Opuntia humifusa Supplementation Increased Bone Density by Regulating Parathyroid Hormone and Osteocalcin in Male Growing Rats

Abstract
:1. Introduction
2. Results
2.1. Body Weight, FER and Food Intake
2.2. Changes in Serum Parameters
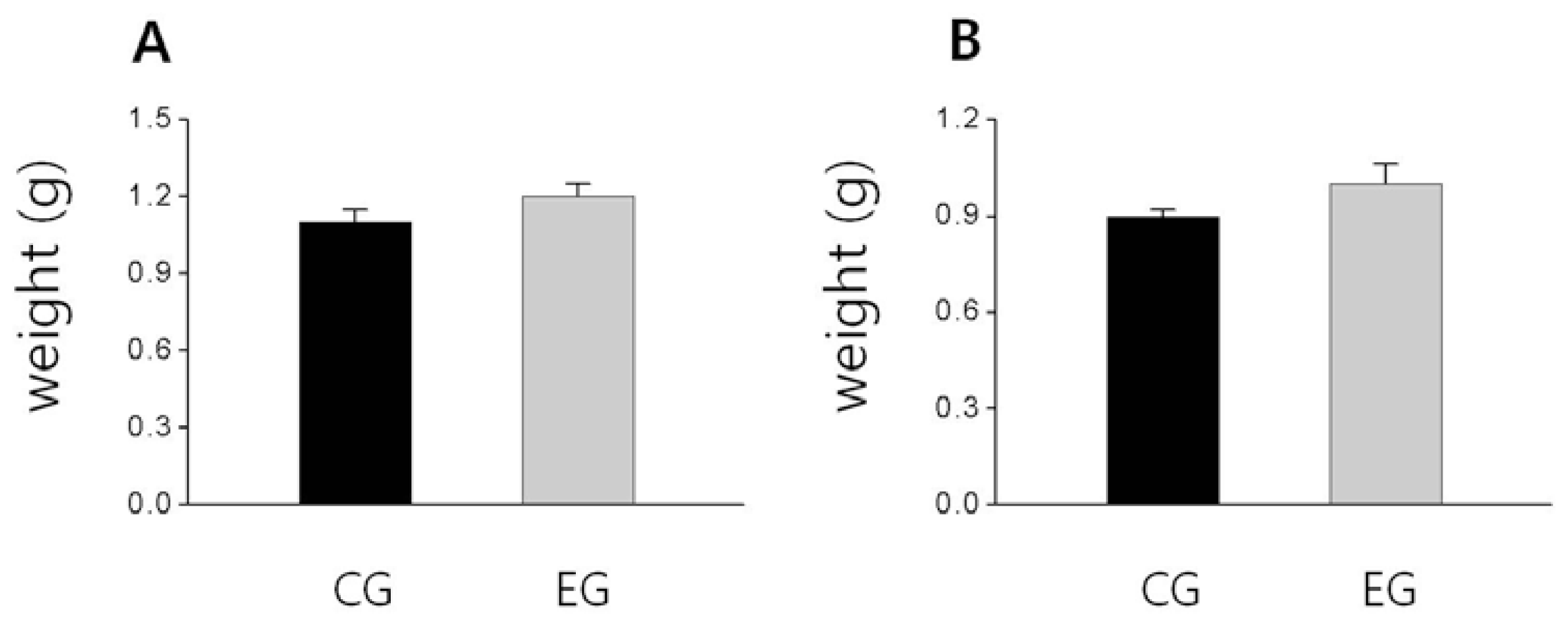
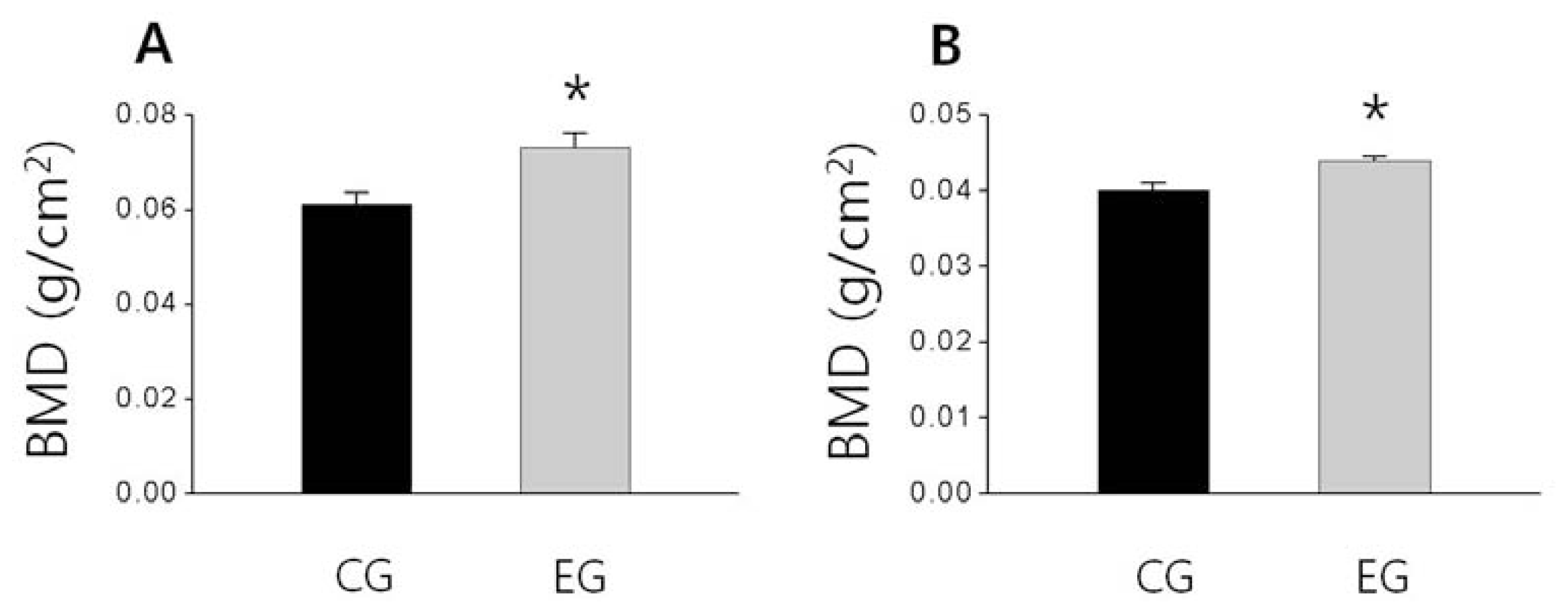
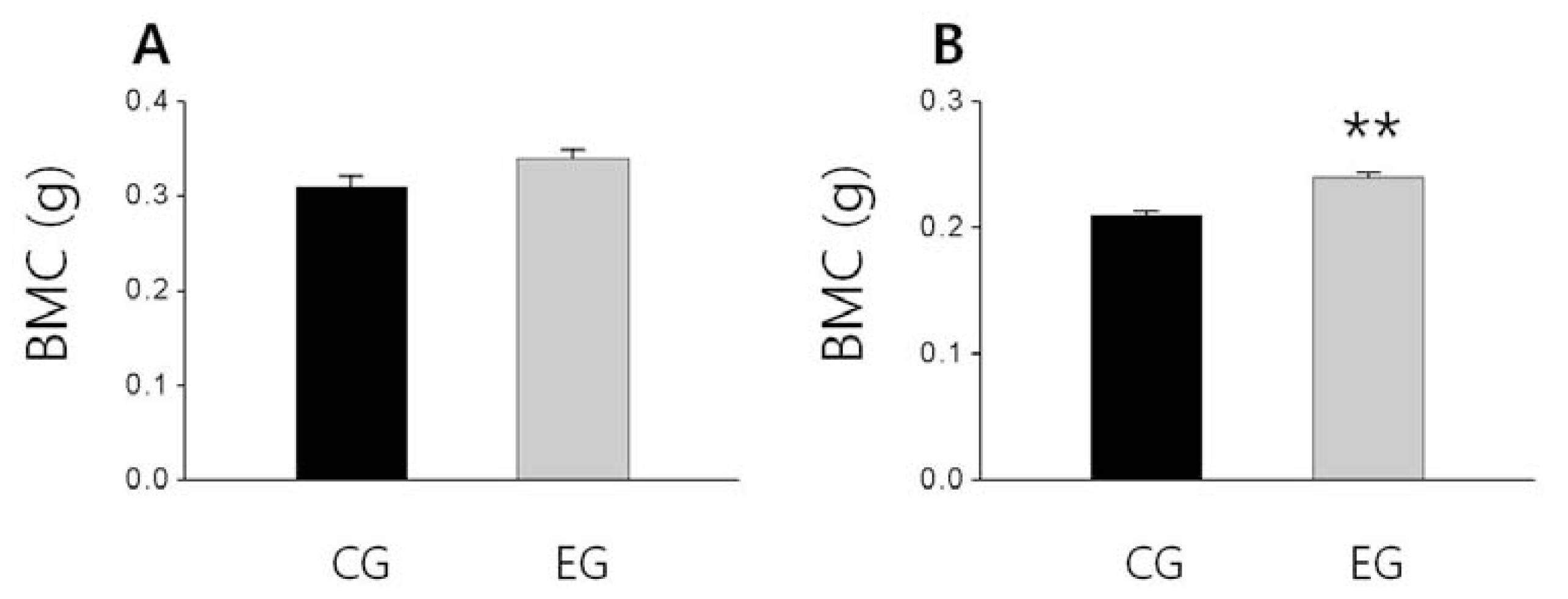
2.3. Bone Weight, BMD and BMC
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Preparation of Experimental Diet
4.3. Serum Analysis
4.4. Bone Weight, BMD and BMC Analysis
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
References
- NIH Consensus development panel on osteoporosis prevention diagnosis and therapy. Osteoporosis prevention, diagnosis, and therapy. J. Am. Med. Assoc. 2001, 285, 785–795.
- Smith, D.M.; Nance, W.E.; Kang, K.W.; Chrustian, J.C.; Johnstone, C.C. Genetic factors in determining bone mass. J. Clin. Invert 1973, 52, 2800–2808. [Google Scholar]
- Rana, A.R.; Michalsky, M.P.; Teich, S.; Groner, J.I.; Caniano, D.A.; Schuster, D.P. Childhood obesity: A risk factor for injuries observed at a level-1 trauma center. J. Pediatr. Surg 2009, 44, 1601–1605. [Google Scholar]
- Rocher, E.; Chappard, C.; Jaffre, C.; Benhamou, C.L.; Courteix, D. Bone mineral density in prepubertal obese and control children: Relation to body weight, lean mass, and fat mass. J. Bone Miner. Metab 2008, 26, 73–78. [Google Scholar]
- Chan, G.; Hoffman, K.; Mcmurry, M. Effects of dairy products on bone and body composition in pubertal girls. J. Pediatr 1995, 126, 551–556. [Google Scholar]
- Heaney, R.P. Nutritional factors in osteoporosis. Annu. Rev. Nutr 1993, 13, 287–316. [Google Scholar]
- Fassler, A.L.C.; Bonjour, J.P. Osteoporosis as a pediatric problem. Pediatr. Clin. North Am 1995, 42, 811–824. [Google Scholar]
- Rude, R.K.; Oldham, S.H. Disorders of Magnesium Metabolism. In The metabolic and molecular Basis of Acquired Disease; Cohen, R.D., Lewis, B., Alberti, K.G.M.M., Denmon, A.M., Eds.; Balliere Tindall: London, UK, 1990; pp. 1124–1148. [Google Scholar]
- Stending-Lindberg, G.; Tepper, R.; Leichter, I. Trabecular bone density in a two-year controlled trial of peroral magnesium in osteoporosis. Magnesium Res 1993, 6, 155–163. [Google Scholar]
- Rude, R.K.; Oldham, S.B.; Sharp, C.F., Jr; Singer, F.R. Parathyroid hormone secretion in magnesium deficiency. J. Clin. Endocrinol. Metab. 1978, 47, 800–806. [Google Scholar]
- Reginster, J.Y.; Strause, L.; Deroisy, R.; Lecart, M.P.; Saltman, P.; Franchimont, P. Preliminary report of decreased serum magnesium in postmenopausal osteoporosis. Magnesium 1989, 8, 106–109. [Google Scholar]
- Campbell, L. Thyroid and parathyroid hormones and calcium homeostasis. Anaesth. Intens. Care Med 2011, 12, 465–468. [Google Scholar]
- Sahota, O.; Masud, T.; San, P.; Hosking, D.J. Vitamin D insufficiency increases bone turnover markers and enhances bone loss at the hips in patients with established osteoporosis. Clin. Endocrinol 1999, 51, 217–221. [Google Scholar]
- LeBoff, M.S.; Kohlmeier, L.; Hurwitz, S.; Franklin, J.; Wright, J.; Glowacki, J. Occult vitamin D deficiency in post menopausal US women with acute hip fracture. J. Am. Med. Assoc 1999, 281, 1505–1511. [Google Scholar]
- Cho, J.Y.; Park, S.C.; Kim, T.W.; Kim, K.S.; Song, J.C.; Kim, S.K.; Lee, H.M.; Sung, H.J.; Park, H.J.; Song, Y.B.; et al. Radical scavenging and anti-inflammatory activity of extracts from Opuntia humifusa Raf. J. Pharm. Pharmacol 2006, 58, 113–119. [Google Scholar]
- Hahm, S.W.; Park, J.; Son, Y.S. Opuntia humifusa stems lower blood glucose and cholesterol levels in streptozotocin-induced diabetic rats. Nutr. Res 2011, 31, 479–487. [Google Scholar]
- Park, E.H.; Kahng, J.H.; Paek, E.A. Studies on the pharmacological action of cactus: Identification of its anti-inflammatory effect. Arch. Pharm. Res 1998, 21, 20–34. [Google Scholar]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Phenolic composition, antioxidant capacity and in vitro cancer cell cytotoxicity of nine prickly pear (Opuntia spp.) juices. Plant Foods Hum. Nutr 2009, 64, 146–152. [Google Scholar]
- Lee, J.C.; Kim, H.R.; Kim, J.; Jang, Y.S. Antioxidant property of an ethanol extract of the stem of Opuntia ficus-indica var. Saboten. J. Agric. Food Chem 2002, 50, 6490–6496. [Google Scholar]
- Hernández-Urbiola, M.I.; Pérez-Torrero, E.; Rodríguez-García, M.E. Chemical analysis of nutritional content of prickly pads (Opuntia ficus indica) at varied ages in an organic harvest. Int. J. Environ. Res. Public Health 2011, 8, 1287–1295. [Google Scholar]
- Brown, J.P.; Delmas, P.D.; Malaval, L.; Chapuy, M.C.; Delmas, P.D.; Edouard, C.; Meunier, P.J. Serum bone Gla protein: A specific marker for bone formation in post-menopausal osteoporosis. Lancet 1984, 1, 1091–1093. [Google Scholar]
- Power, M.J.; Fottrell, P.F. Osteocalcin: Diagnostic methods and clinical applications. Crit. Rev. Clin. Lab. Sci 1991, 28, 287–335. [Google Scholar]
- Godfrey, J.K.; Kayser, B.D.; Gomez, G.V.; Bennett, J.; Jaque, S.V.; Sumida, K.D. Interrupted resistance training and BMD in growing rats. Int. J. Sports Med 2009, 30, 579–584. [Google Scholar]
- Carpenter, T.O.; Mackowiak, S.J.; Troiano, N.; Gundberg, C.M. Osteocalcin and its message: Relationship to bone histology in magnesium-deprived rats. Am. J. Physiol 1992, 263, 107–114. [Google Scholar]
- Broadus, A.E. Physiological Functions of Calcium, Magnesium, and Phosphorus and Mineral ion Balance. In Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism; Favus, M.J., Ed.; Raven Press: New York, NY, USA, 1993; pp. 41–46. [Google Scholar]
- Jagtap, V.R.; Ganu, J.V.; Nagane, N.S. BMD and serum intact osteocalcin in postmenopausal osteoporosis women. Indian J. Clin. Biochem 2011, 26, 70–73. [Google Scholar]
- Toba, Y.; Kajita, Y.; Masuyama, R.; Takada, Y.; Suzuki, K.; Aoe, S. Dietary magnesium supplementation affects bone metabolism and dynamic strength of bone in ovariectomized rats. J. Nutr 2000, 130, 216–220. [Google Scholar]
- Kumar, R.; Thompson, J.R. The regulation of parathyroid hormone secretion and synthesis. J. Am. Soc. Nephrol 2011, 22, 216–224. [Google Scholar]
- Creedon, A.; Cashman, K.D. The effect of calcium intake on bone composition and bone resorption in the young growing rat. Br. J. Nutr 2001, 86, 453–459. [Google Scholar]
- Reid, I.R.; Ames, R.W.; Evans, M.C.; Gamble, G.D.; Sharpe, S.J. Effect of calcium supplementation on bone loss in postmenopausal women. N. Engl. J. Med 1993, 328, 460–464. [Google Scholar]
- Grados, F.; Brazier, M.; Kamel, S.; Duver, S.; Heurtebize, N.; Maamer, M.; Mathieu, M.; Garabédian, M.; Sebert, J.L.; Fardellone, P. Effects on bone mineral density of calcium and vitamin D supplementation in elderly women with vitamin D deficiency. Joint Bone Spine 2003, 70, 203–208. [Google Scholar]
- Rude, R.K.; Adams, J.S.; Ryzen, E.; Endres, D.B.; Niimi, H.; Horst, R.L.; Haddad, J.G., Jr; Singer, F.R. Low serum concentrations of 1,25-dihydroxyvitamin and in human magnesium deficiency. J. Clin. Endocrinol. Metab. 1985, 61, 933–940. [Google Scholar]
- Rude, R.K.; Gruber, H.E.; Norton, H.J.; Wei, L.Y.; Frausto, A.; Mills, B.G. Bone loss induced by dietary magnesium reduction to 10% of the nutrient requirement in rats is associated with increased release of substance P and tumor necrosis factor-α. J. Nutr 2004, 134, 79–85. [Google Scholar]
- Kenney, M.A.; McCoy, H.; Williams, L. Effects of magnesium deficiency on strength, mass and composition of rat femur. Calcif. Tissue Int 1994, 54, 44–49. [Google Scholar]
- Boskey, A.L.; Rimnac, C.M.; Bansal, M.; Federman, M.; Lian, J.; Boyan, B.D. Effect of short-term hypomagnesemia on the chemical and mechanical properties of rat bone. J. Orthop. Res 1992, 10, 774–783. [Google Scholar]
- Rude, R.K.; Kirchen, M.E.; Gruber, H.E.; Meyer, M.H.; Luck, J.S.; Crawford, D.L. Magnesium deficiency-induced osteoporosis in the rat: Uncoupling of bone formation and bone resorption. Magnes. Res 1999, 12, 257–267. [Google Scholar]
- Kang, J.; Kwon, D.; Park, J.; Shin, Y.O.; Lee, J.B.; Igawa, S.; Song, Y. Deep-sea water supplementation and swimming exercise improves bone metabolism in growing rats fed a high-fat diet. Food Sci. Biotechnol 2011, 20, 665–670. [Google Scholar]
- Bogden, J.D.; Kemp, F.W.; Huang, A.E.; Shapses, S.A.; Ambia-Sobhan, H.; Jagpal, S.; Brown, I.L.; Birkett, A.M. Bone mineral density and content during weight cycling in female rats: Effects of dietary amylase-resistant starch. Nutr. Metab 2008, 26, 5–34. [Google Scholar]
- Kim, Y.J.; Park, T.S. Genes are differentially expressed in the epididymal fat of rats rendered obese by a high-fat diet. Nutr. Res 2008, 28, 414–422. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| CG | EG | |
|---|---|---|
| Initial body weight (g) | 235.9 ± 3.89 | 235.8 ± 3.97 |
| Final body weight (g) | 428.2 ± 10.86 | 419.3 ± 14.67 |
| FER | 0.26 ± 0.008 | 0.26 ± 0.011 |
| Food intake (g/day) | 16.2 ± 0.38 | 14.9 ± 0.16 |
| CG | EG | |
|---|---|---|
| Ca (mg/dL) | 10.5 ± 0.14 | 10.2 ± 0.21 |
| P (mg/dL) | 7.4 ± 0.33 | 7.6 ± 0.17 |
| ALP (IU/L) | 87.0 ± 2.55 | 91.0 ± 3.18 |
| OC (ng/mL) | 25.8 ± 0.53 | 30.4 ± 1.41 * |
| PTH (pg/mL) | 13.7 ± 0.88 | 10.7 ± 0.45 * |
| Ingredients | Contents |
|---|---|
| moisture (% w/w) | 2.89 |
| ash (% w/w) | 13.8 |
| carbohydrate (g/100 g) | 46.56 |
| crude protein (g/100 g) | 4.91 |
| crude fat (g/100 g) | 3.06 |
| fiber (g/100 g) | 28.78 |
| Fe2+ (mg/g) | 5.76 |
| Ca2+ (mg/100 g) | 2931.3 |
| Mg2+ (mg/100 g) | 1227.9 |
| K+ (mg/100 g) | 2155.5 |
| Na+ (mg/100 g) | 30.9 |
| P2+ (mg/100 g) | 653.2 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kang, J.; Park, J.; Choi, S.H.; Igawa, S.; Song, Y. Opuntia humifusa Supplementation Increased Bone Density by Regulating Parathyroid Hormone and Osteocalcin in Male Growing Rats. Int. J. Mol. Sci. 2012, 13, 6747-6756. https://doi.org/10.3390/ijms13066747
Kang J, Park J, Choi SH, Igawa S, Song Y. Opuntia humifusa Supplementation Increased Bone Density by Regulating Parathyroid Hormone and Osteocalcin in Male Growing Rats. International Journal of Molecular Sciences. 2012; 13(6):6747-6756. https://doi.org/10.3390/ijms13066747
Chicago/Turabian StyleKang, Junyong, Jinho Park, Seong Hee Choi, Shoji Igawa, and Youngju Song. 2012. "Opuntia humifusa Supplementation Increased Bone Density by Regulating Parathyroid Hormone and Osteocalcin in Male Growing Rats" International Journal of Molecular Sciences 13, no. 6: 6747-6756. https://doi.org/10.3390/ijms13066747