Beta-Glucans Improve Growth, Viability and Colonization of Probiotic Microorganisms

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
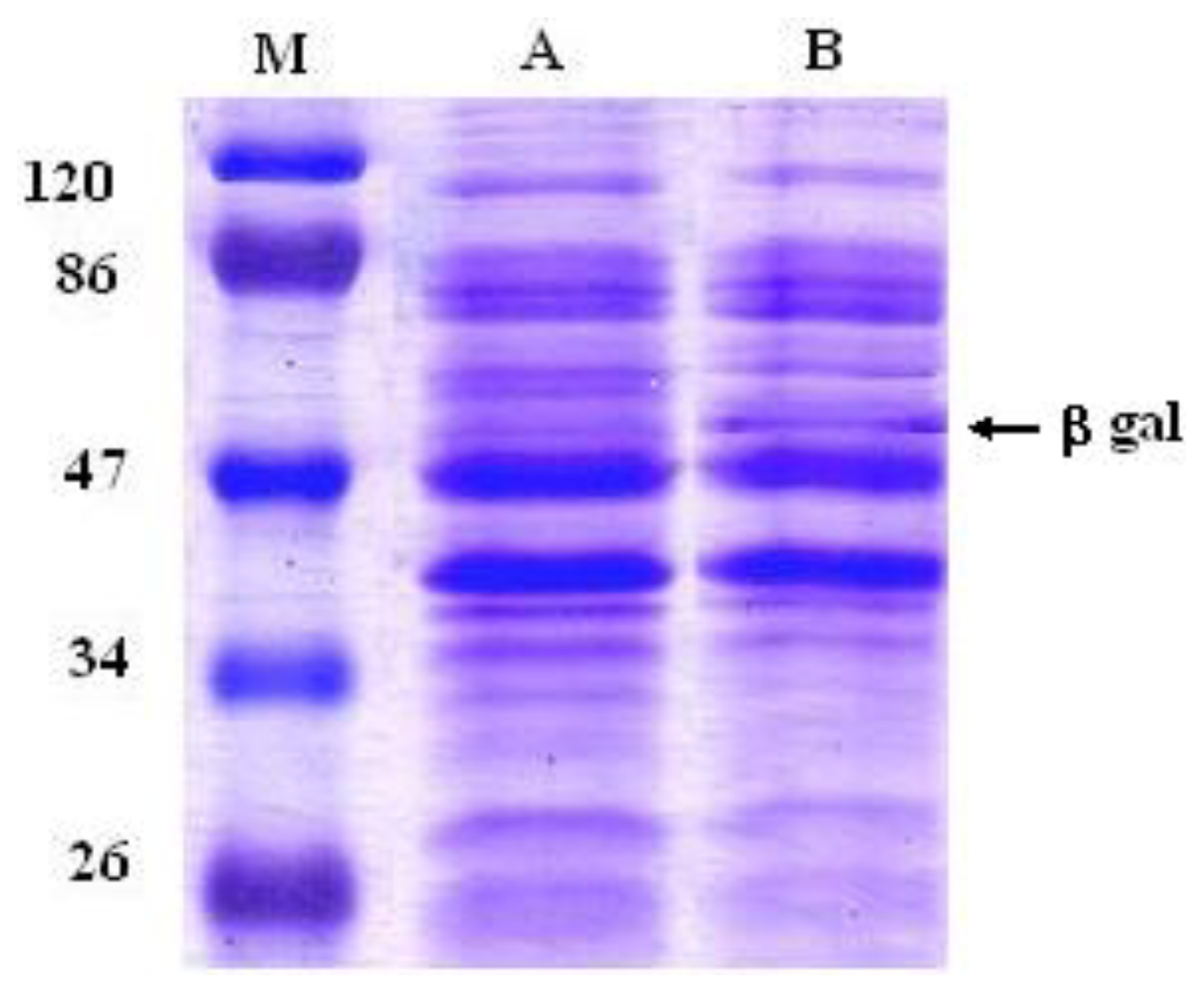
2.1. Overexpression of a β-Glycosidase Gene in L. plantarum WCFS1
2.2. Prebiotic Characterization of EPS
3. Experimental Section
3.1. Bacterial Strains and Media
3.2. Production of EPS by P. parvulus 2.6
3.3. Overexpression of Bgl Gene in L. plantarum
3.4. Protein Extraction and SDS-PAGE
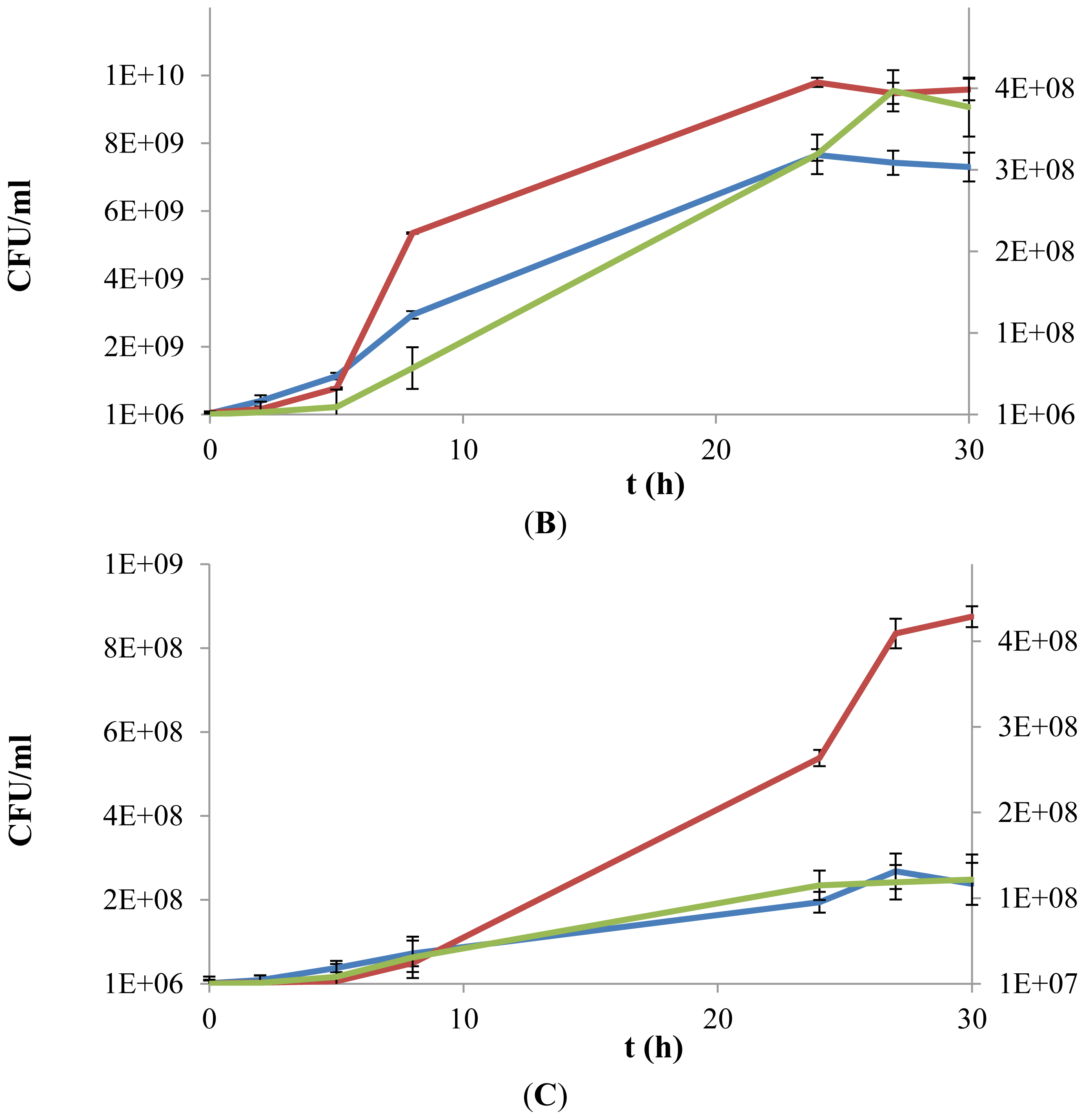
3.5. Kinetics of Cell Growth with Different Carbon Sources
3.6. Adhesion Assay
3.7. Statiscal Analysis
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Reid, G.; Sanders, M.E.; Gaskins, H.R.; Gibson, G.R.; Mercenier, A.; Rastall, R.; Roberfroid, M.; Rowland, I.; Cherbut, C.; Klaenhammer, T.R. New scientific paradigms for probiotics and prebiotics. J. Clin. Gastroenterol 2003, 37, 105–118. [Google Scholar]
- De Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, and synbiotics. Adv. Biochem. Eng. Biotechnol 2008, 111, 1–66. [Google Scholar]
- Cummings, J.H.; Macfarlane, G.T.; Englyst, H.N. Prebiotic digestion and fermentation. Am. J. Clin. Nutr 2001, 73, 415–420. [Google Scholar]
- Marteau, P.R.; de Vrese, M.; Cellier, C.J.; Schrezenmeir, J. Protection from gastrointestinal diseases with the use of probiotics. Am. J. Clin. Nutr 2001, 73, 430–436. [Google Scholar]
- Gibson, G.R.; Probert, H.M.; van loo, J.; Rastall, R.A.; Roberfroid, M. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev 2004, 17, 259–275. [Google Scholar]
- Sutherland, I.W. Microbial polysaccharides from Gram-negative bacteria. Int. Dairy J 2001, 11, 663–674. [Google Scholar]
- Ruas-Madiedo, P.; Moreno, J.A.; Salazar, N.; Delgado, S.; Mayo, B.; Margolles, A.; de los Reyes-Gavilán, C.G. Screening of exopolysaccharide-producing lactobacillus and bifidobacterium strains isolated from the human intestinal microbiota. Appl. Environ. Microbiol 2007, 73, 4385–4388. [Google Scholar]
- Liu, C.-F.; Tseng, K.-C.; Chiang, S.S.; Lee, B.H.; Hsu, W.-H.; Pan, T.-M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric 2011, 91, 2284–2291. [Google Scholar]
- Dueñas-Chasco, M.T.; Rodríguez-Carvajal, M.A.; Mateo, P.T.; Franco-Rodríguez, G.; Espartero, J.; Irastorza-Iribas, A.; Gil-Serrano, A.M. Structural analysis of the exopolysaccharide produced by Pediococcus damnosus 2.6. Carbohydr. Res 1997, 303, 453–458. [Google Scholar]
- Velasco, S.E.; Areizaga, J.; Irastorza, A.; Dueñas, M.T.; Santamaría, A.; Muñoz, M.E. Chemical and rheological properties of the β-glucan produced by Pediococcus parvulus 2.6. J. Agric. Food Chem 2009, 57, 1827–1834. [Google Scholar]
- Zekovic, D.B.; Kwiatkowski, S.; Vrvic, M.M.; Jakovljevic, D.; Moran, C.A. Natural and modified (1→3)-β-d-glucans in health promotion and disease alleviation. Crit. Rev. Biotechnol 2005, 25, 205–230. [Google Scholar]
- Sletmoen, M.; Stokke, B.T. Higher order structure of (1,3)-β-d-glucans and its influence on their biological activities and complexation abilities. Biopolymers 2008, 89, 310–321. [Google Scholar]
- Fernández de Palencia, P.; Werning, M.L.; Sierra-Filardi, E.; Dueñas, M.T.; Irastorza, A.; Corbí, A.L.; López, P. Probiotic properties of the 2-substituted (1,3)-β-d-glucan-producing bacterium Pediococcus parvulus 2.6. Appl. Environ. Microbiol 2009, 75, 4887–4891. [Google Scholar]
- Garai-Ibabe, G.; Dueñas, M.T.; Irastorza, A.; Sierra-Filardi, E.; Werning, M.L.; López, P.; Corbi, A.L.; Fernández de Palencia, P. Naturally occurring 2-substituted (1,3)-β-d-glucan producing Lactobacillus suebicus and Pediococcus parvulus strains with potential utility in the production of functional foods. Biores. Technol 2010, 101, 9254–9263. [Google Scholar]
- Stack, H.M.; Kearney, N.; Stanton, C.; Gerald, F.; Fitzgerald, G.F.; Ross, R.P. Association of β-glucan endogenous production with increased stress tolerance of intestinal lactobacilli. Appl. Environ. Microbiol 2010, 76, 500–507. [Google Scholar]
- Kearney, N.; Stack, H.M.; Tobin, J.; Chaurin, V.; Fenelon, M.A.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Lactobacillus paracasei NFBC 338 producing recombinant beta-glucan positively influences the functional properties of yoghurt. Int. Dairy J 2011, 21, 561–567. [Google Scholar]
- Mårtensson, O.; Biörklund, M.; Lambo, M.A.; Dueñas-Chasco, M.T.; Irastorza, A.; Holst, O.; Norin, E.; Walling, G.; Öste, R.; Önning, G. Fermented ropy, oat based products reduce cholesterol levels and stimulate the bifidobacteria flora in humans. Nutr. Res 2005, 25, 429–442. [Google Scholar]
- Korakli, M.; Gänzle, M.G.; Vogel, R.F. Metabolism by bifidobacteria and lactic acid bacteria of polysaccharides from wheat and rye, and exopolysaccharides produced by Lactobacillus sanfranciscensis. J. Appl. Microbiol 2002, 92, 958–965. [Google Scholar]
- Manning, T.S.; Gibson, G.R. Prebiotics. Best Pract. Res. Clin. Gastroenterol 2004, 18, 287–298. [Google Scholar]
- Spano, G.; Rinaldi, A.; Ugliano, M.; Moio, L.; Beneduce, L.; Massa, S. A β-glucosidase gene isolated from wine Lactobacillus plantarum is regulated by abiotic stresses. J. Appl. Microbiol 2005, 98, 855–861. [Google Scholar]
- Acebrón, I.; Curiel, J.A.; de las Rivas, B.; Muñoz, R.; Mancheño, J.M. Cloning, production, purification and preliminary crystallographic analysis of a glycosidase from the food lactic acid bacterium Lactobacillus plantarum CECT 748T. Prot. Expr. Purif 2009, 68, 177–182. [Google Scholar]
- Ketudat-Cairns, J.R.; Esen, A. β-glucosidases. Cell. Mol. Life Sci 2010, 67, 3389–3405. [Google Scholar]
- Molin, G. Probiotics in foods not containing milk or milk constituents, with special reference to Lactobacillus plantarum 299v. Am. J. Clin. Nutr 2001, 73, 380–385. [Google Scholar]
- Altermann, E.; Russell, W.M.; Azcarate-Peril, M.A.; Barrangou, R.; Buck, B.L.; McAuliffe, O.; Souther, N.; Dobson, A.; Duong, T.; Callanan, M.; et al. Complete genome sequence of the probiotic lactic acid bacterium Lactobacillus acidophilus NCFM. Proc. Natl. Acad. Sci. USA 2005, 102, 3906–3912. [Google Scholar]
- Terrade, N.; Noël, R.; Couillaud, R.; Mira de Orduña, R. A new chemically defined medium for wine lactic acid bacteria. Food Res. Int 2009, 42, 363–367. [Google Scholar]
- Terrade, N.; Mira de Orduña, R. Determination of the essential nutrient requirements of wine-related bacteria from the genera Oenococcus and Lactobacillus. Int. J. Food Microbiol 2009, 133, 8–13. [Google Scholar]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.; et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar]
- Katayama, T.; Fujita, K.; Yamamoto, K. Novel bifidobacterial glycosidases acting on sugar chains of mucin glycoproteins. J. Biosci. Bioeng 2005, 99, 457–465. [Google Scholar]
- Schell, M.A.; Karmirantzou, M.; Snel, B.; Vilanova, D.; Berger, B.; Pessi, G.; Zwahlen, M.C.; Desiere, F.; Bork, P.; Delley, M.; et al. The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. USA 2002, 99, 14422–14427. [Google Scholar]
- Dols-Lafargue, M.; Lee, H.Y.; le marrec, C.; Heyraud, A.; Chambat, G.; Lonvaud-Funel, A. Characterization of gtf, a glucosyltransferase gene in the genomes of Pediococcus parvulus and Oenococcus oeni. Appl. Environ. Microbiol 2008, 74, 4079–4090. [Google Scholar]
- Fernández de Palencia, P.; López, P.; Corbí, A.L.; Peláez, C.; Requena, T. Probiotic strains: Survival under simulated gastrointestinal conditions, in vitro adhesion to Caco-2 cells and effect on cytokine secretion. Eur. Food Res. Technol 2008, 227, 1475–1484. [Google Scholar]
- Van den Abbeele, P.; van de Wiele, T.; Grootaert, C.; Verstraete, W.; Gérard, P.; Bruneau, A.; Rabot, S.; Possemiers, S. Arabinoxylans and Inulin Modulate the Luminal and Mucosa-Associated Bacteria in Vitro and in Vivo. In Dietary Fibre: New Frontiers for Food and Health; van der Kamp, J.W., Jones, J.M., McCleary, B.V., Topping, D.L., Eds.; Wageningen Academic Publishers: Waltham, MA, USA, 2010; pp. 233–249. [Google Scholar]
- Brink, M.; Todorov, S.D.; Martin, J.H.; Senekal, M.; Dicks, L.M. The effect of prebiotics on production of antimicrobial compounds, resistance to growth at low pH and in the presence of bile, and adhesion of probiotic cells to intestinal mucus. J. Appl. Microbiol 2006, 100, 813–820. [Google Scholar]
- Rhoades, J.; Gibson, G.R.; Formentin, K.; Beer, M.; Rastall, R.A. Inhibition of the adhesion of enteropathogenic Escherichia coli strains to HT-29 Cells in Culture by Chito-oligosaccharides. Carbohydr. Polym 2006, 64, 57–59. [Google Scholar]
- Shoaf, K.; Mulvey, G.L.; Armstrong, G.D.; Hutkins, R.W. Prebiotic galactooligosaccharides reduce adherence of enteropathogenic Escherichia coli to tissue culture cells. Infect. Immun 2006, 74, 6920–6928. [Google Scholar]
- Isolauri, E.; Salminen, S.; Ouwehand, A.C. Probiotics. Best Pract. Res. Clin. Gastroenterol 2004, 18, 299–313. [Google Scholar]
- Van den Ende, W.; Peshev, D.; de Gara, L. Disease prevention by natural antioxidants and prebiotics acting as ROS scavengers in the gastrointestinal tract. Trends Food Sci. Technol 2011, 22, 689–697. [Google Scholar]
- Capozzi, V.; Arena, M.P.; Crisetti, E.; Spano, G.; Fiocco, D. The hsp 16 gene of the probiotic Lactobacillus acidophilus is differently regulated by salt, high temperature and acidic stresses as revealed by reverse transcription quantitative PCR (qRT-PCR) analysis. Int. J. Mol. Sci 2011, 12, 5390–5405. [Google Scholar]
- Velasco, S.; Årskold, E.; Paese, M.; Grage, A.; Irastorza, A.; Rådstrom, P.; van Niel, E.W.J. Environmental factors influencing growth of and exopolysaccharide formation by Pediococcus parvulus 2.6. Int. J. Food Microbiol 2006, 111, 252–258. [Google Scholar]
- Bron, P.A.; Sally, M.; Hoffer, I.; van Swam, I.; de Vos, W.M.; Kleerebezem, M. Selection and characterization of conditionally active promoters in Lactobacillus plantarum, using alanine racemase as a promoter probe. Appl. Environ. Microbiol 2004, 70, 310–317. [Google Scholar]
- Bove, P.; Gallone, A.; Russo, P.; Capozzi, V.; Albenzio, M.; Spano, G.; Fiocco, D. Probiotic features of Lactobacillus plantarum mutant strains. Appl. Microbiol. Biotechnol 2012. [Google Scholar] [CrossRef]





© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Russo, P.; López, P.; Capozzi, V.; De Palencia, P.F.; Dueñas, M.T.; Spano, G.; Fiocco, D. Beta-Glucans Improve Growth, Viability and Colonization of Probiotic Microorganisms. Int. J. Mol. Sci. 2012, 13, 6026-6039. https://doi.org/10.3390/ijms13056026
Russo P, López P, Capozzi V, De Palencia PF, Dueñas MT, Spano G, Fiocco D. Beta-Glucans Improve Growth, Viability and Colonization of Probiotic Microorganisms. International Journal of Molecular Sciences. 2012; 13(5):6026-6039. https://doi.org/10.3390/ijms13056026
Chicago/Turabian StyleRusso, Pasquale, Paloma López, Vittorio Capozzi, Pilar Fernández De Palencia, María Teresa Dueñas, Giuseppe Spano, and Daniela Fiocco. 2012. "Beta-Glucans Improve Growth, Viability and Colonization of Probiotic Microorganisms" International Journal of Molecular Sciences 13, no. 5: 6026-6039. https://doi.org/10.3390/ijms13056026