Reference Gene Selection in the Desert Plant Eremosparton songoricum

Abstract
:1. Introduction
2. Results
2.1. Sequence Analysis of the Candidate Reference Genes
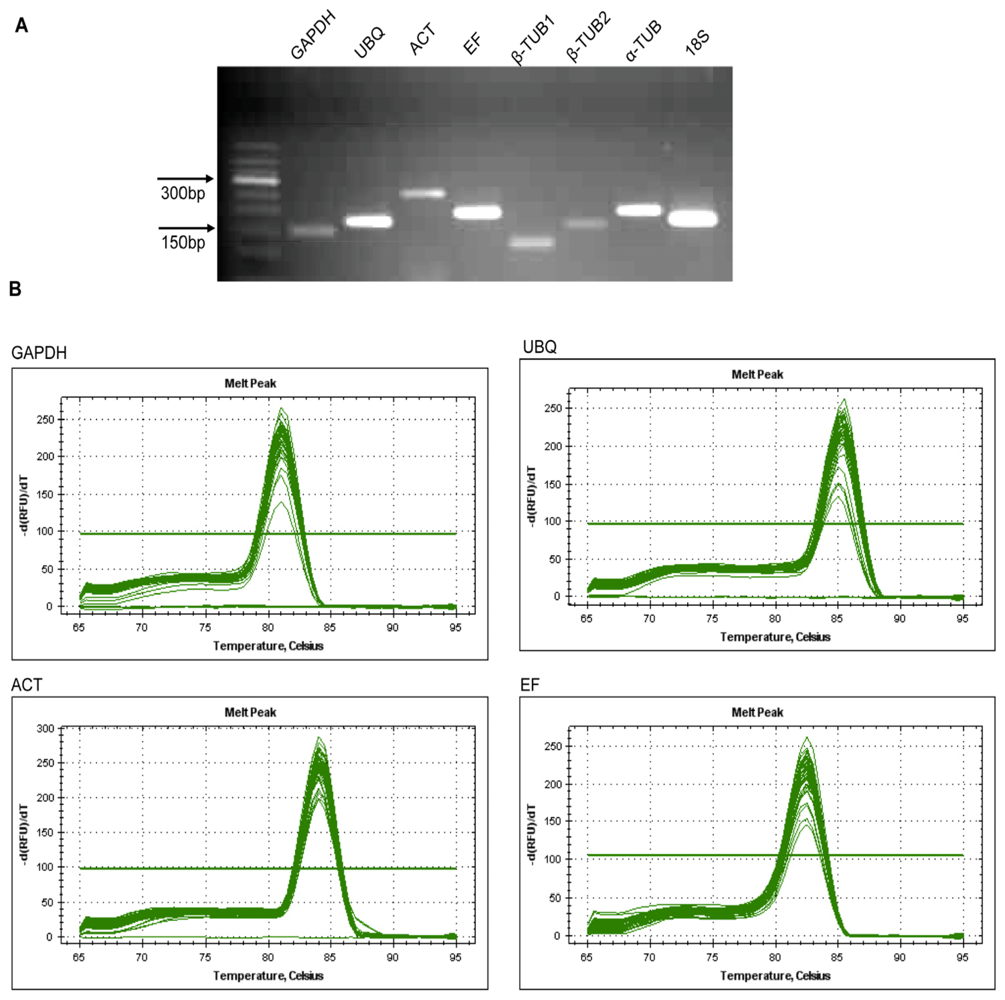
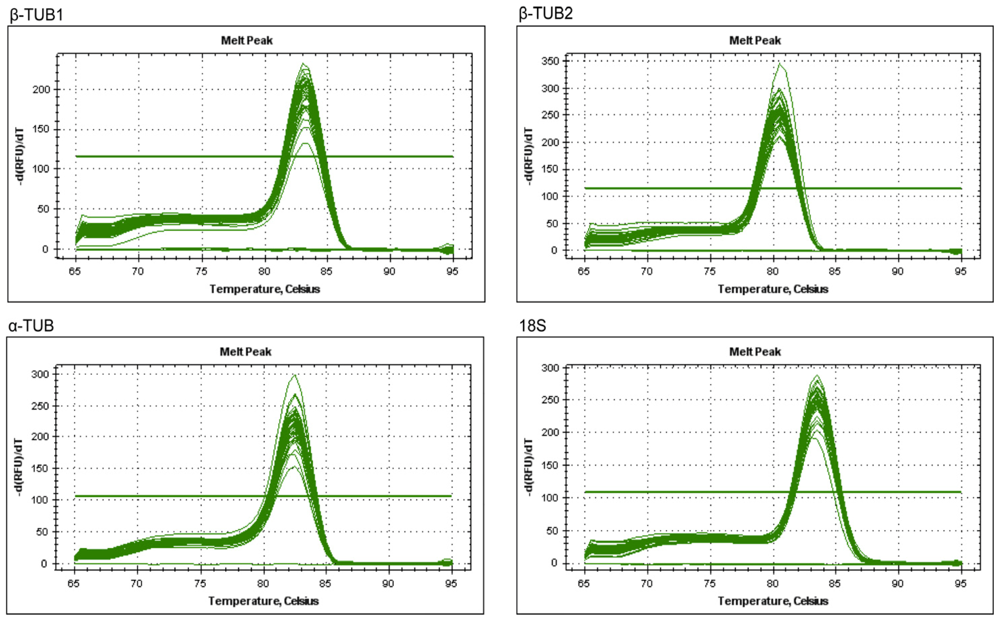
2.2. Verification of Primer Specificity and Efficiency
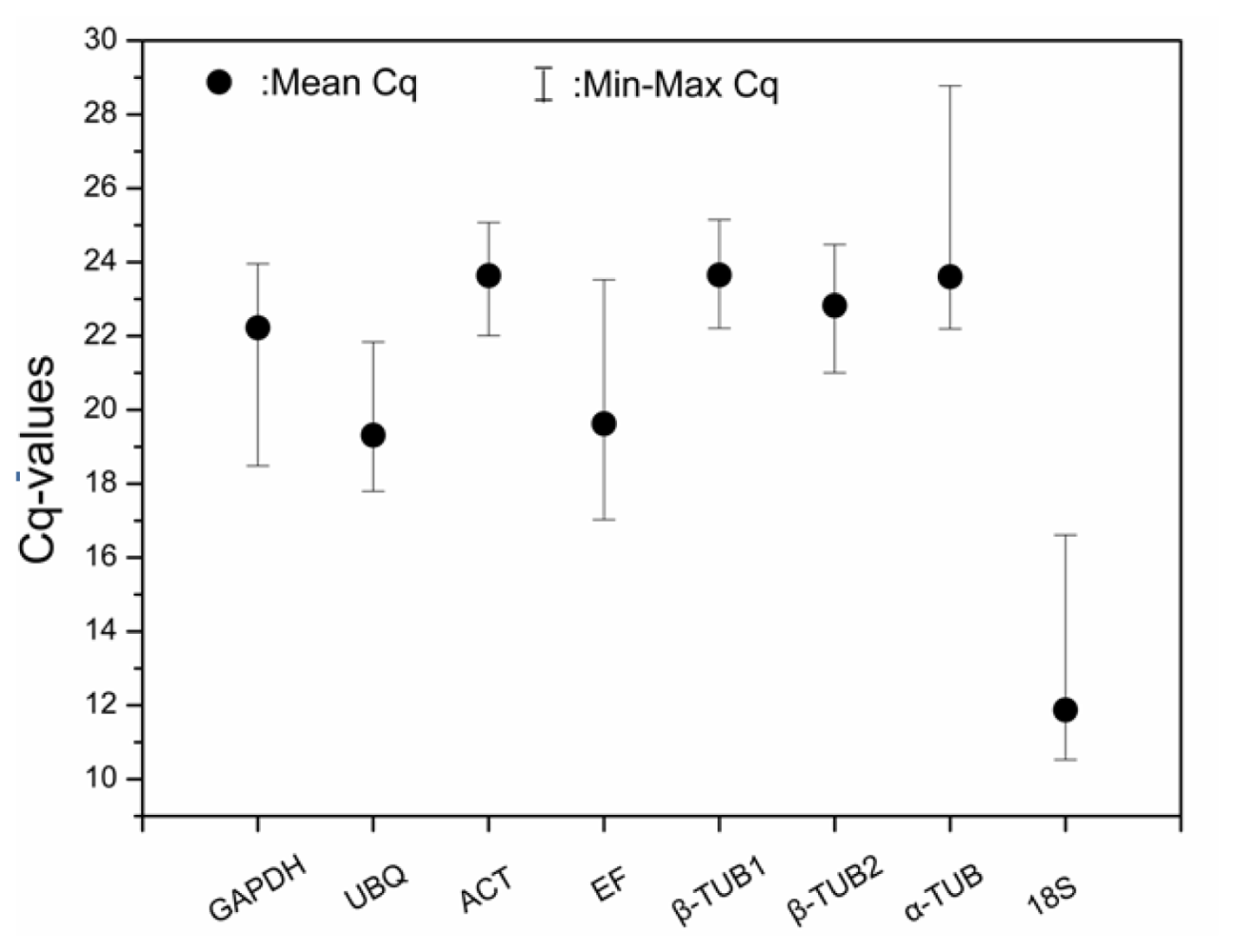
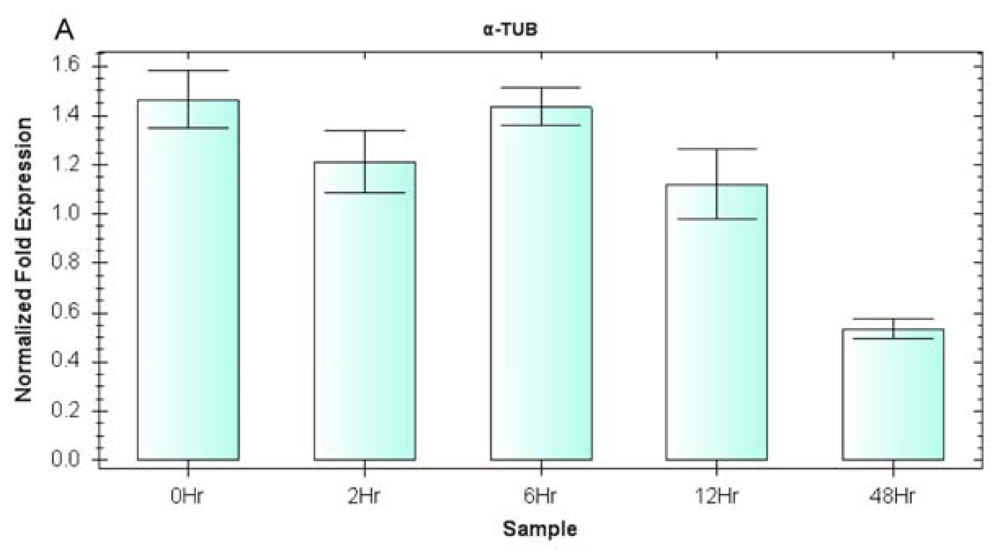
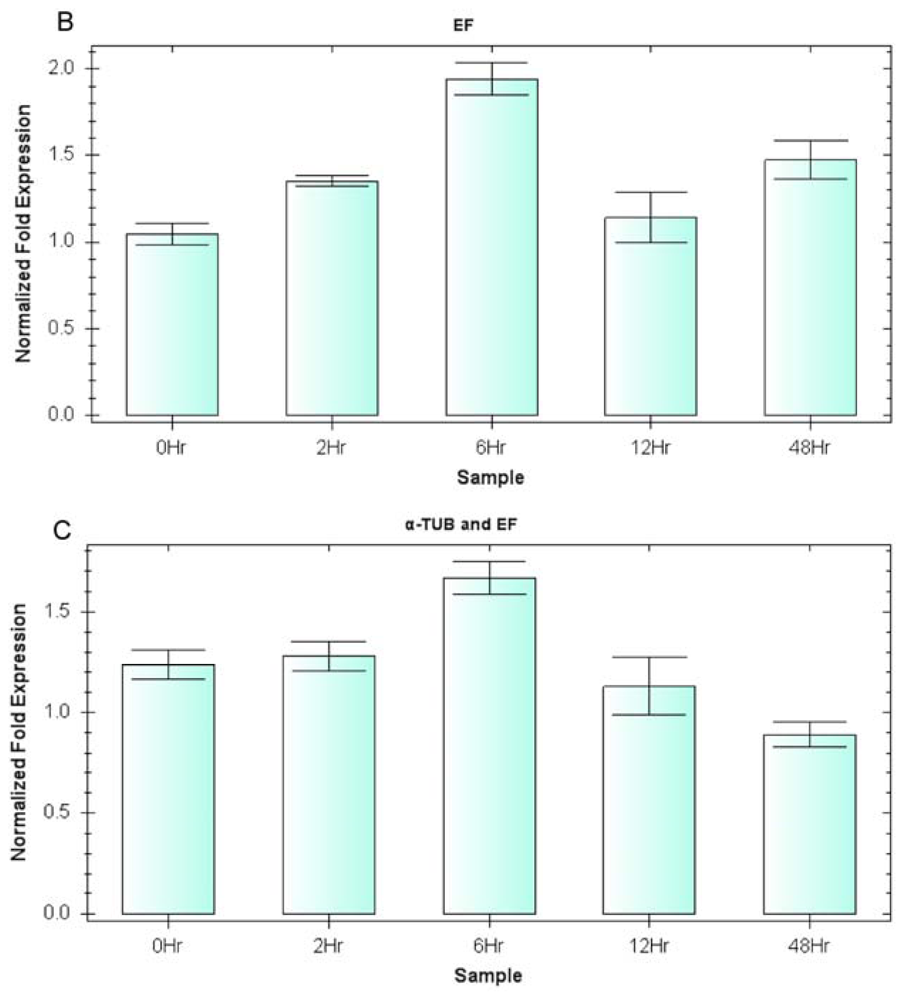
2.3. Expression Profiling of Reference Genes (qRT-PCR Assay)
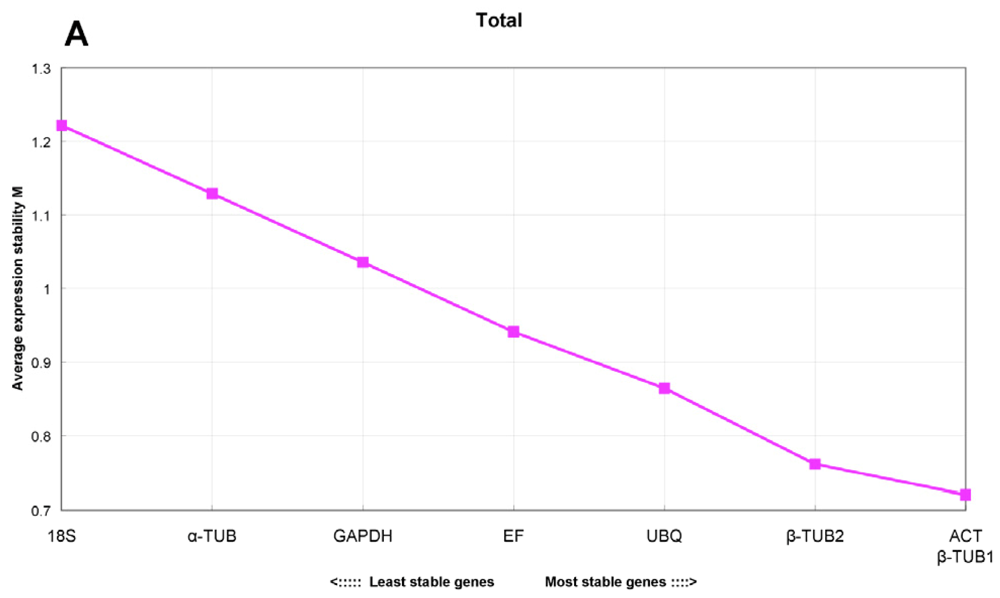
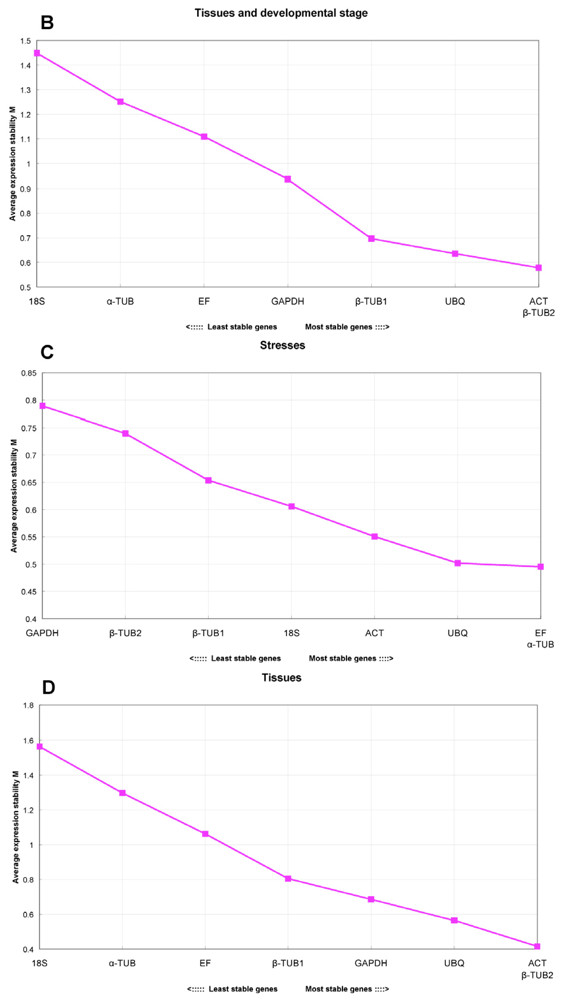
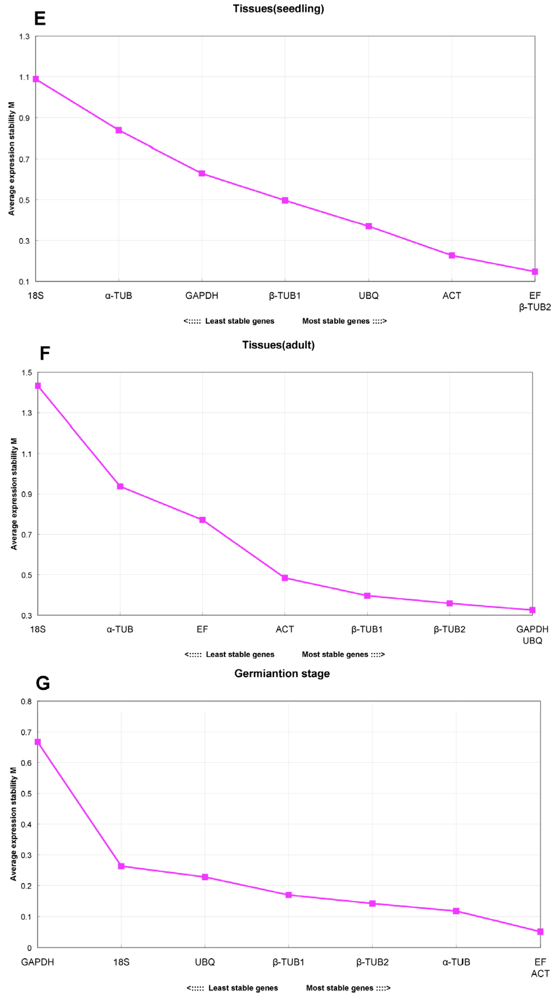
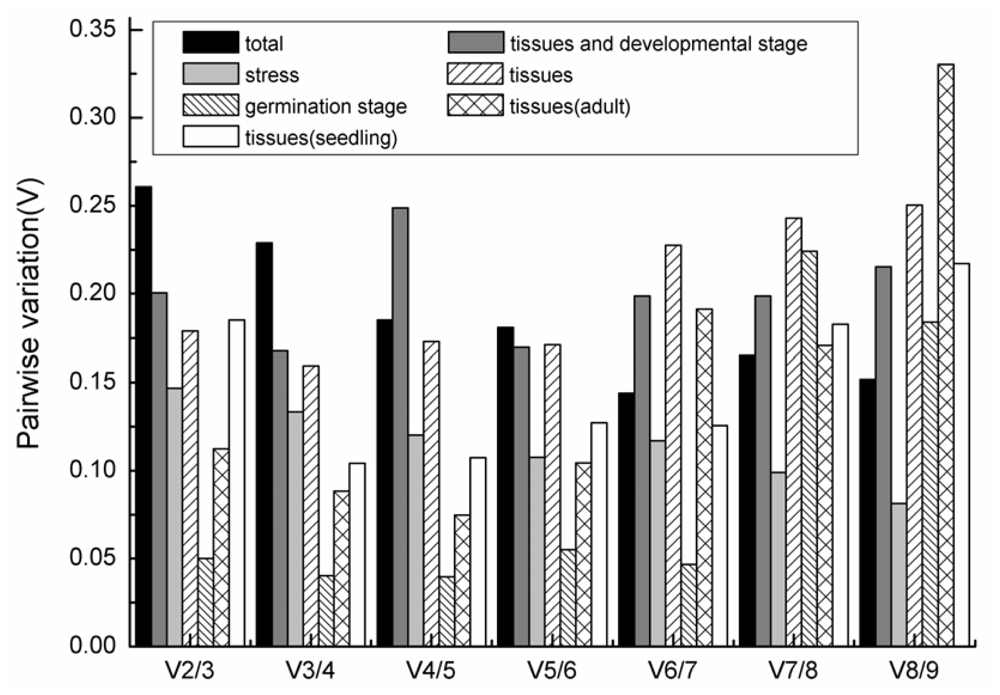
2.4. GeNorm Analysis
2.5. EsDREB2 Gene Expression
3. Discussion
4. Experimental Section
4.1. Plant Materials and Treatments
4.2. RNA Extraction and cDNA Synthesis
4.3. Cloning the Partial Sequences of the Candidate Reference Genes
4.4. QRT-PCR Primer Design and Testing
4.5. Quantitative Real-Time PCR
4.6. Statistical Analysis of Gene Expression Stability
4.7. Normalization of a Gene of Interest, EsDREB2
5. Conclusions
- For all the tested samples, ACT and β-TUB1 were the most stable reference genes, and seven genes were needed for accurate qRT-PCR normalization for the experimental design of all samples (V7/8 = 0.163).
- For the different tissues and developmental stage subgroups, EsACT, Esβ-TUB2 were the most suitable reference genes, and the addition of the EsUBQ gene was necessary for a reliable normalization result (V3/4 = 0.168)
- For untreated plants or stress-treated samples, including NaCl, PEG, ABA, cold, heat, UV, oxidation, injury and metal treatments, EsEF and Esα-TUB were the most stable genes, and the combination of the two was sufficient for normalization (V2/3 = 0.147)
- For seedling tissues grown under controlled conditions, EsEF and Esβ-TUB2 were the most stable reference genes and were sufficient for normalization (V2/3 = 0.087). For adult plant tissues collected from the field, EsGAPDH and EsUBQ were the two most stable genes and are sufficient for normalization (V2/3 = 0.113)
- Comparing tissue of seedlings grown under controlled conditions and adult plants collected from the field, EsACT and Esβ-TUB2 were the most stable genes, but the addition of another two or more stable genes is recommended for better normalization (V4/5 = 0.186)
- For the different stages of germination, EsEF and EsACT were the most stable genes, and these two genes were sufficient for normalization (V2/3 = 0.05).
- The Es18S gene was found to be unsuitable as a reference gene in our analysis.
Acknowledgments
References
- Zhang, D.Y.; Ma, W.B.; Shi, X.; Wang, J.C.; Wang, X.Y. Distribution and bio-ecological characteristics of Eremosparton songoricum, a rare plant in Gurbantunggut desert. J. Desert Res 2008, 28, 430–436. [Google Scholar]
- Yin, L.K.; Tan, L.X.; Wang, B. Rare Endangered Endemic High Plants in Xinjiang of China; Xinjiang Science and Technology Press: Urumqi, China, 2006; pp. 074–075. [Google Scholar]
- Zhang, L.Y.; Hai, Y. Plant communities excluded in the book of “The vegetation and its utilization in Xinjiang”. Arid Land Geogr 2002, 25, 84–89. [Google Scholar]
- Xu, Z.S.; Chen, M.; Li, L.C.; Ma, Y.Z. Functions and application of the AP2/ERF transcription factor family in crop improvement. J. Integr. Plant Biol 2011, 53, 570–585. [Google Scholar]
- Agarwal, P.K.; Agarwal, P.; Reddy, M.K.; Sopory, S.K. Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep 2006, 25, 1263–1274. [Google Scholar]
- Chen, M.; Wang, Q.Y.; Cheng, X.G.; Xu, Z.S.; Li, L.C.; Ye, X.G.; Xia, L.Q.; Ma, Y.Z. GmDREB2, a soybean DRE-binding transcription factor, conferred drought and high-salt tolerance in transgenic plants. Biochem. Biophys. Res. Commun 2007, 353, 299–305. [Google Scholar]
- Liu, N.; Zhong, N.Q.; Wang, G.L.; Li, L.J.; Liu, X.L.; He, Y.K.; Xia, G.X. Cloning and functional characterization of PpDBF1 gene encoding a DRE-binding transcription factor from Physcomitrella patens. Planta 2007, 226, 827–838. [Google Scholar]
- Tang, M.J.; Sun, J.W.; Liu, Y.; Chen, F.; Shen, S.H. Isolation and functional characterization of the JcERF gene, a putative AP2/EREBP domain-containing transcription factor, in the woody oil plant Jatropha curcas. Plant Mol. Biol 2007, 63, 419–428. [Google Scholar]
- Peng, X.J.; Ma, X.Y.; Fan, W.H.; Su, M.; Cheng, L.Q.; Alam, I.; Lee, B.H.; Qi, Q.M.; Shen, S.H.; Liu, G.S. Improved drought and salt tolerance of Arabidopsis thaliana by transgenic expression of a novel DREB gene from Leymus chinensis. Plant Cell Rep 2011, 30, 1493–1502. [Google Scholar]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun 2005, 6, 279–284. [Google Scholar]
- Artico, S.; Nardeli, S.M.; Brilhante, O.; Grossi-de-Sa, M.F.; Alves-Ferreira, M. Identification and evaluation of new reference genes in Gossypium hirsutum for accurate normalization of real-time quantitative RT-PCR data. BMC Plant Biol 2010, 10. [Google Scholar] [CrossRef]
- Bustin, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol 2000, 25, 169–193. [Google Scholar]
- Nitsche, A.; Radonic, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun 2004, 313, 856–862. [Google Scholar]
- Schmidt, G.W.; Delaney, S.K. Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during development and abiotic stress. Mol. Genet. Genomics 2010, 283, 233–241. [Google Scholar]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol 2002, 3, research0034:1–research0034:11. [Google Scholar]
- Hu, R.B.; Fan, C.M.; Fu, Y.F. Reference gene selection in plant real-time quantitative reverse transcription PCR (in chinese). J. Agric. Sci. Tech 2009, 11, 30–36. [Google Scholar]
- Maroufi, A.; van Bockstaele, E.; de Loose, M. Validation of reference genes for gene expression analysis in chicory (Cichorium intybus) using quantitative real-time PCR. BMC Mol. Biol 2010, 11, 15. [Google Scholar]
- Silveira, E.D.; Alves-Ferreira, M.; Guimaraes, L.A.; da Silva, F.R.; Carneiro, V.T. Selection of reference genes for quantitative real-time PCR expression studies in the apomictic and sexual grass Brachiaria brizantha. BMC Plant Biol 2009, 9. [Google Scholar] [CrossRef]
- Exposito-Rodriguez, M.; Borges, A.A.; Borges-Perez, A.; Perez, J.A. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process. BMC Plant Biol 2008, 8. [Google Scholar] [CrossRef]
- Lovdal, T.; Lillo, C. Reference gene selection for quantitative real-time PCR normalization in tomato subjected to nitrogen, cold, and light stress. Anal. Biochem 2009, 387, 238–242. [Google Scholar]
- Jian, B.; Liu, B.; Bi, Y.R.; Hou, W.S.; Wu, C.X.; Han, T.F. Validation of internal control for gene expression study in soybean by quantitative real-time PCR. BMC Mol. Biol 2008, 9. [Google Scholar] [CrossRef]
- Boava, L.P.; Laia, M.L.; Jacob, T.R.; Dabbas, K.M.; Goncalves, J.F.; Ferro, J.A.; Ferro, M.I.; Furtado, E.L. Selection of endogenous genes for gene expression studies in Eucalyptus under biotic (Puccinia psidii) and abiotic (acibenzolar-S-methyl) stresses using RT-qPCR. BMC Res. Notes 2010, 3. [Google Scholar] [CrossRef]
- Weyrich, A.; Axtner, J.; Sommer, S. Selection and validation of reference genes for real-time RT-PCR studies in the non-model species Delomys sublineatus, an endemic Brazilian rodent. Biochem. Biophys. Res. Commun 2010, 392, 145–149. [Google Scholar]
- Shen, Y.M.; Li, Y.; Ye, F.; Wang, F.F.; Lu, W.G.; Xie, X. Identification of suitable reference genes for measurement of gene expression in human cervical tissues. Anal. Biochem 2010, 405, 224–229. [Google Scholar]
- Ponton, F.; Chapuis, M.P.; Pernice, M.; Sword, G.A.; Simpson, S.J. Evaluation of potential reference genes for reverse transcription-qPCR studies of physiological responses in Drosophila melanogaster. J. Insect. Physiol 2011, 57, 840–850. [Google Scholar]
- Demidenko, N.V.; Logacheva, M.D.; Penin, A.A. Selection and validation of reference genes for quantitative real-Time PCR in Buckwheat (Fagopyrum esculentum) based on transcriptome sequence data. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Lee, J.M.; Roche, J.R.; Donaghy, D.J.; Thrush, A.; Sathish, P. Validation of reference genes for quantitative RT-PCR studies of gene expression in perennial ryegrass (Lolium perenne L.). BMC Mol. Biol 2010, 11. [Google Scholar] [CrossRef]
- Hong, S.Y.; Seo, P.J.; Yang, M.S.; Xiang, F.N.; Park, C.M. Exploring valid reference genes for gene expression studies in Brachypodium distachyon by real-time PCR. BMC Plant Biol 2008, 8. [Google Scholar] [CrossRef]
- Yan, J.W.; Yuan, F.R.; Long, G.Y.; Qin, L.; Deng, Z.N. Selection of reference genes for quantitative real-time RT-PCR analysis in citrus. Mol. Biol. Rep. 2011, 39, 1831–1838. [Google Scholar]
- Wan, H.J.; Zhao, Z.G.; Qian, C.T.; Sui, Y.H.; Malik, A.A.; Chen, J.F. Selection of appropriate reference genes for gene expression studies by quantitative real-time polymerase chain reaction in cucumber. Anal. Biochem 2010, 399, 257–261. [Google Scholar]
- Tong, Z.G.; Gao, Z.H.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol 2009, 10. [Google Scholar] [CrossRef]
- Migocka, M.; Papierniak, A. Identification of suitable reference genes for studying gene expression in cucumber plants subjected to abiotic stress and growth regulators. Mol. Breeding 2010, 1–15. [Google Scholar]
- Carvalho, K.; de Campos, M.K.F.; Pereira, L.F.P.; Vieira, L.G.E. Reference gene selection for real-time quantitative polymerase chain reaction normalization in “Swingle” citrumelo under drought stress. Anal. Biochem 2010, 402, 197–199. [Google Scholar]
- Zhong, H.Y.; Chen, J.W.; Li, C.Q.; Chen, L.; Wu, J.Y.; Chen, J.Y.; Lu, W.J.; Li, J.G. Selection of reliable reference genes for expression studies by reverse transcription quantitative real-time PCR in litchi under different experimental conditions. Plant Cell Rep 2011, 30, 641–653. [Google Scholar]
- Chen, L.; Zhong, H.Y.; Kuang, J.F.; Li, J.G.; Lu, W.J.; Chen, J.Y. Validation of reference genes for RT-qPCR studies of gene expression in banana fruit under different experimental conditions. Planta 2011, 234, 377–390. [Google Scholar]
- Gutierrez, N.; Gimenez, M.J.; Palomino, C.; Avila, C.M. Assessment of candidate reference genes for expression studies in Vicia faba L. by real-time quantitative PCR. Mol. Breeding 2011, 28, 13–24. [Google Scholar]
- Barsalobres-Cavallari, C.F.; Severino, F.E.; Maluf, M.P.; Maia, I.G. Identification of suitable internal control genes for expression studies in Coffea arabica under different experimental conditions. BMC Mol. Biol 2009, 10. [Google Scholar] [CrossRef]
- Cruz, F.; Kalaoun, S.; Nobile, P.; Colombo, C.; Almeida, J.; Barros, L.M.G.; Romano, E.; Grossi-de-Sa, M.F.; Vaslin, M.; Alves-Ferreira, M. Evaluation of coffee reference genes for relative expression studies by quantitative real-time RT-PCR. Mol. Breed 2009, 23, 607–616. [Google Scholar]
- Luo, H.L.; Chen, S.M.; Wan, H.J.; Chen, F.D.; Gu, C.S.; Liu, Z.L. Candidate reference genes for gene expression studies in water lily. Anal. Biochem 2010, 404, 100–102. [Google Scholar]
- Chen, X.; Truksa, M.; Shah, S.; Weselake, R.J. A survey of quantitative real-time polymerase chain reaction internal reference genes for expression studies in Brassica napus. Anal. Biochem 2010, 405, 138–140. [Google Scholar]
- Yang, Y.F.; Hou, S.; Cui, G.H.; Chen, S.L.; Wei, J.H.; Huang, L.Q. Characterization of reference genes for quantitative real-time PCR analysis in various tissues of Salvia miltiorrhiza. Mol. Biol. Rep 2010, 37, 507–513. [Google Scholar]
- Huis, R.; Hawkins, S.; Neutelings, G. Selection of reference genes for quantitative gene expression normalization in flax (Linum usitatissimum L.). BMC Plant Biol 2010, 10. [Google Scholar] [CrossRef]
- Gonzalez-Verdejo, C.I.; Die, J.V.; Nadal, S.; Jimenez-Marin, A.; Moreno, M.T.; Roman, B. Selection of housekeeping genes for normalization by real-time RT-PCR: Analysis of Or-MYB1 gene expression in Orobanche ramosa development. Anal. Biochem. 2008, 379, 176–181. [Google Scholar]
- Libault, M.; Thibivilliers, S.; Bilgin, D.D.; Radwan, O.; Benitez, M.; Clough, S.J.; Stacey, G. Identification of four soybean reference genes for gene expression normalization. Plant Genome 2008, 1. [Google Scholar] [CrossRef]
- Kakar, K.; Wandrey, M.; Czechowski, T.; Gaertner, T.; Scheible, W.R.; Stitt, M.; Torres-Jerez, I.; Xiao, Y.; Redman, J.C.; Wu, H.C.; et al. A community resource for high-throughput quantitative RT-PCR analysis of transcription factor gene expression in Medicago truncatula. Plant Methods 2008, 4. [Google Scholar] [CrossRef]
- Die, J.V.; Roman, B.; Nadal, S.; Gonzalez-Verdejo, C.I. Evaluation of candidate reference genes for expression studies in Pisum sativum under different experimental conditions. Planta 2010, 232, 145–153. [Google Scholar]
- Garg, R.; Sahoo, A.; Tyagi, A.K.; Jain, M. Validation of internal control genes for quantitative gene expression studies in chickpea (Cicer arietinum L.). Biochem. Biophys. Res. Commun 2010, 396, 283–288. [Google Scholar]
- Cordoba, E.M.; Die, J.V.; Gonzalez-Verdejo, C.I.; Nadal, S.; Roman, B. Selection of reference genes in Hedysarum coronarium under various stresses and stages of development. Anal.Biochem 2011, 409, 236–243. [Google Scholar]
- Li, H.P.; Qin, Y.X.; Xiao, X.; Tang, C. Screening of valid reference genes for real-time RT-PCR data normalization in Hevea brasiliensis and expression validation of a sucrose transporter gene HbSUT3. Plant Sci 2011, 181, 132–139. [Google Scholar]
- Lilly, S.T.; Drummond, R.S.M.; Pearson, M.N.; MacDiarmid, R.M. Identification and validation of reference genes for normalization of transcripts from virus-infected Arabidopsis thaliana. Mol. Plant Microbe In 2011, 24, 862–862. [Google Scholar]
- Chen, J.H.; Xia, X.L.; Yin, W.W. Expression profiling and functional characterization of a DREB2-type gene from Populus euphratica. Biochem. Biophys. Res. Commun 2009, 378, 483–487. [Google Scholar]
- Ahmad, S.; Ahmad, R.; Ashraf, M.Y.; Ashraf, M.; Waraich, E.A. Sunflower (Helianthus Annuus L.) response to drought stress at germination and seedling growth stages. Pak. J. Bot 2009, 41, 647–654. [Google Scholar]
- Yang, Y.F.; Wu, J.; Zhu, K.; Liu, L.Q.; Chen, F.D.; Yu, D.Y. Identification and characterization of two chrysanthemum (Dendronthema × moriforlium) DREB genes, belonging to the AP2/EREBP family. Mol Biol. Rep 2009, 36, 71–81. [Google Scholar]
- Luo, Z.Y.; Lu, Q.H.; Liu, S.P.; Chen, X.H.; Luo, J.Q.; Tan, L.J.; Hu, W.X. Screening and identification of novel genes involved in biosynthesis of ginsenoside in Panax ginseng plant. Acta Bioch. Bioph. Sin 2003, 35, 554–560. [Google Scholar]
- Chang, S.H.; Ying, J.; Zhang, J.J.; Su, J.Y.; Zeng, Y.J.; Tong, Y.P.; Li, B.; Li, Z.S. Expression of a wheat s-like RNAase (WRN1) cDNA during natural- and dark-induced senescence. Acta Bot. Sin 2003, 45, 1071–1075. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol 1990, 215, 403–410. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem 2009, 55, 611–622. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession number | Primer sequence (5′→3′, forward/reverse) | Length (bp) | Amplificatin efficiency (%) | R2 | Amplicon Tm (°C) |
|---|---|---|---|---|---|---|
| GAPDH | JN866814 | AATGTCTTTCCGTGTCC / TCCTCTTCAATGTAACCC | 150 | 93.5 | 0.993 | 81 |
| UBQ | JN866815 | TGGTCGCACCTTAGCCG / TCCCTCCTTATCTTGAATCTTGG | 178 | 91.5 | 0.999 | 82 |
| ACT | JN866816 | AGGAACCACCGATCCAGACA / GGTGCCCTGAGGTCCTGTT | 275 | 92.9 | 0.995 | 83 |
| EF | JN866817 | CGGACATCGTGACTTTATC / TGTGGTGGCATCCATCTT | 199 | 94.9 | 0.992 | 82.5 |
| β-TUB1 | JN866818 | ATTCCTTTCCCTCGTTTG / AATGTGGGATGCCAAGAA | 122 | 99.5 | 0.999 | 83 |
| β-TUB2 | JN866819 | TTACCTCACCGCCTCAG / AAGCCATCTTCAAACCT | 167 | 93 | 0.999 | 80.5 |
| α-TUB | JN866820 | TAGCAGCGTCTTCCTTT / ATGGTTTGATGCCGAGT | 204 | 98.9 | 0.997 | 84 |
| 18S | JN866821 | GGAGAGGGAGCCTGAGA / CACCAGACTTGCCCTCCAA | 188 | 98.4 | 1 | 83.5 |
| EsDREB | HQ687367 | GCTGCTCTTGCTTATGAT / TCTACCCCCGAGTTGTTT | 185 | 94.9 | 0.999 | 84.5 |
| Experiment sets | The three most stable genes | Optimal combination | The least stable gene |
|---|---|---|---|
| total | ACT, β-TUB1, β-TUB2 | V7/8 = 0.163 | 18S |
| tissues and developmental stage | ACT, β-TUB2, UBQ | ACT + β-TUB2 + UBQ (V3/4 = 0.168) | 18S |
| stresses | EF, α-TUB, UBQ | EF + α-TUB (V2/3 = 0.147) | GAPDH |
| tissues | ACT, β-TUB2, UBQ | (V4/5 = 0.186) | 18S |
| tissues (seedling) | EF, β-TUB2, ACT | EF + β-TUB2 (V2/3 = 0.087) | 18S |
| tissues (adult) | GAPDH, UBQ, β-TUB2 | GAPDH + UBQ (V2/3 = 0.113) | 18S |
| germination stage | EF, ACT, α-TUB | EF + ACT (V2/3 = 0.05) | GAPDH |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, X.-S.; Yang, H.-L.; Zhang, D.-Y.; Zhang, Y.-M.; Wood, A.J. Reference Gene Selection in the Desert Plant Eremosparton songoricum. Int. J. Mol. Sci. 2012, 13, 6944-6963. https://doi.org/10.3390/ijms13066944
Li X-S, Yang H-L, Zhang D-Y, Zhang Y-M, Wood AJ. Reference Gene Selection in the Desert Plant Eremosparton songoricum. International Journal of Molecular Sciences. 2012; 13(6):6944-6963. https://doi.org/10.3390/ijms13066944
Chicago/Turabian StyleLi, Xiao-Shuang, Hong-Lan Yang, Dao-Yuan Zhang, Yuan-Ming Zhang, and Andrew J. Wood. 2012. "Reference Gene Selection in the Desert Plant Eremosparton songoricum" International Journal of Molecular Sciences 13, no. 6: 6944-6963. https://doi.org/10.3390/ijms13066944