Anti-Proliferation and Anti-Invasion Effects of Diosgenin on Gastric Cancer BGC-823 Cells with HIF-1α shRNAs

Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of a Hypoxia-Mimetic Chemical Sensitive Gastric Cancer Cell Line
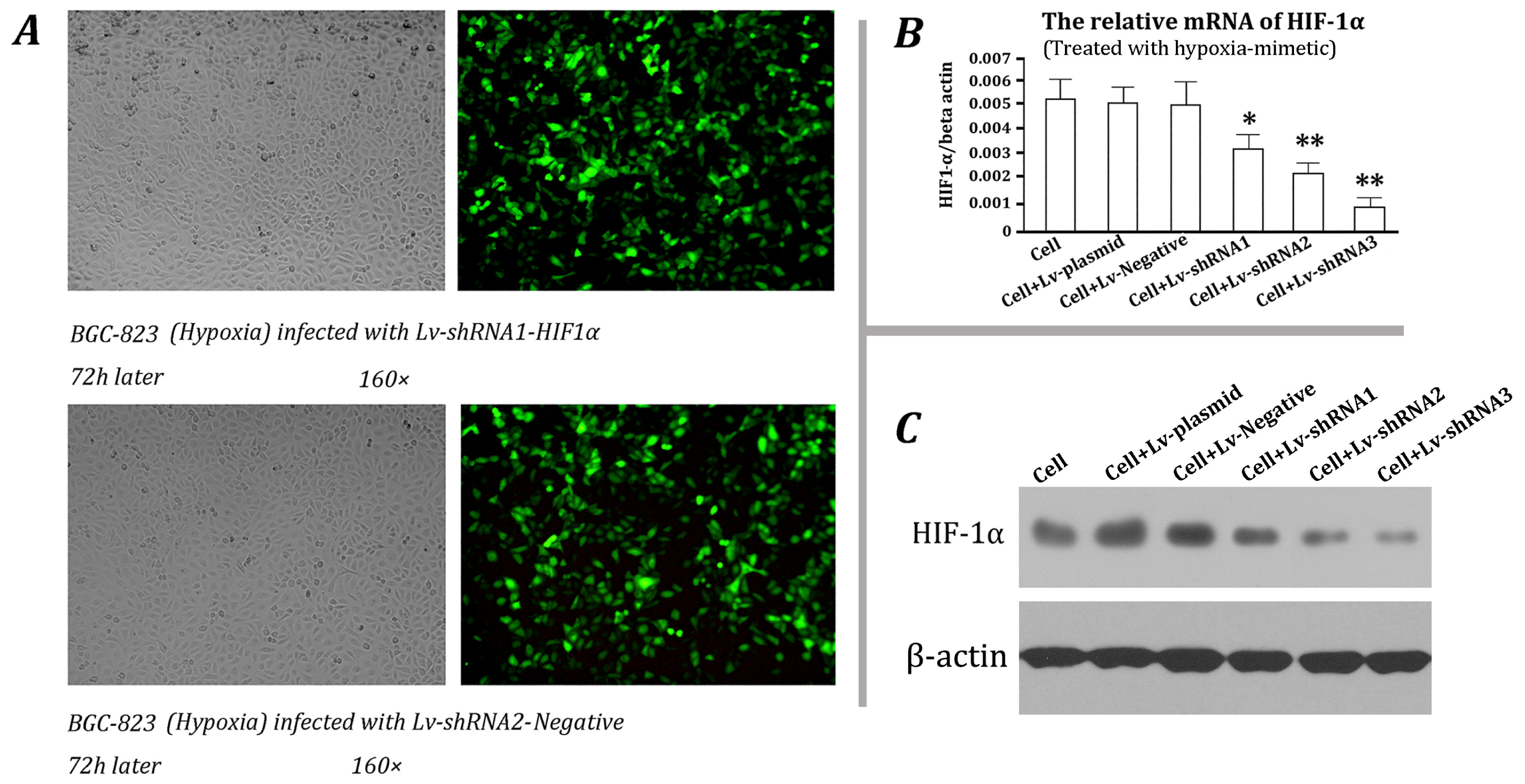
2.2. Screening of Effective shRNA Vector
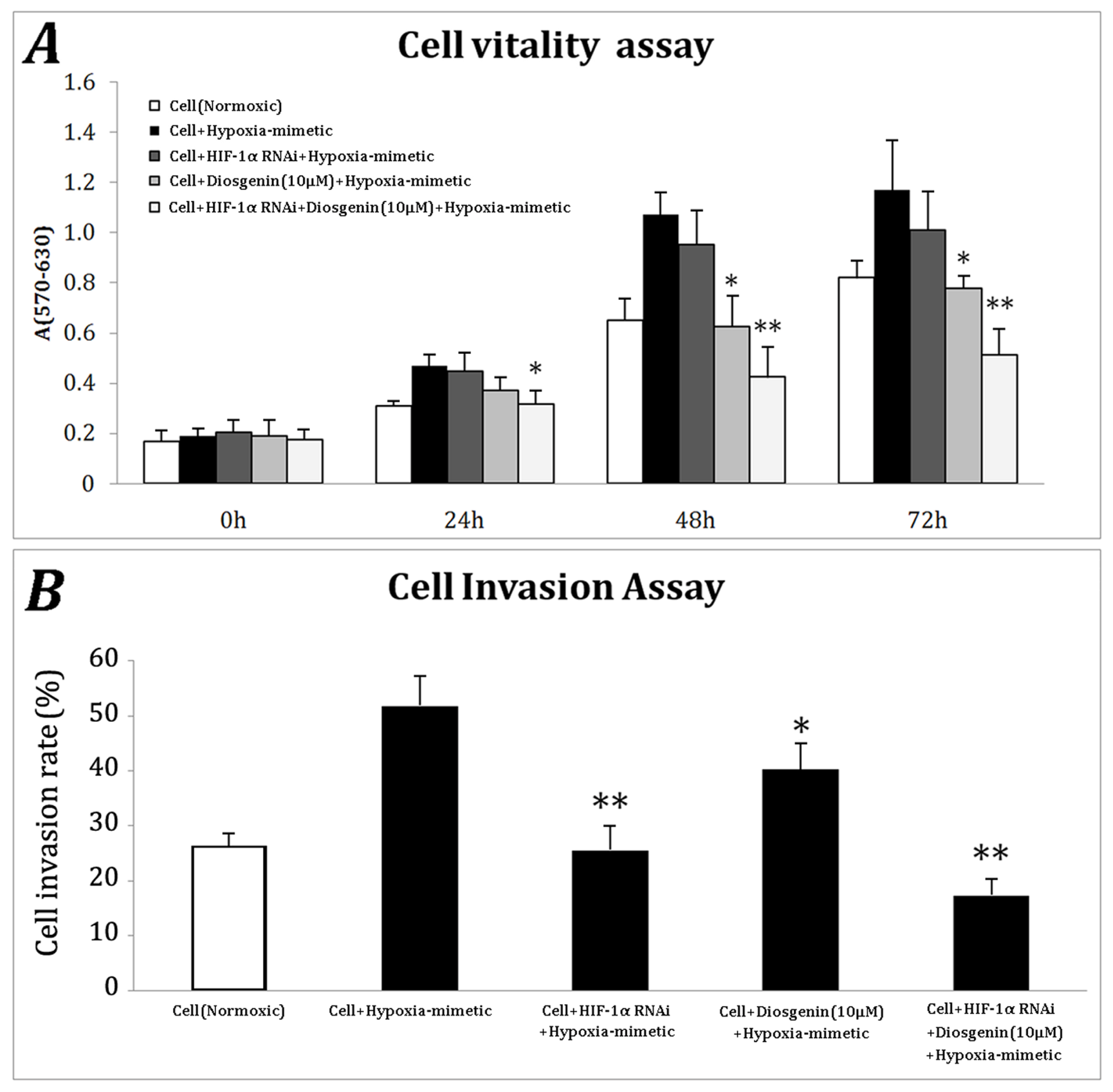
2.3. HIF-1α Knockdown Enhances the Anti-Proliferation and Anti-Invasion Ability of Diosgenin
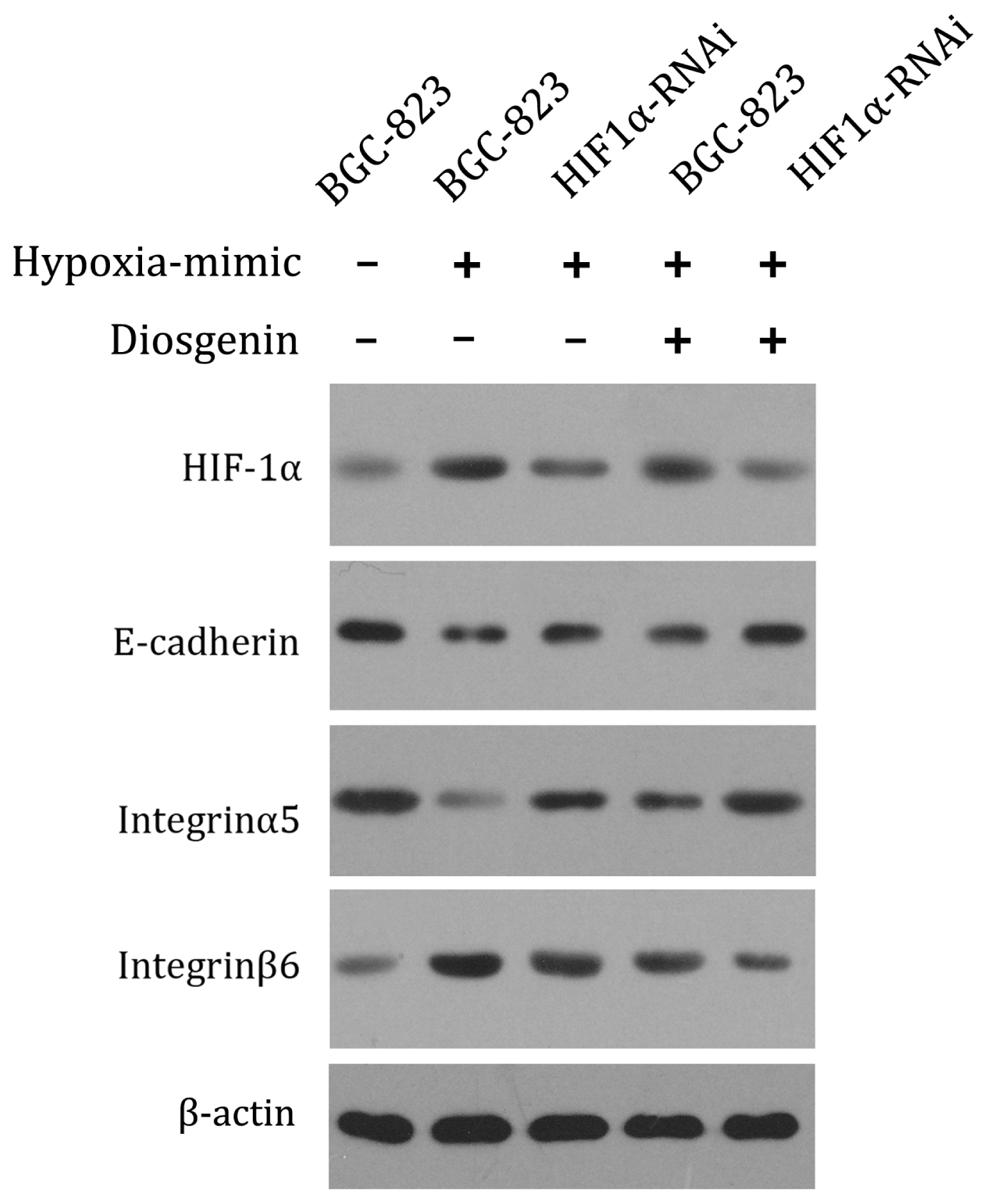
2.4. Anti-Invasion Role of Diosgenin Involves the Alteration of Cell Adhesive Molecules Expression
3. Experimental Section
3.1. Cell Culture and Cobalt Chloride Treatment
3.3. Cell Transduction, Stable Clone Selection, and Cobalt Chloride Treatment
3.4. RNA Extraction and Quantitative RT-PCR
3.5. Western Blotting
3.6. Cell Proliferation
3.7. Cell Invasion Assay
3.8. Statistical Analysis
4. Conclusions
References
- Lin, X.; Zhang, D.; Du, R.; Pan, Y.; Zhao, L.; Sun, S.; Hong, L.; Fan, D. miR-15b and miR-16 modulate multidrug resistance by targeting BCL2 in human gastric cancer cells. Int. J. Cancer 2008, 123, 372–379. [Google Scholar]
- Urano, N.; Fujiwara, Y.; Doki, Y.; Kim, S.J.; Miyoshi, Y.; Noguchi, S.; Miyata, H.; Takiguchi, S.; Yasuda, T.; Yano, M.; et al. Clinical significance of class III β-tubulin expression and its predictive value for resistance to docetaxel-based chemotherapy in gastric cancer. Int. J. Oncol 2006, 28, 375–381. [Google Scholar]
- Dean, M.; Fojo, T.; Bates, S. Tumour stem cells and drug resistance. Nat. Rev. Cancer 2005, 5, 275–284. [Google Scholar]
- Casanovas, O.; Daniel, J.; Bergers, G.; Hanahan, D. Drug resistance by evasion of antiangiogenic targeting of VEGF signaling in late-stage pancreatic islet tumors. Cancer Cell 2005, 8, 299–309. [Google Scholar]
- Siegfried, J.A.; Kennedy, K.A. The role of membranes in the mechanism of action of the antineoplastic agent adriamycin. Spin-labeling studies with chronically hypoxic and drug-resistant tumor cells. J. Biol. Chem 1983, 258, 339–343. [Google Scholar]
- Katrina, M.; Comerford, T.; Wallace, J. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res 2002, 62, 3387–3394. [Google Scholar]
- Xu, R.-H.; Pelicano, H. Inhibition of glycolysis in cancer cells: A novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia. Cancer Res 2005, 65, 613–621. [Google Scholar]
- Huang, L.E.; Arany, Z.; Livingston, D.M.; Bunn, H.F. Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitive stabilization of its α subunit. J. Biol. Chem 1996, 271, 32253–32259. [Google Scholar]
- Krieg, M.; Haas, R.; Brauch, H.; Acker, T.; Flamme, I.; Plate, K.H. Up-regulation of hypoxia-inducible factors HIF-1α and HIF-2α under normoxic conditions in renal carcinoma cells by von Hippel-Lindau tumor suppressor gene loss of function. Oncogene 2000, 19, 5435–5443. [Google Scholar]
- Jones, N.M.; Lee, E.M. Hypoxic preconditioning produces differential expression of hypoxia-inducible factor-1α (HIF-1α) and its regulatory enzyme HIF prolyl hydroxylase 2 in neonatal rat brain. Neurosci. Lett 2006, 404, 72–77. [Google Scholar]
- Talks, K.L.; Turley, H.; Gatter, K.C.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J.; Harris, A.L. The expression and distribution of the hypoxia-inducible factors HIF-1α and HIF-2α in normal human tissues, cancers, and tumor-associated macrophages. Am. J. Pathol 2000, 157, 411–421. [Google Scholar]
- Zhong, H.; de Marzo, A.M. Overexpression of hypoxia-inducible factor 1α in common human cancers and their metastases. Cancer Res 1999, 59, 5830–5835. [Google Scholar]
- Zhou, J.; Schmid, T. Tumor hypoxia and cancer progression. Cancer Lett 2006, 237, 10–21. [Google Scholar]
- Liu, L.; Ning, X. Hypoxia-inducible factor-1 contributes to hypoxia-induced chemoresistance in gastric cancer. Cancer Sci 2008, 99, 121–128. [Google Scholar]
- Giaccia, A.; Siim, B.G.; Johnson, R.S. HIF-1 as a target for drug development. Nat. Rev. Drug Discov 2003, 2, 803–811. [Google Scholar]
- Sautour, M.; Mitaine-Offer, A.C.; Miyamoto, T.; Dongmo, A.; Lacaille-Dubois, M.A. A new steroidal saponin from Dioscorea cayenensis. Chem. Pharm. Bull. (Tokyo) 2004, 52, 1353–1355. [Google Scholar]
- Daisy, P.; Eliza, J. Influence of Costus speciosus (Koen.) Sm. Rhizome extracts on biochemical parameters in streptozotocin induced diabetic rats. J. Health Sci 2008, 54, 675–681. [Google Scholar]
- Raju, J.; Bird, R.P. Diosgenin, a naturally occurring steroid [corrected] saponin suppresses 3-hydroxy-3-methylglutaryl CoA reductase expression and induces apoptosis in HCT-116 human colon carcinoma cells. Cancer Lett 2007, 55, 194–204. [Google Scholar]
- Jeon, J.R.; Lee, J.S.; Lee, C.H.; Kim, J.Y.; Kim, S.D.; Nam, D.H. Effect of ethanol extract of dried Chinese yam (Dioscorea batatas) flour containing dioscin on gastrointestinal function in rat model. Arch. Pharm. Res 2006, 29, 348–353. [Google Scholar]
- Son, I.S.; Kim, J.H.; Shon, H.Y.; Son, K.H.; Kim, J.S.; Kwon, C.S. Antioxidative and hypolipidemic effects of diosgenin, a steroidal saponin of yam (Dioscorea spp.), on high-cholesterol fed rats. Biosci. Biotechnol. Biochem 2007, 71, 3063–3071. [Google Scholar]
- Lee, J.; Jung, K.; Kim, Y.S. Diosgenin inhibits melanogenesis through the activation of hosphatidylinositol-3-kinase pathway (PI3K) signaling. Life Sci 2007, 81, 249–254. [Google Scholar]
- Kang, W.H.; Yoon, K.H.; Lee, E.S.; Kim, J.; Lee, K.B.; Yim, H.; Sohn, S.; Im, S. Histopathalogical characteristics in 56 Korean patients. Br. J. Dermatol 2002, 146, 228–237. [Google Scholar]
- Li, J.; Liu, X.; Guo, M.; Liu, Y.; Liu, S.; Liu, S.; Yao, S. Electrochemical study of breast cancer cells MCF-7 and its application in evaluating the effect of diosgenin. Anal. Sci 2005, 21, 561–564. [Google Scholar]
- Raju, J.; Bird, R.P. Diosgenin, a naturally occurring furostanol saponin suppresses 3-hydroxy-3-m ethylglutaryl CoA reductase expression and induces apoptosis in HCT-116 human colon carcinoma cells. Cancer Lett 2007, 255, 194–204. [Google Scholar]
- Moalic, S.; Liagre, B.; Corbiere, C.; Bianchi, A.; Dauca, M.; Bordji, K.; Beneytout, J.L. A plant steroid, diosgenin, induces apoptosis, cell cycle arrest and COX activity in osteosarcoma cells. FEBS Lett 2001, 506, 225–230. [Google Scholar]
- Rohwer, N.; Welzel, M.; Daskalow, K.; Pfander, D.; Wiedenmann, B.; Detjen, K.; Cramer, T. Hypoxia-inducible factor 1α mediates anoikis resistance via suppression of α5 integrin. Cancer Res 2008, 68, 10113–10118. [Google Scholar]
- Liu, L.; Ning, X.X. Hypoxia-inducible factor-1 contributes to hypoxia-induced chemoresistance in gastric cancer. Cancer Sci 2008, 99, 121–128. [Google Scholar]
- Greco, O.; Marpels, B.; Joiner, M.C.; Scott, S.D. How to overcome (and exploit) tumor hypoxia for targeted gene therapy. J. Cell Physiol 2003, 197, 312–325. [Google Scholar]
- Durand, R.E. Keynote address: The influence of microenvironmental factors on the activity of radiation and drugs. Int. J. Radiat. Oncol. Biol. Phys 1991, 20, 253–258. [Google Scholar]
- Hockel, M.; Vaupel, P. Tumor hypoxia: Definitions and current clinical, biologic, and molecular aspects. J. Natl. Cancer Inst 2001, 93, 266–276. [Google Scholar]
- Greijer, A.E.; de Jong, M.C.; Scheffer, G.L.; Shvarts, A.; van Diest, P.J.; van der wall, E. Hypoxia-induced acidification causes mitoxantrone resistance not mediated by drug transporters in human breast cancer cells. Cell. Oncol 2005, 27, 43–49. [Google Scholar]
- Matthews, N.E.; Adams, M.A.; Maxwell, L.R.; Gofton, T.E.; Graham, C.H. Nitric oxide-mediated regulation of chemosensitivity in cancer cells. J. Natl. Cancer Inst 2001, 93, 1879–1885. [Google Scholar]
- Flamant, L.; Notte, A.; Ninane, N.; Raes, M.; Michiels, C. Anti-apoptotic role of HIF-1 and AP-1 in paclitaxel exposed breast cancer cells under hypoxia. Mol. Cancer 2010, 9. [Google Scholar] [CrossRef]
- Griffiths, E.A.; Pritchard, S.A.; Welch, I.M.; Price, P.M.; West, C.M. Is the hypoxia-inducible factor pathway important in gastric cancer? Eur. J. Cancer 2005, 41, 2792–2805. [Google Scholar]
- Takeshita, F.; Ochiya, T. Therapeutic potential of RNA interference against cancer. Cancer Sci 2006, 97, 689–696. [Google Scholar]
- Dame, C.; Haugstetter, A.; Rohwer, N.; Wiedenmann, B.; Detjen, K.; Schmitt, C.A.; Cramer, T. Hypoxia-inducible factor 1α determines gastric cancer chemosensitivity via modulation of p53 and NF-kB. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Murthy, N.S.; Mukherjee, S.; Ray, G.; Ray, A. Dietary factors and cancer chemoprevention: An overview of obesity-related malignancies. J. Postgrad. Med 2009, 55, 45–54. [Google Scholar]
- Berx, G.; Cleton-Jansen, A.M.; Nollet, F.; de Leeuw, W.J.; van de Vijver, M.; Cornelisse, C.; van Roy, F. E-cadherin is a tumour/invasion suppressor gene mutated in human lobular breast cancers. EMBO J 1995, 14, 6107–6115. [Google Scholar]
- Wong, A.S.; Gumbiner, B.M. Adhesion-independent mechanism for suppression of tumor cell invasion by E-cadherin. J. Cell Biol 2003, 161, 1191–1203. [Google Scholar] [Green Version]
- Zutter, M.M.; Mazoujian, G.; Santoro, S.A. Decreased expression of integrin adhesive protein receptors in adenocarcinoma of the breast. Am. J. Pathol 1990, 137, 863–870. [Google Scholar]
- Ramos, D.M.; But, M.; Regezi, J.; Schmidt, B.L.; Atakilit, A.; Dang, D.; Ellis, D.; Jordan, R.; Li, X. Expression of integrin beta 6 enhances invasive behavior in oral squamous cell carcinoma. Matrix Biol 2002, 21, 297–307. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| shRNA | Target sequence (5′–3′) | Oligonucleotide sequence(5′–3′) | |
|---|---|---|---|
| shRNA 1 | GACTTTCCTCAGTCGACAC | Forward | GATCCGACTTTCCTCAGTCGACACCTTCCTGTCAGAGTGTCGACTGAGGAAAGTCTTTTTG |
| Reverse | AATTCAAAAAGACTTTCCTCAGTCGACACTCTGACAGGAAGGTGTCGACTGAGGAAAGTCG | ||
| shRNA 2 | GTCACCACAGGACAGTACA | Forward | GATCCGTCACCACAGGACAGTACACTTCCTGTCAGATGTACTGTCCTGTGGTGACTTTTTG |
| Reverse | AATTCAAAAAGTCACCACAGGACAGTACATCTGACAGGAAGTGTACTGTCCTGTGGTGACG | ||
| shRNA 3 | GTAGTGCTGACCCTGCACT | Forward | GATCCGTAGTGCTGACCCTGCACTCTTCCTGTCAGAAGTGCAGGGTCAGCACTACTTTTTG |
| Reverse | AATTCAAAAAGTAGTGCTGACCCTGCACTTCTGACAGGAAGAGTGCAGGGTCAGCACTACG | ||
| shRNA-negative | GAAGCCAGATCCAGCTTCC | Forward | GATCCGAAGCCAGATCCAGCTTCCCTTCCTGTCAGAGGAAGCTGGATCTGGCTTCTTTTTG |
| Reverse | AATTCAAAAAGAAGCCAGATCCAGCTTCCTCTGACAGGAAGGGAAGCTGGATCTGGCTTCG | ||
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mao, Z.-J.; Tang, Q.-J.; Zhang, C.-A.; Qin, Z.-F.; Pang, B.; Wei, P.-k.; Liu, B.; Chou, Y.-N. Anti-Proliferation and Anti-Invasion Effects of Diosgenin on Gastric Cancer BGC-823 Cells with HIF-1α shRNAs. Int. J. Mol. Sci. 2012, 13, 6521-6533. https://doi.org/10.3390/ijms13056521
Mao Z-J, Tang Q-J, Zhang C-A, Qin Z-F, Pang B, Wei P-k, Liu B, Chou Y-N. Anti-Proliferation and Anti-Invasion Effects of Diosgenin on Gastric Cancer BGC-823 Cells with HIF-1α shRNAs. International Journal of Molecular Sciences. 2012; 13(5):6521-6533. https://doi.org/10.3390/ijms13056521
Chicago/Turabian StyleMao, Zhu-Jun, Qian-Jue Tang, Ci-An Zhang, Zhi-Feng Qin, Bin Pang, Pin-kang Wei, Bo Liu, and Yuan-Neng Chou. 2012. "Anti-Proliferation and Anti-Invasion Effects of Diosgenin on Gastric Cancer BGC-823 Cells with HIF-1α shRNAs" International Journal of Molecular Sciences 13, no. 5: 6521-6533. https://doi.org/10.3390/ijms13056521