The Effect of Toll-Like Receptor 4 on Macrophage Cytokines During Endotoxin Induced Uveitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
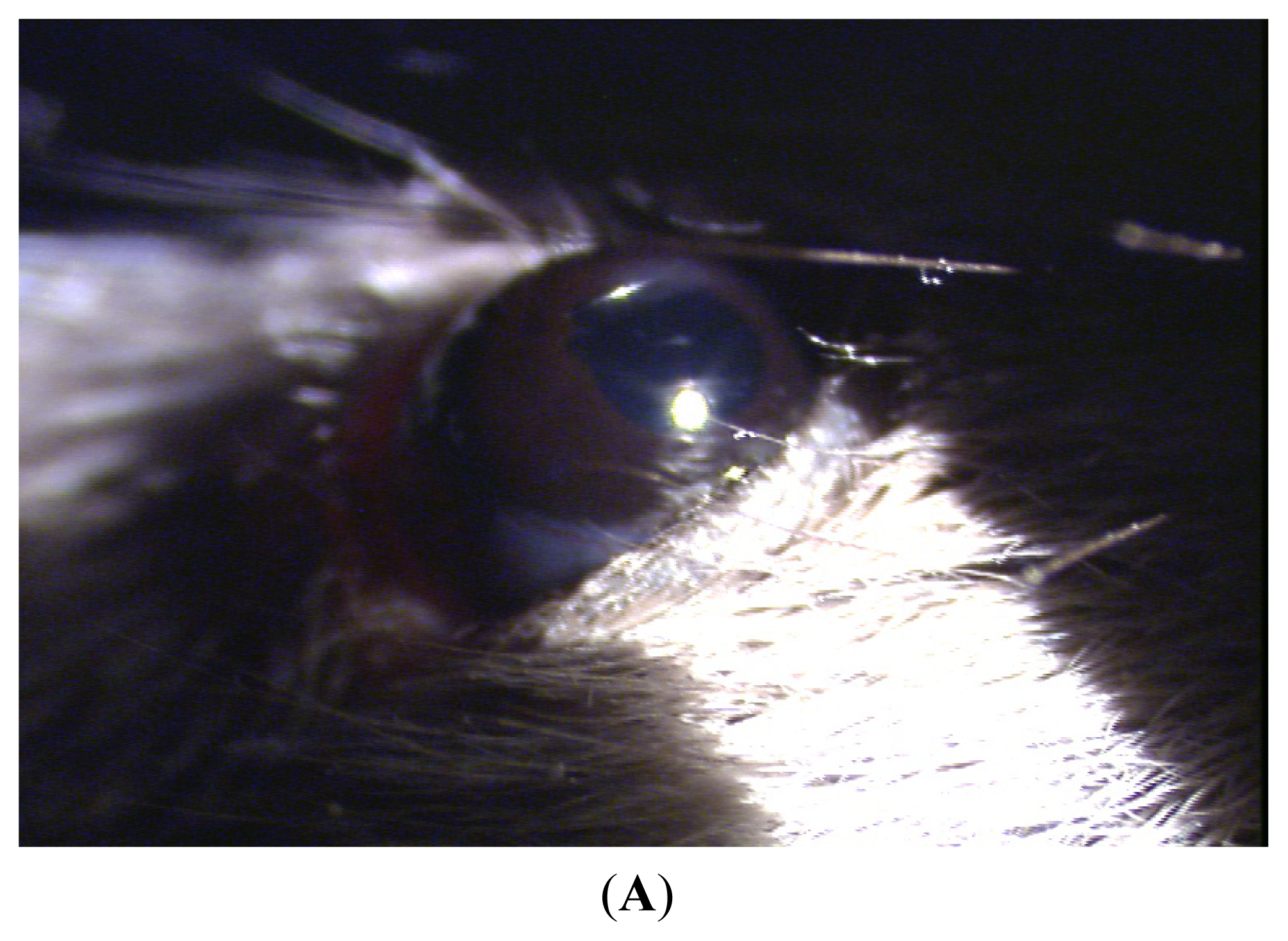
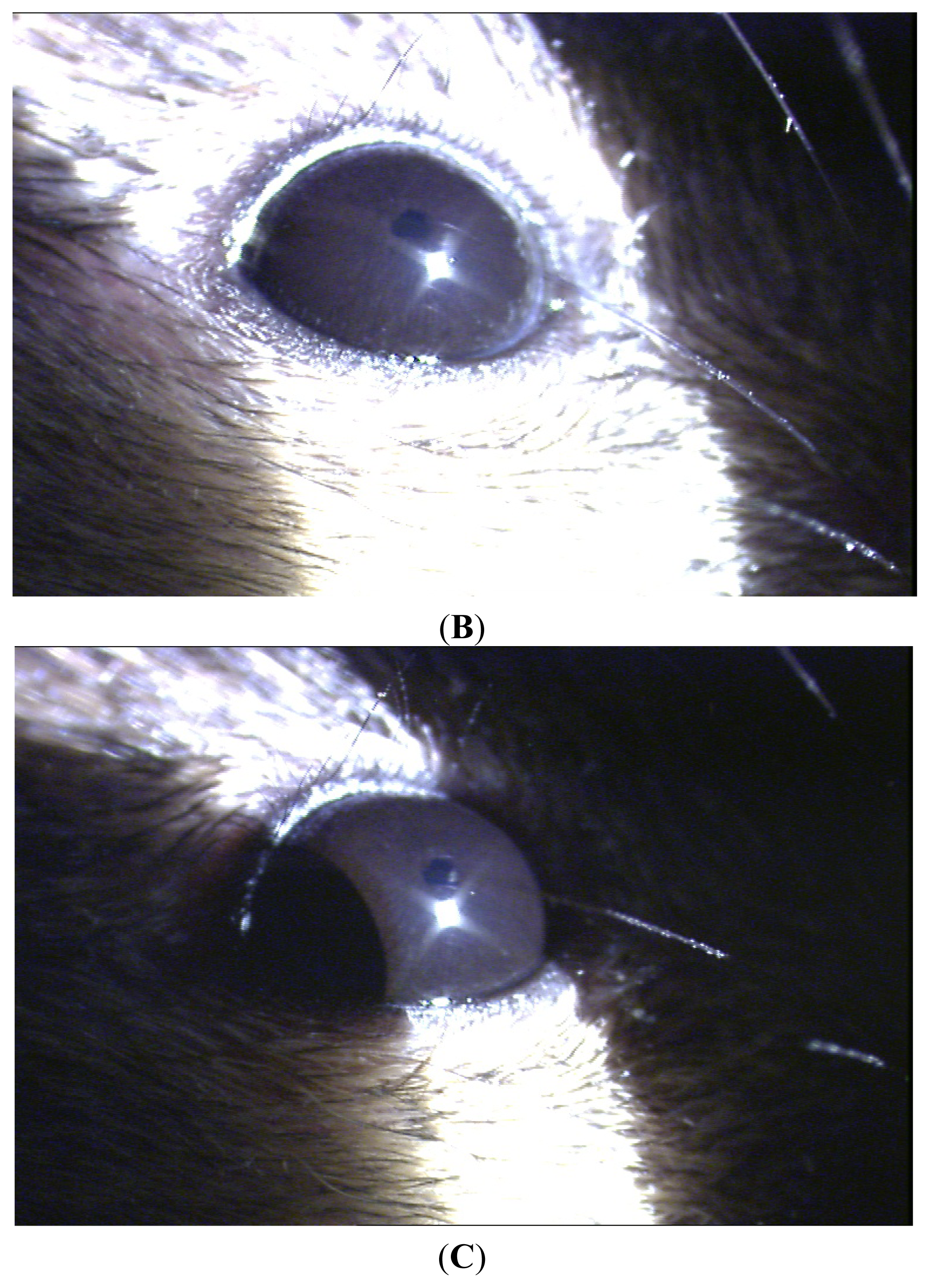
Clinical manifestation of endotoxin-induced uveitis (EIU)
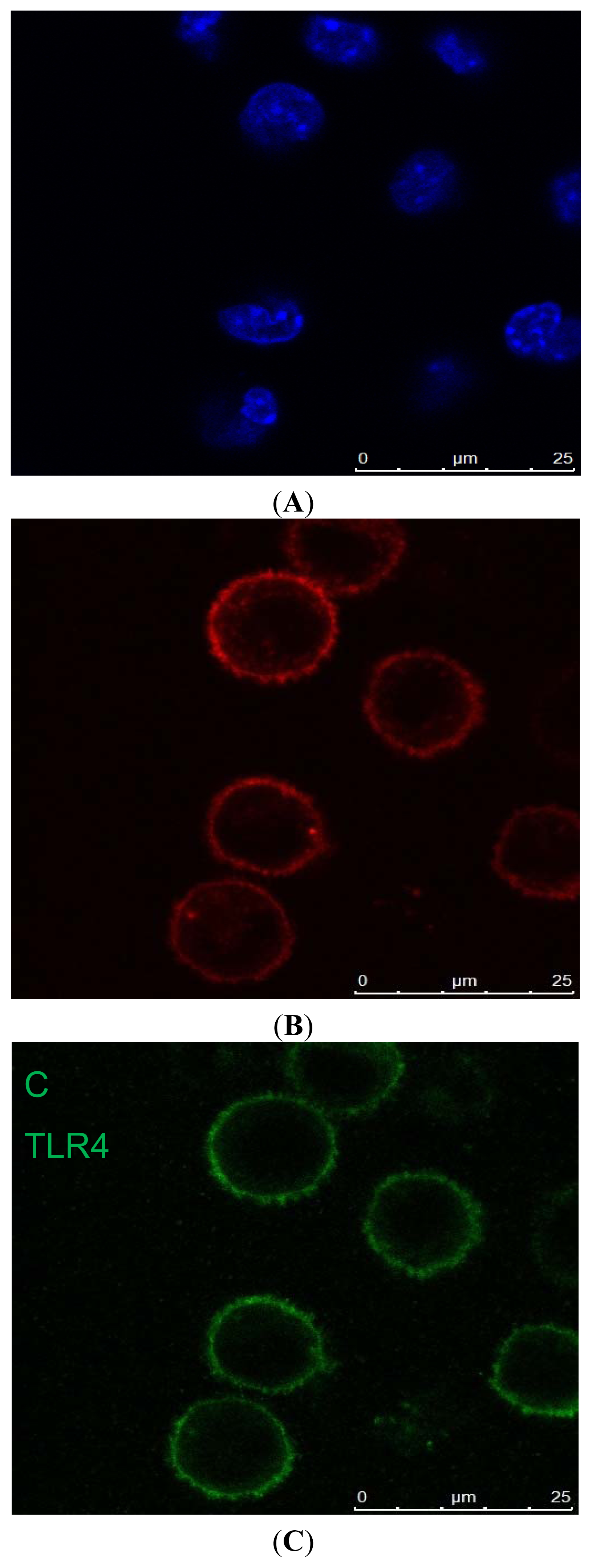
Cell Identification
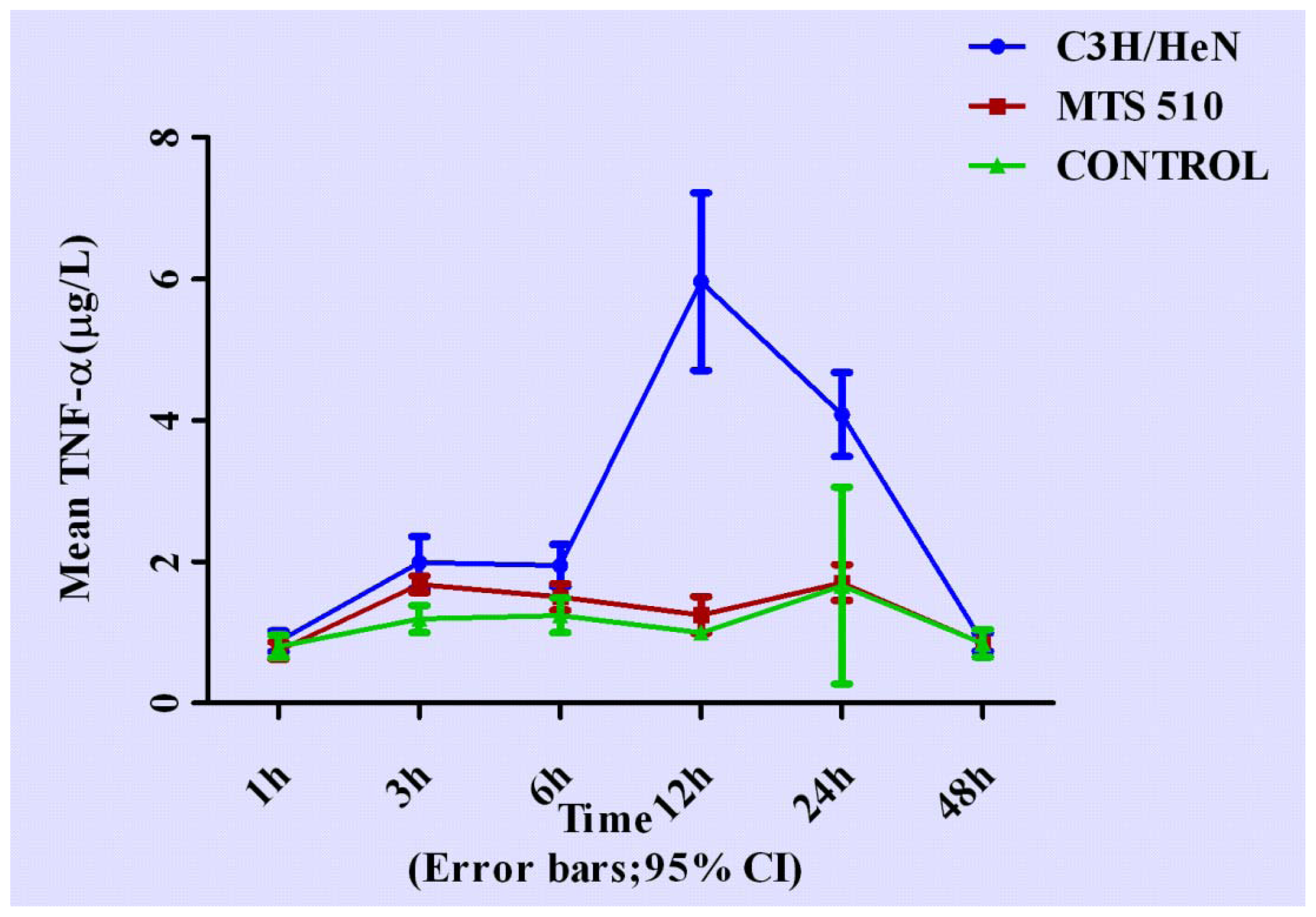
2.1. Concentration of Tumor Necrosis Factor-α in Groups of C3H/HeN, MTS 510 and CONTROL
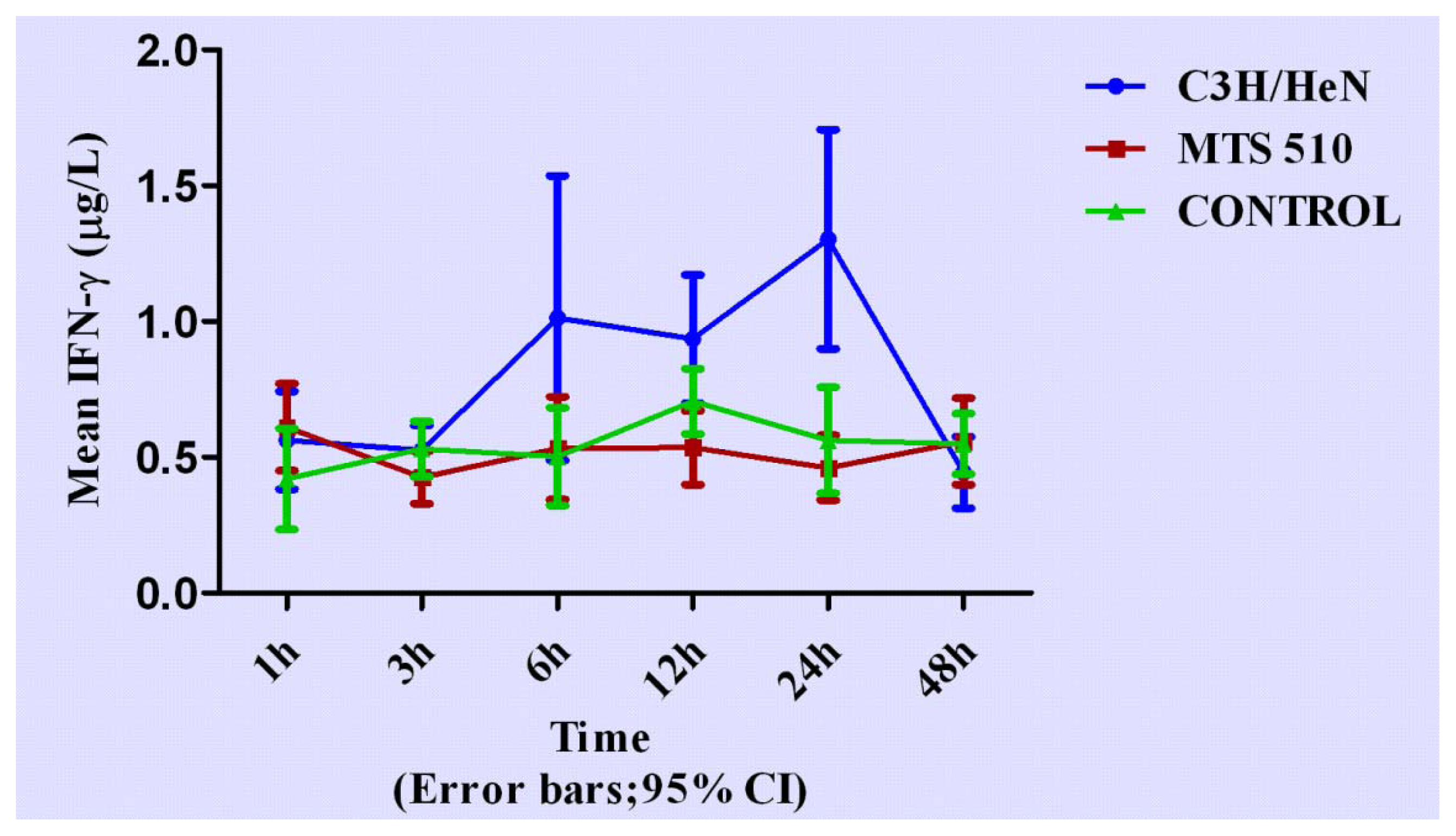
2.2. Concentration of Interferon-γ in Groups of C3H/HeN, MTS 510 and CONTROL
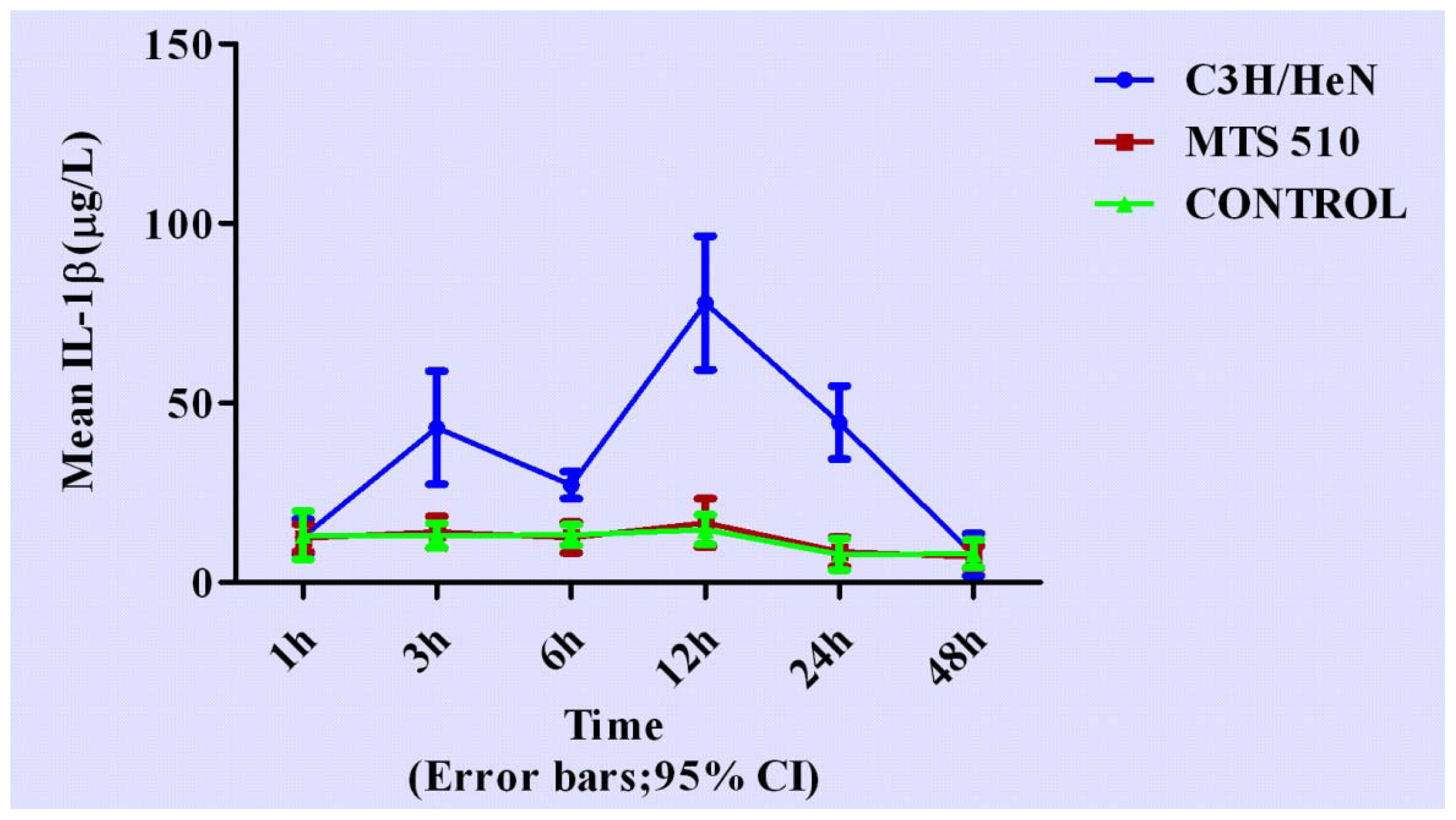
2.3. Concentration of Interleukin-1β in Groups of C3H/HeN, MTS 510 and CONTROL
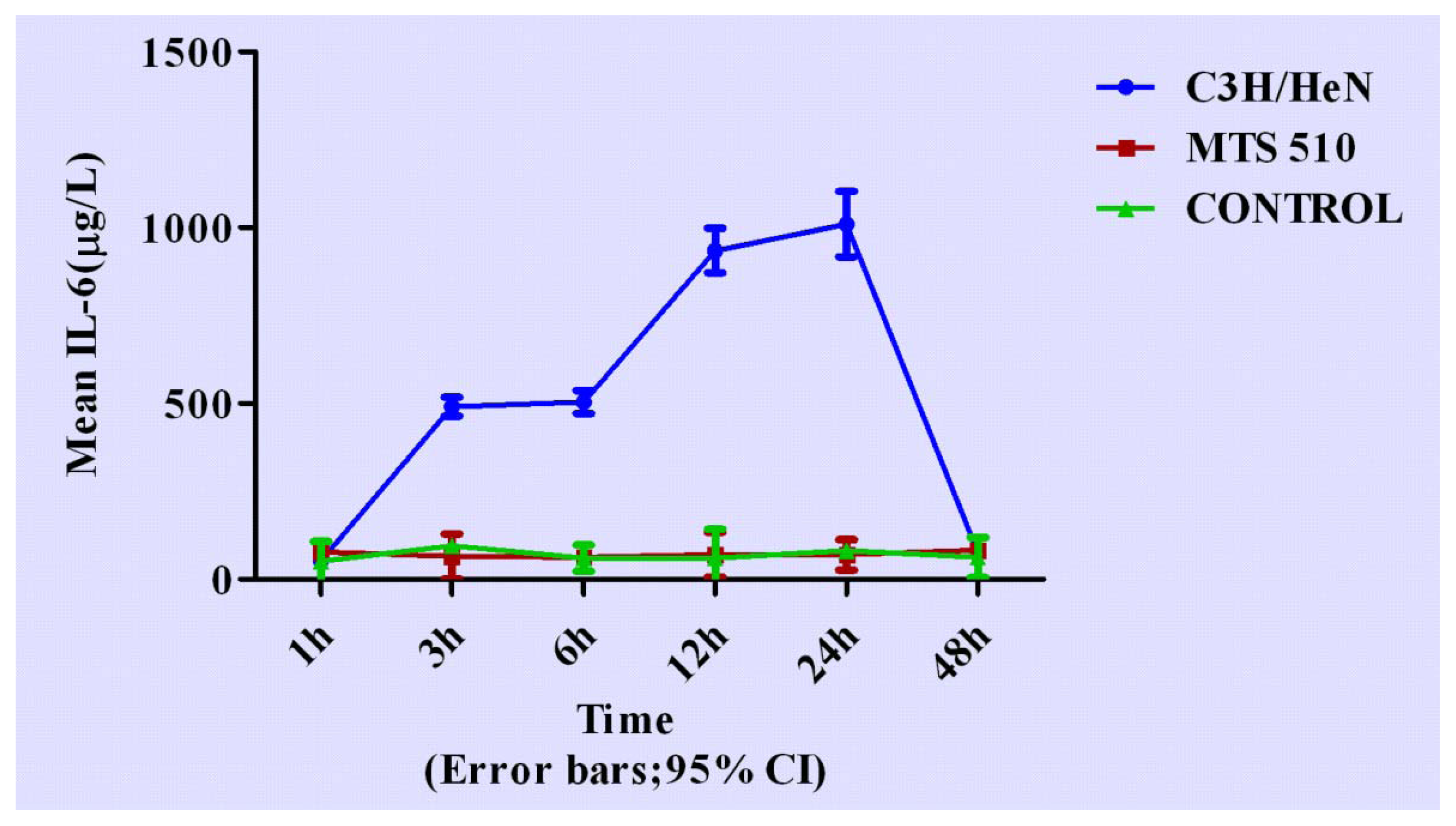
2.4. Concentration of Interleukin-6 in Groups of C3H/HeN, MTS 510 and CONTROL
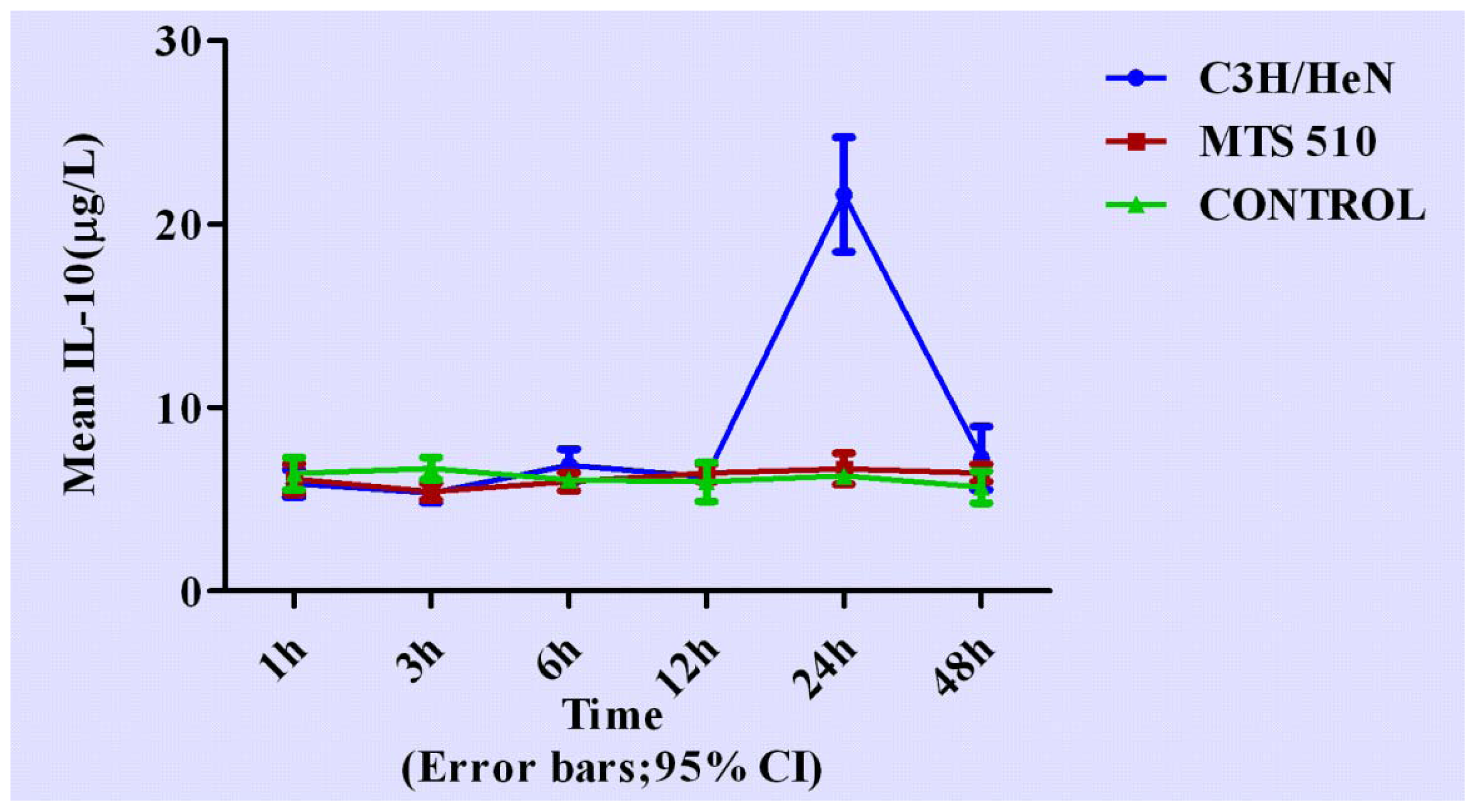
2.5. Concentration of Interleukin-10 in Groups of C3H/HeN, MTS 510 and CONTROL
3. Discussion
4. Experimental Section
4.1. Animals
4.2. Experimental Groups
4.3. Establishing the EIU Model
4.4. Culture and LPS Stimulation of Peritoneal Macrophages
4.5. Immunofluorescence
4.6. ELISA
4.7. Data Processing and Statistical Analysis
5. Conclusions
Acknowledgments
References
- Rathinam, S.R.; Namperumalsamy, P. Global variation and pattern changes in epidemiology of uveitis. Indian J. Ophthalmol 2007, 55, 173–183. [Google Scholar]
- Chang, J.H.; McCluskey, P.J.; Wakefield, D. Acute anterior uveitis and HLA-B27. Surv. Ophthalmol 2005, 50, 364–388. [Google Scholar]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol 2005, 17, 1–14. [Google Scholar]
- Chen, W.; Hu, X.; Zhao, L.; Li, S.; Lu, H. Expression of toll-like receptor 4 in uvea-resident tissue macrophages during endotoxin-induced uveitis. Mol. Vis 2009, 15, 619–628. [Google Scholar]
- McMenamin, P.G. Dendritic cells and macrophages in the uveal tract of the normal mouse eye. Br. J. Ophthalmol 1999, 83, 598–604. [Google Scholar]
- Xu, Y.Z.; Chen, W.; Hu, X.F.; Li, S.; Wang, J.; Zhao, L.; Lu, H. The expression of cytokines in the aqueous humor and serum during endotoxin-induced uveitis in C3H/HeN mice. Mol. Vis 2010, 16, 1689–1695. [Google Scholar]
- Verhasselt, V.; Buelens, C.; Willems, F.; de Groote, D.; Haeffner-Cavaillon, N.; Goldman, M. Bacterial lipopolysaccharide stimulates the production of cytokines and the expression of costimulatory molecules by human peripheral blood dendritic cells: Evidence for a soluble CD14-dependent pathway. J. Immunol 1997, 158, 2919–2925. [Google Scholar]
- Akashi, S.; Shimazu, R.; Ogata, H.; Nagai, Y.; Takeda, K.; Kimoto, M.; Miyake, K. Cell surface expression and lipopolysaccharide signaling via the Toll-like receptor 4-MD-2 complex on mouse peritoneal macrophages. J. Immunol 2000, 164, 3471–3475. [Google Scholar]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; van huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088. [Google Scholar]
- Zhang, X.J.; Li, Y.; Tai, G.X.; Xu, G.Y.; Zhang, P.Y.; Yang, Y.; Lao, F.X.; Liu, Z.H. Effects of activin A on the activities of mouse peritoneal macrophages. Cell. Mol. Immunol 2005, 2, 63–67. [Google Scholar]
- Drozina, G.; Kohoutek, J.; Nishiya, T.; Peterlin, B.M. Sequential modifications in class II transactivator isoform 1 induced by lipopolysaccharide stimulate major histocompatibility complex class II transcription in macrophages. J. Biol. Chem 2006, 281, 39963–39970. [Google Scholar]
- Crume, K.P.; Miller, J.H.; La Flamme, A.C. Peloruside A, an antimitotic agent, specifically decreases tumor necrosis factor-{alpha} production by lipopolysaccharide-stimulated murine macrophages. Exp. Biol. Med 2007, 232, 607–613. [Google Scholar]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol 1999, 162, 3749–3752. [Google Scholar]
- Akashi, S.; Shimazu, R.; Ogata, H.; Nagai, Y.; Takeda, K.; Kimoto, M. Cell surface expression and lipopolysaccharide signaling via the toll-like receptor 4-MD-2 complex on mouse peritoneal macrophages. J. Immunol 2000, 164, 3471–3475. [Google Scholar]
- Chen, W.; Hu, X.; Zhao, L.; Li, S.; Lu, H. Expression of toll-like receptor 4 in uvea-resident tissue macrophages during endotoxin-induced uveitis. Mol. Vis 2009, 15, 619–628. [Google Scholar]
- Montolio, M.; Biarnes, M.; Tellez, N.; Escoriza, J.; Soler, J.; Montanya, E. Interleukin-1β and inducible form of nitric oxide synthase expression in early syngeneic islet transplantation. J. Endocrinol 2007, 192, 169–177. [Google Scholar]
- Del Prete, G.; de Carli, M.; Almerigogna, F.; Giudizi, M.G.; Biagiotti, R.; Romagnani, S. Human IL-10 is produced by both type 1 helper (Th1) and type 2 helper (Th2) T cell clones and inhibits their antigen-specific proliferation and cytokine production. J. Immunol 1993, 150, 353–360. [Google Scholar]
- Stumpo, R.; Kauer, M.; Martin, S.; Kolb, H. Alternative activation of macrophages by IL-10. Pathobiology 1999, 67, 245–248. [Google Scholar]
- Schreiber, T.; Ehlers, S.; Heitmann, L.; Rausch, A.; Mages, J.; Murray, P.J. Autocrine IL-10 induces hallmarks of alternative activation in macrophages and suppresses antituberculosis effector mechanisms without compromising T cell immunity. J. Immunol 2009, 183, 1301–1312. [Google Scholar]
- Muzio, M.; Bosisio, D.; Polentarutti, N.; D’amico, G.; Stoppacciaro, A.; Mancinelli, R.; van’t Veer, C.; Penton-Rol, G.; Ruco, L.P.; Allavena, P.; et al. Differential expression and regulation of Toll-like recaptors (TLR) in human leukoeytes:selective expression of TLR3 in dendritic cells. J. Immunol 2000, 164, 5998–6004. [Google Scholar]
- Matta, B.; Jha, P.; Bora, P.S.; Bora, N.S. Tolerance to melanin-associated antigen in autoimmune uveitis is mediated by CD4+CD25+ T-regulatory cells. Am. J. Pathol 2008, 173, 1440–1454. [Google Scholar]








© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, S.; Lu, H.; Wang, J.; Qi, X.; Liu, X.; Zhang, X. The Effect of Toll-Like Receptor 4 on Macrophage Cytokines During Endotoxin Induced Uveitis. Int. J. Mol. Sci. 2012, 13, 7508-7520. https://doi.org/10.3390/ijms13067508
Yang S, Lu H, Wang J, Qi X, Liu X, Zhang X. The Effect of Toll-Like Receptor 4 on Macrophage Cytokines During Endotoxin Induced Uveitis. International Journal of Molecular Sciences. 2012; 13(6):7508-7520. https://doi.org/10.3390/ijms13067508
Chicago/Turabian StyleYang, Shuo, Hong Lu, Jing Wang, Xin Qi, Xuhui Liu, and Xiaolong Zhang. 2012. "The Effect of Toll-Like Receptor 4 on Macrophage Cytokines During Endotoxin Induced Uveitis" International Journal of Molecular Sciences 13, no. 6: 7508-7520. https://doi.org/10.3390/ijms13067508