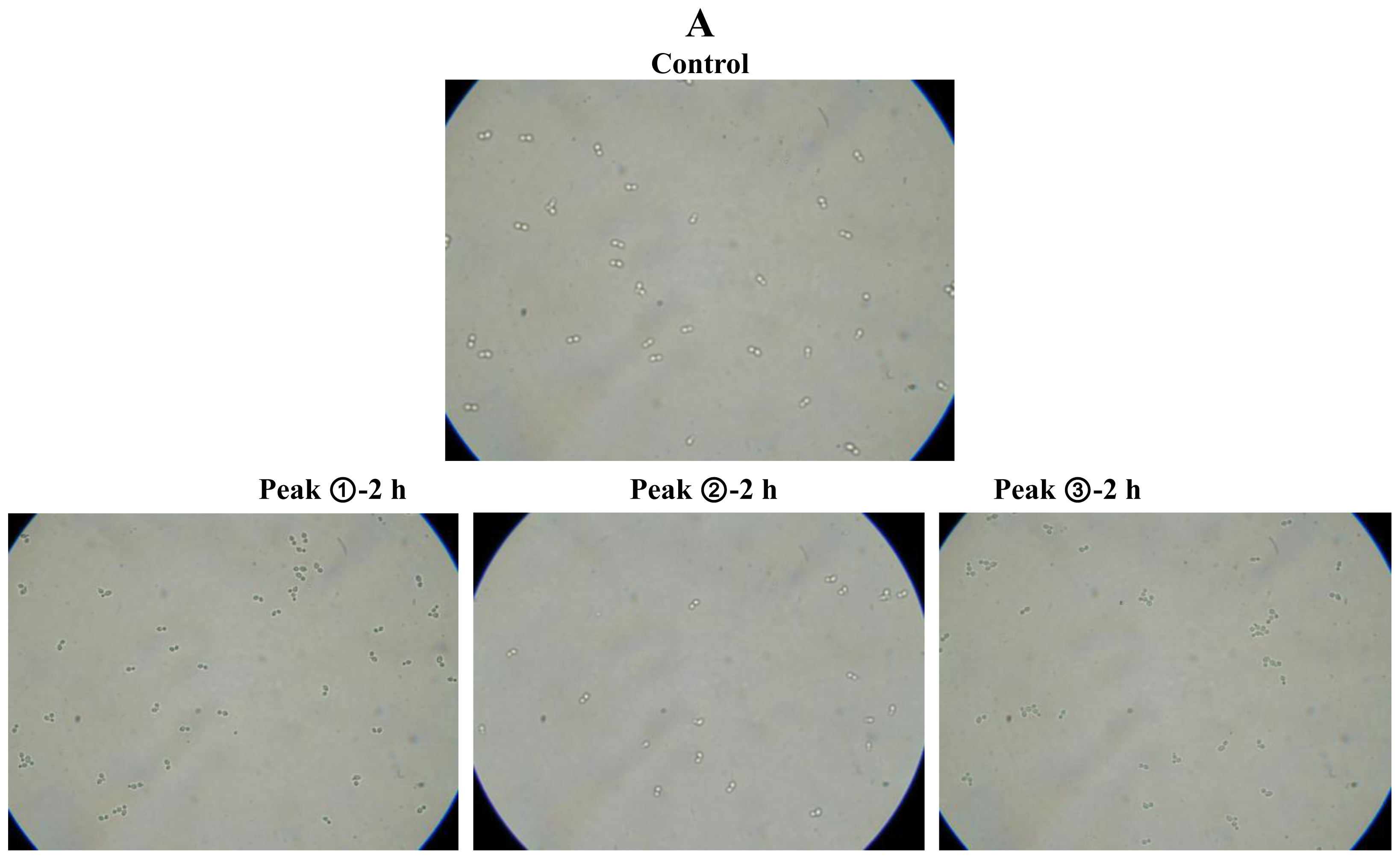
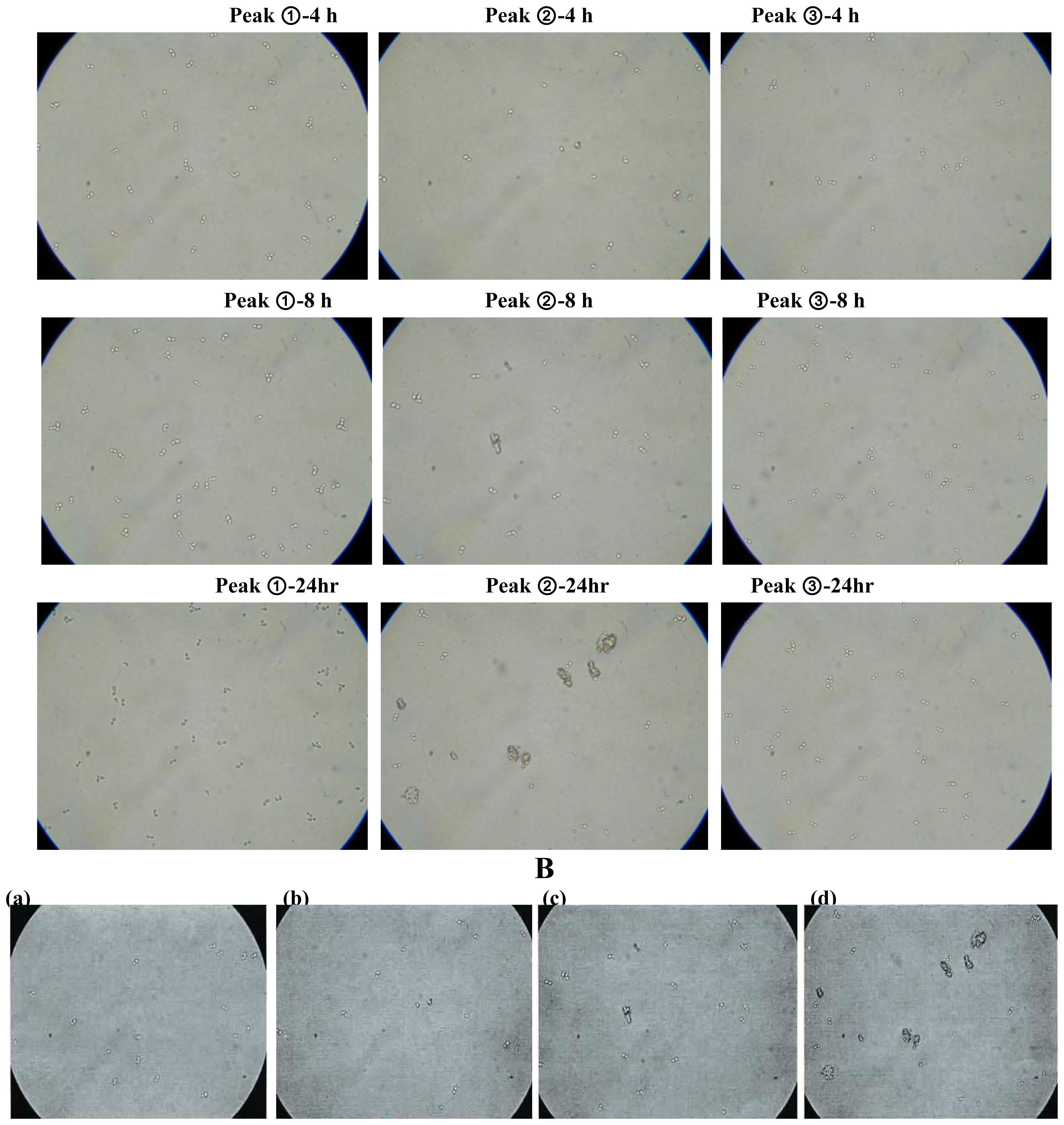
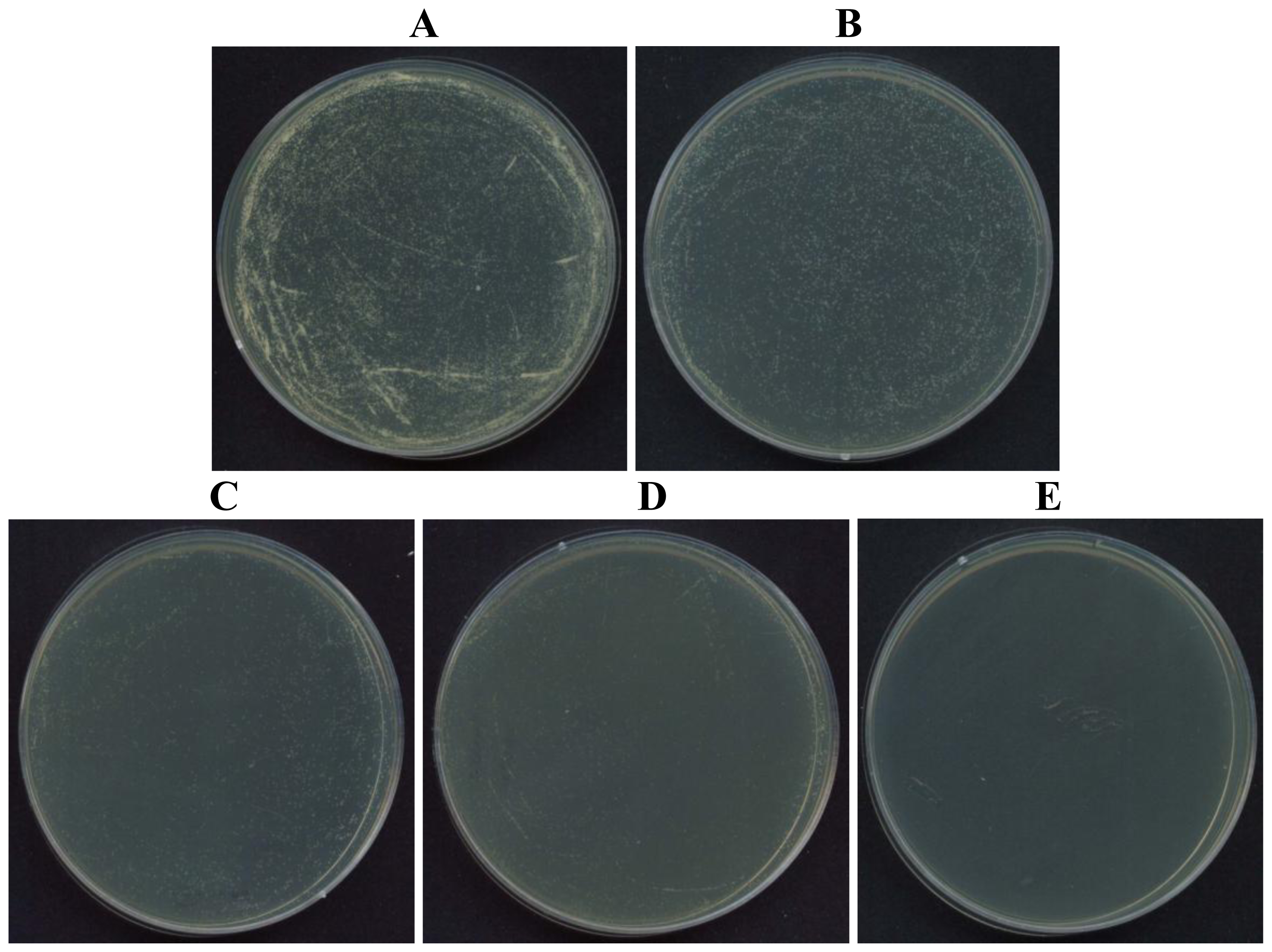
2.2. Antifungal and Non-hemolytic Effects of Potide-J
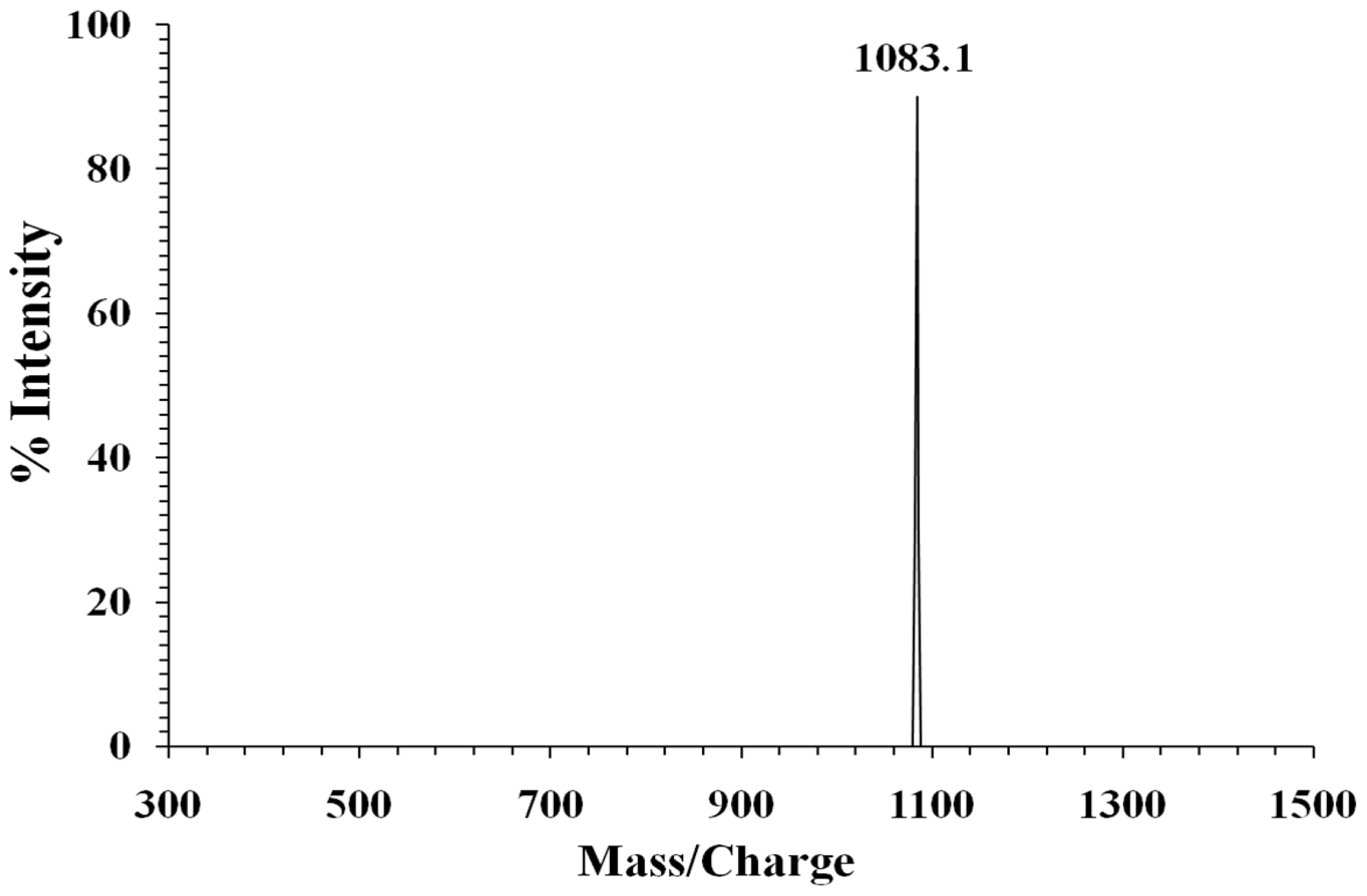
2.3. Protein Identification
3. Experimental Section
3.1. Potato Tubers
3.2. Step I: Preparation of AFP-J
3.3. Step II: Preparation of Potide-J
3.4. Assay for Antifungal Activity
3.5. Visualize on Agar Plate of Synthetic Peptides
3.6. N-terminal Amino Acid Sequencing
3.7. Mass Spectrometry
3.8. Peptide Synthesis and Purification
4. Conclusions
Acknowledgements
References and Notes
- Birk, Y. The Bowman-Birk inhibitor, trypsin- and chymotrypsin-inhibitor from soybeans. Int. J. Pept. Protein Res 1985, 25, 113–131. [Google Scholar]
- Ye, X.Y.; Wang, H.X.; Ng, T.B. First chromatographic isolation of an antifungal thaumatin-like protein from French bean legumes and demonstration of its antifungal activity. Biochem. Biophys. Res. Commun 1999, 263, 130–134. [Google Scholar]
- Benhamou, N.; Broglie, K.; Broglie, R.; Chet, I. Antifungal effect of bean endochitinase on Rhizoctonia solani: Ultrastructural changes and cytochemical aspects of chitin breakdown. Can. J. Microbiol 1993, 39, 318–328. [Google Scholar]
- Vogelsang, R.; Barz, W. Purification, characterization and differential hormonal regulation of a β-1,3-glucanase and two chitinases from chickpea (Cicer arietinum L.). Planta 1993, 189, 60–69. [Google Scholar]
- Kim, J.Y.; Park, S.C.; Kim, M.H.; Lim, H.T.; Park, Y.; Hahm, K.S. Antimicrobial activity studies on a trypsin chymotrypsin protease inhibitor obtained from potato. Biochem. Biophys. Res. Commun 2005, 330, 921–927. [Google Scholar]
- Khalf, M.; Goulet, C.; Vorster, J.; Brunelle, F.; Anguenot, R.; Fliss, I.; Michaud, D. Tubers from potato lines expressing a tomato Kunitz protease inhibitor are substantially equivalent to parental and transgenic controls. Plant Biotechnol. J 2010, 8, 155–169. [Google Scholar]
- Walker-Simmons, M.; Ryan, C.A. Immunological identification of proteinase inhibitors I and II in isolated tomato leaf vacuoles. Plant Physiol 1977, 60, 61–63. [Google Scholar]
- Schirra, H.J.; Anderson, M.A.; Craik, D.J. Structural refinement of insecticidal plant proteinase inhibitors from Nicotiana alata. Protein Pept. Lett 2008, 15, 903–909. [Google Scholar]
- Leah, R.; Tommerup, H.; Svendsen, I.; Mundy, J. Biochemical and molecular characterization of three barley seed proteins with antifungal properties. J. Biol. Chem 1991, 246, 1564–1573. [Google Scholar]
- Kamemura, K.; Furuichi, Y.; Umekawa, H.; Takahashi, H.C. Purification and characterization of novel lectins from Great Northern bean, Phaseolus ulgaris L. Biochim. Biophys. Acta 1993, 1158, 181–188. [Google Scholar]
- Pouvreau, L.; Gruppen, H.; van Koningsveld, G.A.; van den Broek, L.A.M.; Voragen, A.G.J. The most abundant protease inhibitor in potato tuber (cv. Elkana) is a protease inhibitor from the Kunitz family. J. Agric. Food Chem 2003, 51, 5001–5005. [Google Scholar]
- Valueva, T.A.; Revina, T.A.; Mosolov, V.V.; Mentele, R. Primary structure of potato Kunitz-type serine proteinase inhibitor. Biol. Chem 2000, 381, 1215–1221. [Google Scholar]
- Sagaram, U.S.; Pandurangi, R.; Kaur, J.; Smith, T.J.; Shah, D.M. Structure-activity determinants in antifungal plant defensins MsDef1 and MtDef4 with different modes of action againstFusarium graminearum. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Mazumdar, S.; Leighton, S.M.; Babu, C.R. Kunitz proteinase inhibitor from Archidendron ellipticum seeds: Purification, characterization, and kinetic properties. Phytochemistry 2006, 67, 232–241. [Google Scholar]
- Kim, J.Y.; Park, S.-C.; Hwang, I.; Cheong, H.; Nah, J.-W.; Hahm, K.-S.; Park, Y. Protease Inhibitors from Plants with Antimicrobial Activity. Int. J. Mol. Sci 2009, 10, 2860–2872. [Google Scholar]
- Herzog, M.; Dorne, A.M.; Grellet, F. GASA, a gibberellin-regulated gene family from Arabidopsis thaliana related to the tomato GAST1 gene. Plant Mol. Biol 1995, 27, 743–752. [Google Scholar]
- Park, C.J.; Park, C.B.; Hong, S.-S.; Lee, H.-S.; Lee, S.Y.; Kim, C. Characterization and cDNA cloning of two glycine- and histidine-rich antimicrobial peptides from the roots of shepherd’s purse, Capsella bursa-pastoris. Plant Mol. Biol 2000, 44, 187–197. [Google Scholar]
- Tam, J.P.; Lu, Y.-A.; Yang, J.-L.; Chiu, K.-W. Anunusual structural motif antimicrobial peptides containing end-to-end macrocycle and cystein-knot disulfides. Proc. Natl. Acad. Sci. USA 1999, 96, 8913–8918. [Google Scholar]
- Benchekroun, A.; Alami, M.; Ghadouan, M.; Lachkar, A.; Kasmaoui, H.; Marzouk, M.; Faik, M. Urinary candidiasis revealed by ureteral obstruction: Report of 2 cases. Ann. Urol (Paris) 2000, 34, 171–174. [Google Scholar]
- Hoy, J.; Hsu, K.C.; Rolston, K.; Hopfer, R.L.; Luna, M.; Bodey, G.P. Trichosporon beigelii infection: A review. Rev. Infect. Dis 1986, 8, 959–967. [Google Scholar]
- Moreno, M.; Segura, A.; García-Olmedo, F. Pseudothionin-St1, a potato peptide active against potato pathogens. Eur. J. Biochem 1994, 223, 135–139. [Google Scholar] [Green Version]
- Laemmli, U.K.; Favre, M. Maturation of the head of bacteriophage T4. J. Mol. Biol 1973, 80, 575–579. [Google Scholar]
- Lee, D.G.; Shin, S.Y.; Maeng, C.-Y.; Jin, Z.Z.; Kim, K.L.; Hahm, K.-S. Isolation and Characterization of a Novel Antifungal Peptide from Aspergillus niger. Biochem. Biophys. Res. Commun 1999, 263, 646–651. [Google Scholar]
- Jahn, B.; Martin, E.; Stueben, A.; Bhakdi, S. Susceptibility testing of Candida albicans and Aspergillus species by a simple microtiter menadione-augmented 3-(4,5-dimethyl-2-thiazolyl)- 2,5-diphenyl-2H-tetrazolium bromide assay. J. Clin. Microbiol 1995, 33, 661–667. [Google Scholar]
- María, E.; Paloma, M.; María, Y.; Mireia, M.; Salvador, V.; Jose, F.M. Selectivity and antimicrobial action of bovine lactoferrin derived peptides against wine lactic acid bacteria. Food Microbiol 2009, 26, 340–346. [Google Scholar]
- Moleyar, V.; Narasimham, P. Antifungal activity of some essential oil components. Food Microbiol 1986, 3, 331–336. [Google Scholar]
- Pouvreau, L.; Gruppen, H.; Piersma, S.R.; van den Broek, L.A.M.; van Koningsveld, G.A.; Voragen, A.G.J. Relative abundance and inhibitory distribution of protease inhibitors in potato juice from cv. Elkana. J. Agric. Food Chem 2001, 49, 2864–2874. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Yield (mg) |
|---|---|
| Potato tuber | 200,000 |
| Protein Extraction Buffer | 85.42 |
| Gel filtration | 15.3 |
| FPLC | 7.86 |
| First C18-HPLC | 1.56 |
| Second C18-HPLC | 0.05 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, J.-K.; Gopal, R.; Seo, C.H.; Cheong, H.; Park, Y. Isolation and Purification of a Novel Deca-Antifungal Peptide from Potato (Solanum tuberosum L. cv. Jopung) Against Candida albicans. Int. J. Mol. Sci. 2012, 13, 4021-4032. https://doi.org/10.3390/ijms13044021
Lee J-K, Gopal R, Seo CH, Cheong H, Park Y. Isolation and Purification of a Novel Deca-Antifungal Peptide from Potato (Solanum tuberosum L. cv. Jopung) Against Candida albicans. International Journal of Molecular Sciences. 2012; 13(4):4021-4032. https://doi.org/10.3390/ijms13044021
Chicago/Turabian StyleLee, Jong-Kook, Ramamourthy Gopal, Chang Ho Seo, Hyeonsook Cheong, and Yoonkyung Park. 2012. "Isolation and Purification of a Novel Deca-Antifungal Peptide from Potato (Solanum tuberosum L. cv. Jopung) Against Candida albicans" International Journal of Molecular Sciences 13, no. 4: 4021-4032. https://doi.org/10.3390/ijms13044021