By-Passing Large Screening Experiments Using Sequencing as a Tool to Identify scFv Fragments Targeting Atherosclerotic Lesions in a Novel In Vivo Phage Display Selection

Abstract
:
1. Introduction
2. Results and Discussion
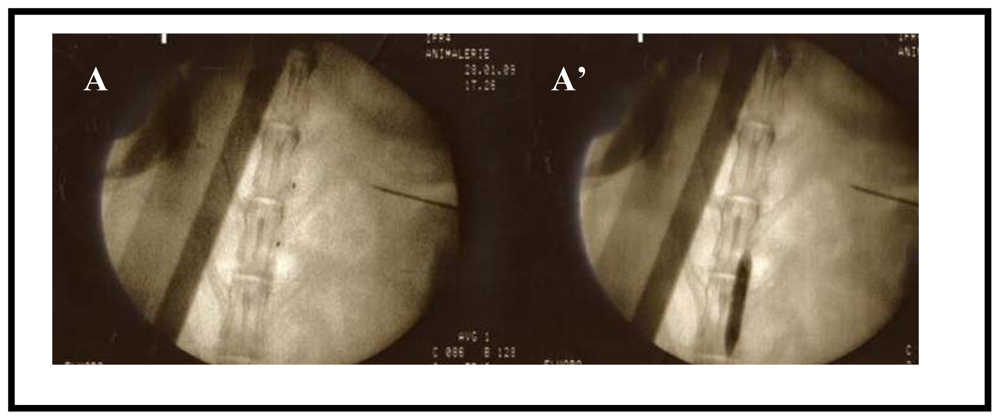
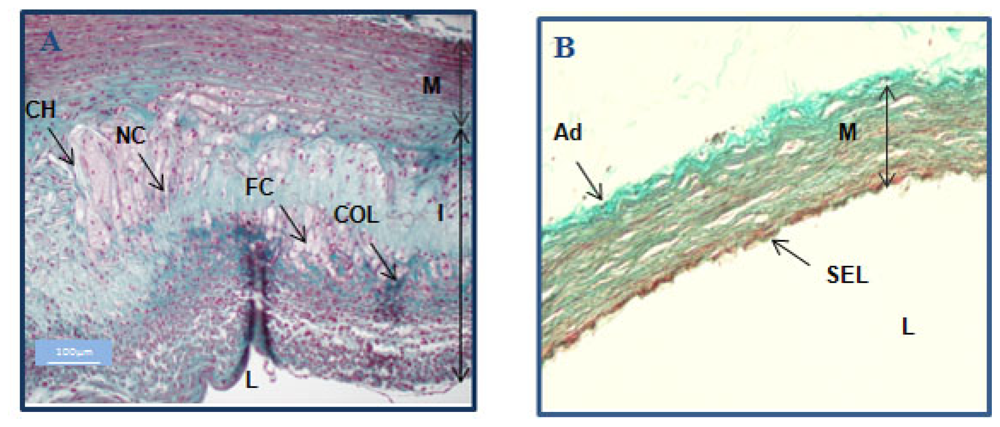
2.1. Rabbit model of Atherosclerosis
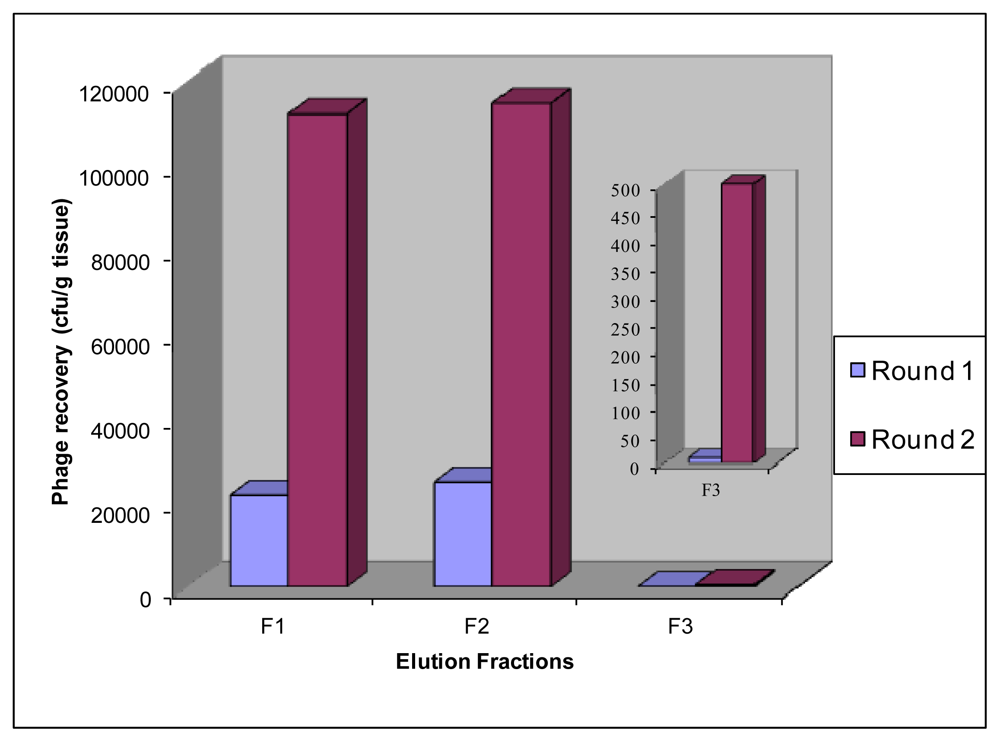
2.2. In Vivo Biopanning
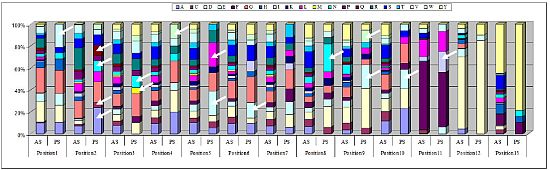
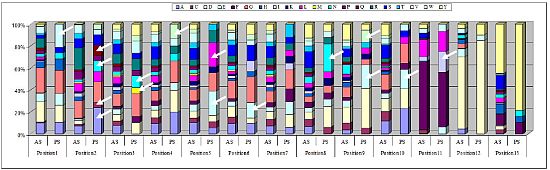
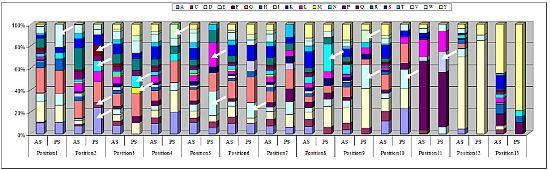
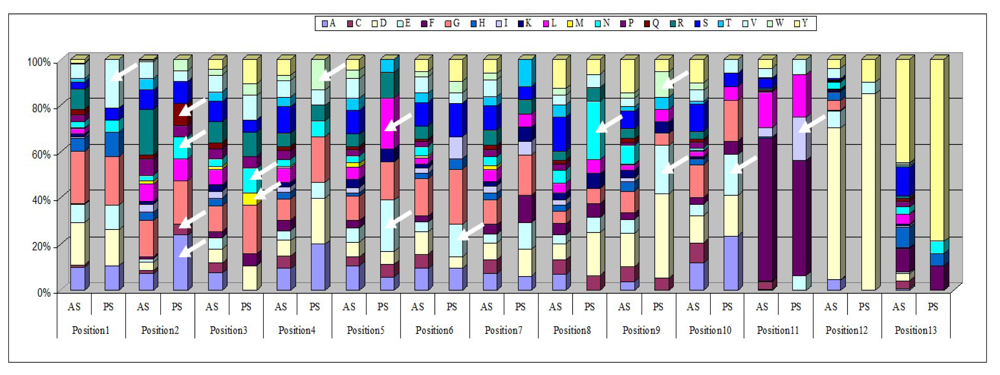
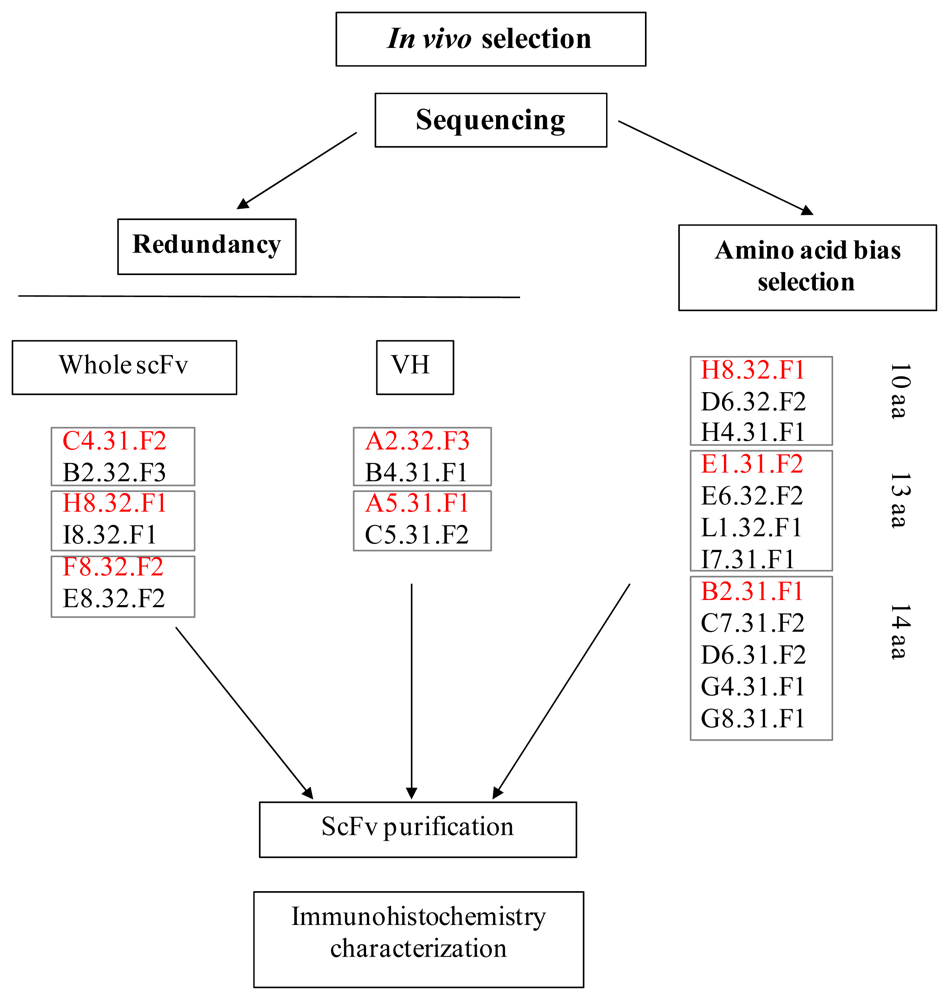
2.3. Identification of in Vivo-Selected Clones by Sequencing
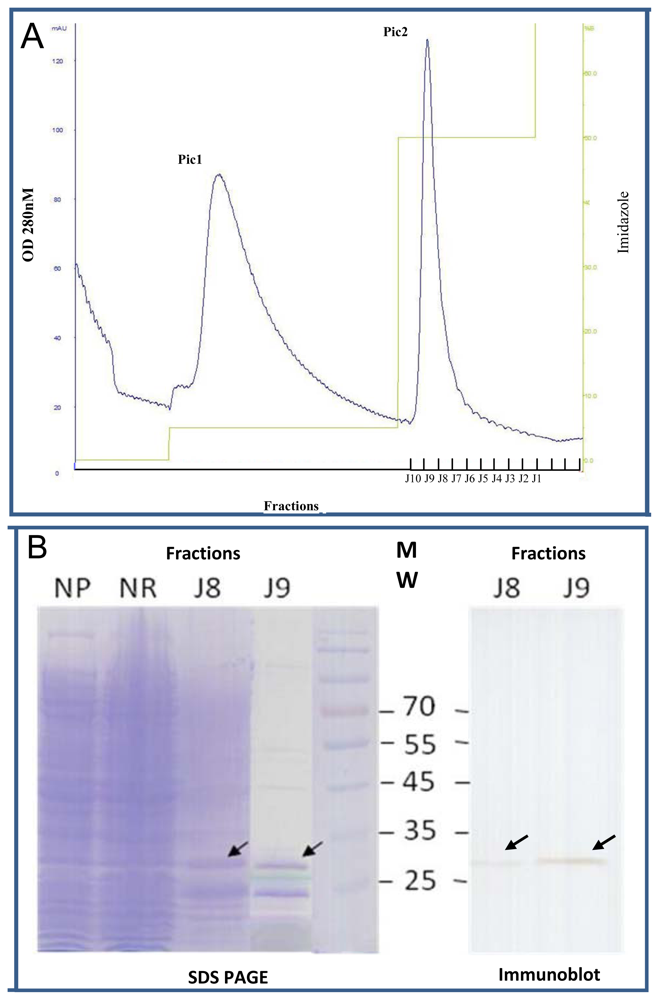
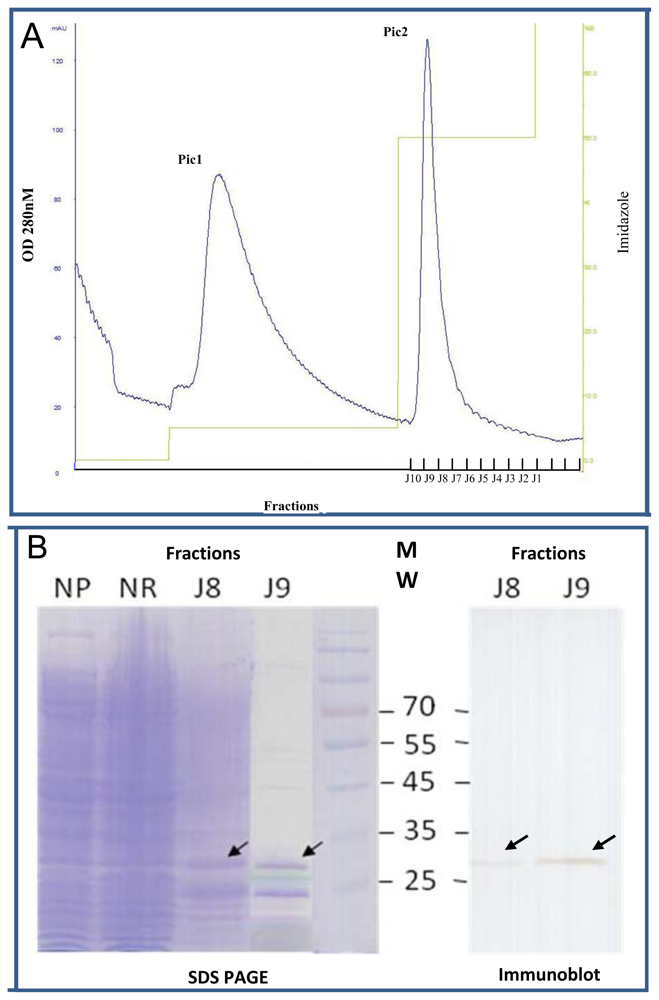
2.4. Expression of Selected Clones as Soluble scFv Fragments
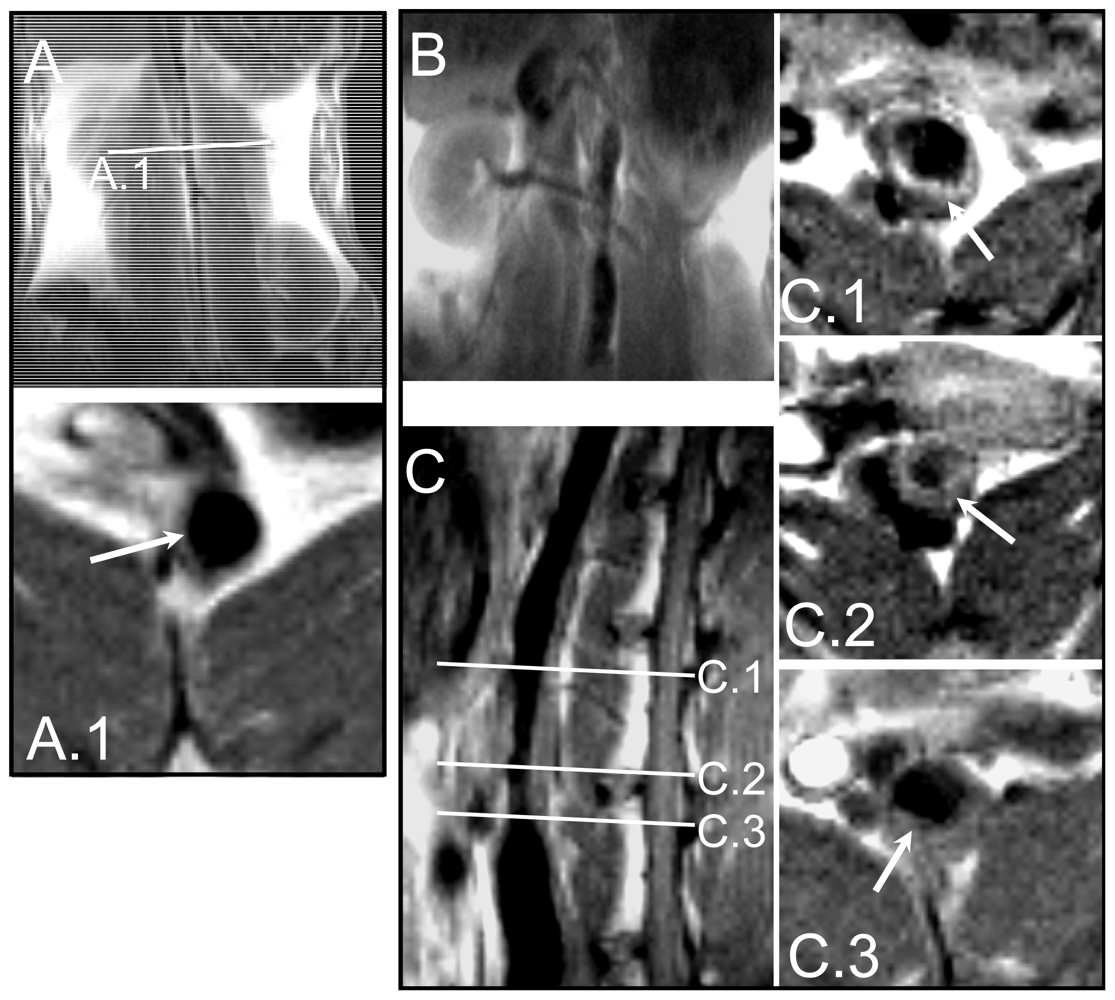
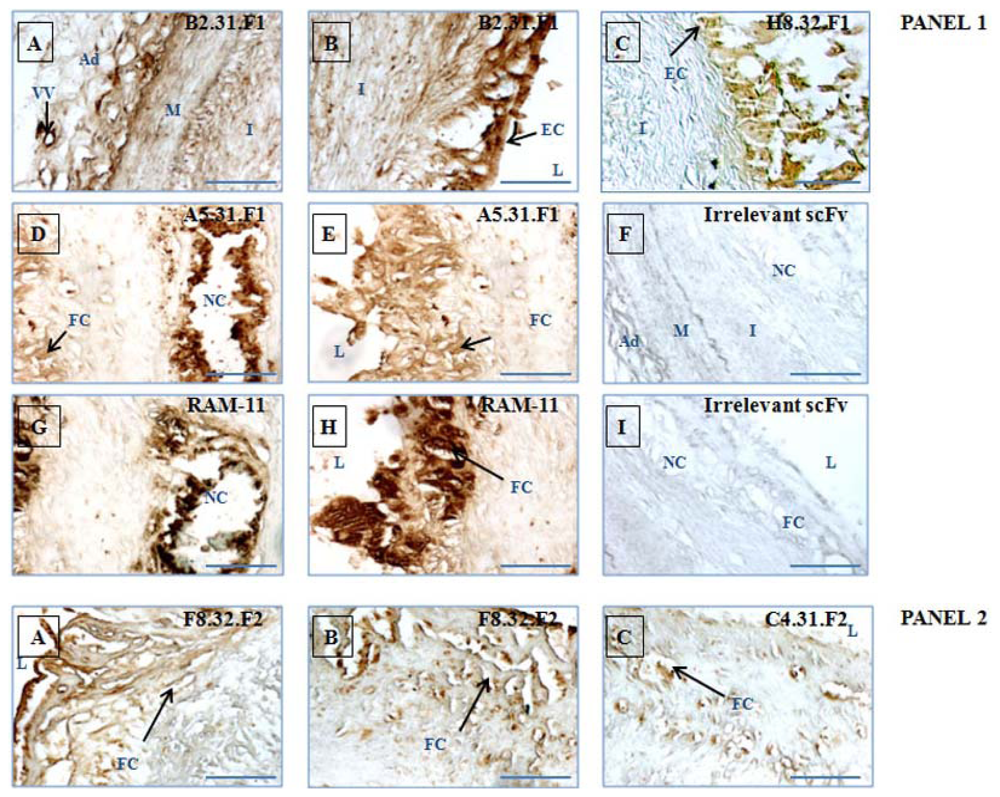
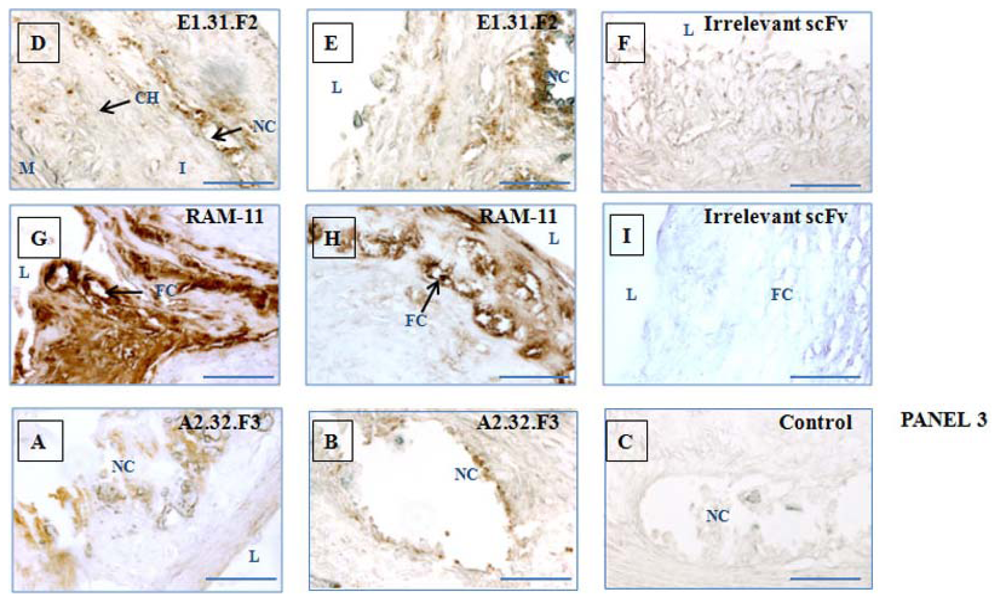
2.5. Immunohistochemical Validation of scFv Human Antibody Fragments Selected by Sequencing
3. Experimental Section
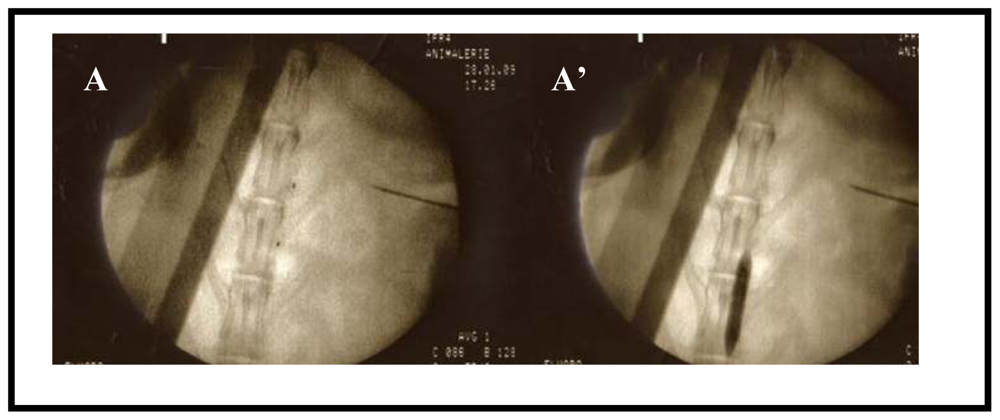
3.1. Animal Model
3.2. Magnetic Resonance Imaging
3.3. Human scFv Phage Library
3.4. In Vivo Biopanning in the Atherosclerotic New Zealand Rabbit Model
3.5. Sequencing of Randomly Picked Clones
3.6. Soluble scFv Production from Bacterial Culture
3.7. scFvs Purification on Nickel-Affinity Chromatography
3.8. Histological Examination
3.9. Immunohistochemical Analysis of Rabbit Sections
4. Conclusions
Acknowledgments
References
- Blanco-Colio, L.M.; Martin-Ventura, J.L.; Vivanco, F.; Michel, J.B.; Meilhac, O.; Egido, J. Biology of atherosclerotic plaques: What we are learning from proteomic analysis. Cardiovasc. Res 2006, 72, 18–29. [Google Scholar]
- Wickline, S.A.; Neubauer, A.M.; Winter, P.M.; Caruthers, S.D.; Lanza, G.M. Molecular imaging and therapy of atherosclerosis with targeted nanoparticles. J. Magn. Reson. Imaging 2007, 25, 667–680. [Google Scholar]
- Shaw, S.Y. Molecular imaging in cardiovascular disease: Targets and opportunities. Nat. Rev. Cardiol 2009, 6, 569–579. [Google Scholar]
- Antoniades, C.; Psarros, C.; Tousoulis, D.; Bakogiannis, C.; Shirodaria, C.; Stefanadis, C. Nanoparticles: A promising therapeutic approach in atherosclerosis. Curr. Drug Deliv 2010, 7, 303–311. [Google Scholar]
- Koenig, W.; Khuseyinova, N. Biomarkers of atherosclerotic plaque instability and rupture. Arterioscler. Thromb. Vasc. Biol 2007, 27, 15–26. [Google Scholar]
- Briley-Saebo, K.C.; Mulder, W.J.; Mani, V.; Hyafil, F.; Amirbekian, V.; Aguinaldo, J.G.; Fisher, E.A.; Fayad, Z.A. Magnetic resonance imaging of vulnerable atherosclerotic plaques: Current imaging strategies and molecular imaging probes. J. Magn. Reson. Imaging 2007, 26, 460–479. [Google Scholar]
- Rader, D.J.; Daugherty, A. Translating molecular discoveries into new therapies for atherosclerosis. Nature 2008, 451, 904–913. [Google Scholar]
- Sanz, J.; Fayad, Z.A. Imaging of atherosclerotic cardiovascular disease. Nature 2008, 451, 953–957. [Google Scholar]
- Weissleder, R.; Kelly, K.; Sun, E.Y.; Shtatland, T.; Josephson, L. Cell-specific targeting of nanoparticles by multivalent attachment of small molecules. Nat. Biotechnol 2005, 23, 1418–1423. [Google Scholar]
- Lam, K.S.; Salmon, S.E.; Hersh, E.M.; Hruby, V.J.; Kazmierski, W.M.; Knapp, R.J. A new type of synthetic peptide library for identifying ligand-binding activity. Nature 1991, 354, 82–84. [Google Scholar]
- Alvim, J., Jr; Severino, R.P.; Marques, E.F.; Martinelli, A.M.; Vieira, P.C.; Fernandes, J.B.; da Silva, M.F.; Correa, A.G. Solution phase synthesis of a combinatorial library of chalcones and flavones as potent cathepsin V inhibitors. J. Comb. Chem. 2010, 12, 687–695. [Google Scholar]
- Aina, O.H.; Liu, R.; Sutcliffe, J.L.; Marik, J.; Pan, C.X.; Lam, K.S. From combinatorial chemistry to cancer-targeting peptides. Mol. Pharm 2007, 4, 631–651. [Google Scholar]
- Toepert, F.; Knaute, T.; Guffler, S.; Pires, J.R.; Matzdorf, T.; Oschkinat, H.; Schneider-Mergener, J. Combining SPOT synthesis and native peptide ligation to create large arrays of WW protein domains. Angew. Chem. Int. Ed. Engl 2003, 42, 1136–1140. [Google Scholar]
- Iwamoto, S.; Nishimichi, N.; Tateishi, Y.; Sato, Y.; Horiuchi, H.; Furusawa, S.; Sawamura, T.; Matsuda, H. Generation and characterization of chicken monoclonal antibodies against human LOX-1. MAbs 2009, 1, 357–363. [Google Scholar]
- Hong, H.Y.; Lee, H.Y.; Kwak, W.; Yoo, J.; Na, M.H.; So, I.S.; Kwon, T.H.; Park, H.S.; Huh, S.; Oh, G.T.; et al. Phage display selection of peptides that home to atherosclerotic plaques: IL-4 receptor as a candidate target in atherosclerosis. J. Cell. Mol. Med 2008, 12, 2003–2014. [Google Scholar]
- Burtea, C.; Laurent, S.; Port, M.; Lancelot, E.; Ballet, S.; Rousseaux, O.; Toubeau, G.; Vander Elst, L.; Corot, C.; Muller, R.N. Magnetic resonance molecular imaging of vascular cell adhesion molecule-1 expression in inflammatory lesions using a peptide-vectorized paramagnetic imaging probe. J. Med. Chem 2009, 52, 4725–4742. [Google Scholar]
- Yao, V.J.; Ozawa, M.G.; Trepel, M.; Arap, W.; McDonald, D.M.; Pasqualini, R. Targeting pancreatic islets with phage display assisted by laser pressure catapult microdissection. Am. J. Pathol 2005, 166, 625–636. [Google Scholar]
- Kelly, K.A.; Nahrendorf, M.; Yu, A.M.; Reynolds, F.; Weissleder, R. In vivo phage display selection yields atherosclerotic plaque targeted peptides for imaging. Mol. Imaging Biol 2006, 8, 201–207. [Google Scholar]
- Liu, C.; Bhattacharjee, G.; Boisvert, W.; Dilley, R.; Edgington, T. In vivo interrogation of the molecular display of atherosclerotic lesion surfaces. Am. J. Pathol 2003, 163, 1859–1871. [Google Scholar]
- Houston, P.; Goodman, J.; Lewis, A.; Campbell, C.J.; Braddock, M. Homing markers for atherosclerosis: Applications for drug delivery, gene delivery and vascular imaging. FEBS Lett 2001, 492, 73–77. [Google Scholar]
- Krag, D.N.; Shukla, G.S.; Shen, G.P.; Pero, S.; Ashikaga, T.; Fuller, S.; Weaver, D.L.; Burdette-Radoux, S.; Thomas, C. Selection of tumor-binding ligands in cancer patients with phage display libraries. Cancer Res 2006, 66, 7724–7733. [Google Scholar]
- Philibert, P.; Stoessel, A.; Wang, W.; Sibler, A.P.; Bec, N.; Larroque, C.; Saven, J.G.; Courtete, J.; Weiss, E.; Martineau, P. A focused antibody library for selecting scFvs expressed at high levels in the cytoplasm. BMC Biotechnol 2007, 7. [Google Scholar] [CrossRef]
- Rekhter, M.D.; Hicks, G.W.; Brammer, D.W.; Work, C.W.; Kim, J.S.; Gordon, D.; Keiser, J.A.; Ryan, M.J. Animal model that mimics atherosclerotic plaque rupture. Circ. Res 1998, 83, 705–713. [Google Scholar]
- Phinikaridou, A.; Hallock, K.J.; Qiao, Y.; Hamilton, J.A. A robust rabbit model of human atherosclerosis and atherothrombosis. J. Lipid Res 2009, 50, 787–797. [Google Scholar]
- Zou, J.; Dickerson, M.T.; Owen, N.K.; Landon, L.A.; Deutscher, S.L. Biodistribution of filamentous phage peptide libraries in mice. Mol. Biol. Rep 2004, 31, 121–129. [Google Scholar]
- Deramchia, K.; Jacobin-Valat, M.J.; Vallet, A.; Bazin, H.; Santarelli, X.; Sanchez, S.; Dos Santos, P.; Franconi, J.M.; Claverol, S.; Bonetto, S.; Clofent-Sanchez, G. New human antibody fragments homing to atherosclerotic endothelial and subendothelial tissues: An in vivo phage display targeting human antibodies homing to atherosclerotic tissues. Am. J. Pathol 2012, in press. [Google Scholar]
- De Wildt, R.M.; Mundy, C.R.; Gorick, B.D.; Tomlinson, I.M. Antibody arrays for high-throughput screening of antibody-antigen interactions. Nat. Biotechnol 2000, 18, 989–994. [Google Scholar]
- Honegger, A.; Pluckthun, A. Yet another numbering scheme for immunoglobulin variable domains: An automatic modeling and analysis tool. J. Mol. Biol 2001, 309, 657–670. [Google Scholar]
- AHo’s Amazing Atlas of Antibody Anatomy. Available online: http://www.bioc.uzh.ch/antibody/Structures/AgContact/VHProteinContactFrame.html accessed on 30 April 2012.
- AHo’s Amazing Atlas of Antibody Anatomy. Available online: http://www.bioc.uzh.ch/antibody/Structures/AgContact/VLProteinContactFrame.html accessed on 30 April 2012.
- Wilson, I.A.; Stanfield, R.L. Antibody-antigen interactions: New structures and new conformational changes. Curr. Opin. Struct. Biol 1994, 4, 857–867. [Google Scholar]
- Tsukada, T.; Rosenfeld, M.; Ross, R.; Gown, A.M. Immunocytochemical analysis of cellular components in atherosclerotic lesions. Use of monoclonal antibodies with the Watanabe and fat-fed rabbit. Arteriosclerosis 1986, 6, 601–613. [Google Scholar]
- IMGT/V-QUEST, IMGT/V-QUEST Programme, version 3.2.25. Available online: http://www.imgt.org/IMGT_vquest/share/textes/ accessed on 24 April 2012.
- Arap, W.; Kolonin, M.G.; Trepel, M.; Lahdenranta, J.; Cardo-Vila, M.; Giordano, R.J.; Mintz, P.J.; Ardelt, P.U.; Yao, V.J.; Vidal, C.I.; et al. Steps toward mapping the human vasculature by phage display. Nat. Med 2002, 8, 121–127. [Google Scholar]
- Rajotte, D.; Arap, W.; Hagedorn, M.; Koivunen, E.; Pasqualini, R.; Ruoslahti, E. Molecular heterogeneity of the vascular endothelium revealed by in vivo phage display. J. Clin. Invest 1998, 102, 430–437. [Google Scholar]
- Robert, R.; Jacobin-Valat, M.J.; Daret, D.; Miraux, S.; Nurden, A.T.; Franconi, J.M.; Clofent-Sanchez, G. Identification of human scFvs targeting atherosclerotic lesions: Selection by single round in vivo phage display. J. Biol. Chem 2006, 281, 40135–40143. [Google Scholar]
- Johns, M.; George, A.J.; Ritter, M.A. In vivo selection of sFv from phage display libraries. J. Immunol. Methods 2000, 239, 137–151. [Google Scholar]
- Ueberberg, S.; Meier, J.J.; Waengler, C.; Schechinger, W.; Dietrich, J.W.; Tannapfel, A.; Schmitz, I.; Schirrmacher, R.; Koller, M.; Klein, H.H.; et al. Generation of novel single-chain antibodies by phage-display technology to direct imaging agents highly selective to pancreatic beta- or alpha-cells in vivo. Diabetes 2009, 58, 2324–2334. [Google Scholar]
- Chertok, B.; Cole, A.J.; David, A.E.; Yang, V.C. Comparison of electron spin resonance spectroscopy and inductively-coupled plasma optical emission spectroscopy for biodistribution analysis of iron-oxide nanoparticles. Mol. Pharm 2010, 7, 375–385. [Google Scholar]
- Jacobin-Valat, M.J.; Deramchia, K.; Mornet, S.; Hagemeyer, C.E.; Bonetto, S.; Robert, R.; Biran, M.; Massot, P.; Miraux, S.; Sanchez, S.; et al. MRI of inducible P-selectin expression in human activated platelets involved in the early stages of atherosclerosis. NMR Biomed 2010, 24, 413–424. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VH CDR3 Length: 10 | |||||||||||||||||
| POSITION * | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||||||
| CONSENSUS ** | V | R | X | X | D/I | R | X | Y | X | V | E | D | |||||
| CLONES | F8.32.F2 | V | R | X | X | I | X | X | X | X | X | E | X | ||||
| H8.32.F1 | V | R | X | X | D | X | X | X | X | V | X | D | |||||
| D6.32.F2 | V | R | X | X | X | R | X | Y | X | X | E | X | |||||
| H4.32.F1 | V | R | X | X | D | X | X | Y | X | V | X | X | |||||
| VH CDR3 Length: 13 | |||||||||||||||||
| POSITION * | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | ||||
| CONSENSUS ** | V | R | V | A/C/N/Q | M/N | W | E/L | E | X | N | W/E | E | E | X | X | ||
| CLONES | E1.31.F2 | V | R | X | A | X | X | E | X | X | N | X | X | X | X | X | |
| E6.32F2 | V | R | X | N | X | X | L | X | X | N | X | X | X | X | X | ||
| L1.32F1 | V | R | V | N | N | X | X | X | X | N | E | X | X | X | X | ||
| I7.31.F1 | V | R | X | X | N | W | X | X | X | X | X | E | X | X | X | ||
| VH CDR3 Length: 14 | |||||||||||||||||
| POSITION * | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |||
| CONSENSUS** | V | R | E/S | D/E/M | W | I/R/W | P | W/M | G | X | E | X | X | Y | X | N | |
| CLONES | B2.32.F1 | V | R | E | X | X | W | P | X | X | X | E | X | X | X | X | X |
| C7.31CF2 | V | R | S | M | X | I | X | X | X | X | X | X | X | Y | X | X | |
| D6.31.F2 | V | R | E | E | X | W | X | X | X | X | X | X | X | X | X | X | |
| G4.31.F1 | V | R | X | D | X | R | X | M | X | X | X | X | X | X | X | N | |
| G8.31.F1 | V | R | E | X | W | X | X | W | G | X | X | X | X | X | X | X | |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Deramchia, K.; Jacobin-Valat, M.-J.; Laroche-Traineau, J.; Bonetto, S.; Sanchez, S.; Dos Santos, P.; Massot, P.; Franconi, J.-M.; Martineau, P.; Clofent-Sanchez, G. By-Passing Large Screening Experiments Using Sequencing as a Tool to Identify scFv Fragments Targeting Atherosclerotic Lesions in a Novel In Vivo Phage Display Selection. Int. J. Mol. Sci. 2012, 13, 6902-6923. https://doi.org/10.3390/ijms13066902
Deramchia K, Jacobin-Valat M-J, Laroche-Traineau J, Bonetto S, Sanchez S, Dos Santos P, Massot P, Franconi J-M, Martineau P, Clofent-Sanchez G. By-Passing Large Screening Experiments Using Sequencing as a Tool to Identify scFv Fragments Targeting Atherosclerotic Lesions in a Novel In Vivo Phage Display Selection. International Journal of Molecular Sciences. 2012; 13(6):6902-6923. https://doi.org/10.3390/ijms13066902
Chicago/Turabian StyleDeramchia, Kamel, Marie-Josee Jacobin-Valat, Jeanny Laroche-Traineau, Stephane Bonetto, Stephane Sanchez, Pierre Dos Santos, Philippe Massot, Jean-Michel Franconi, Pierre Martineau, and Gisele Clofent-Sanchez. 2012. "By-Passing Large Screening Experiments Using Sequencing as a Tool to Identify scFv Fragments Targeting Atherosclerotic Lesions in a Novel In Vivo Phage Display Selection" International Journal of Molecular Sciences 13, no. 6: 6902-6923. https://doi.org/10.3390/ijms13066902