l-2-Oxothiazolidine-4-Carboxylic Acid or α-Lipoic Acid Attenuates Airway Remodeling: Involvement of Nuclear Factor-κB (NF-κB), Nuclear Factor Erythroid 2p45-Related Factor-2 (Nrf2), and Hypoxia-Inducible Factor (HIF)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
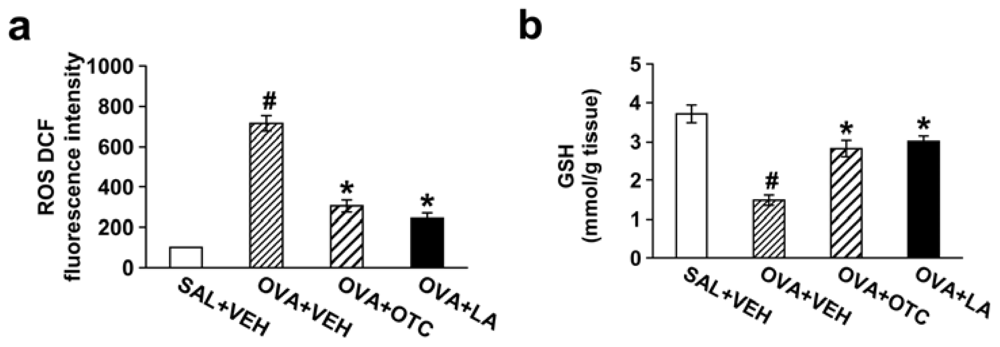
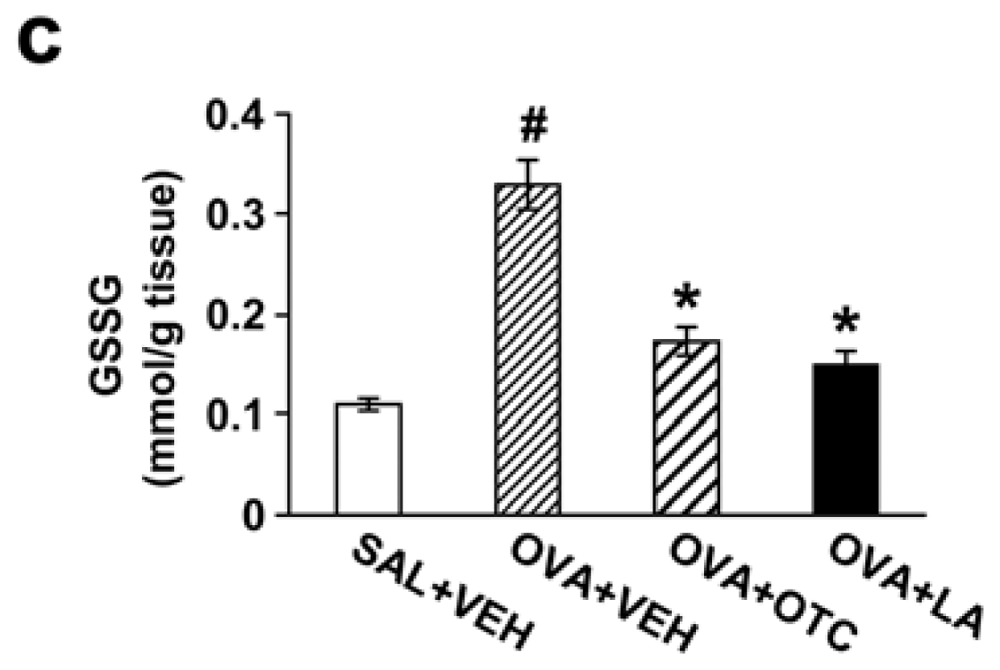
2.1. OTC or LA Decreases ROS Generation and Increases GSH Levels in the Lung
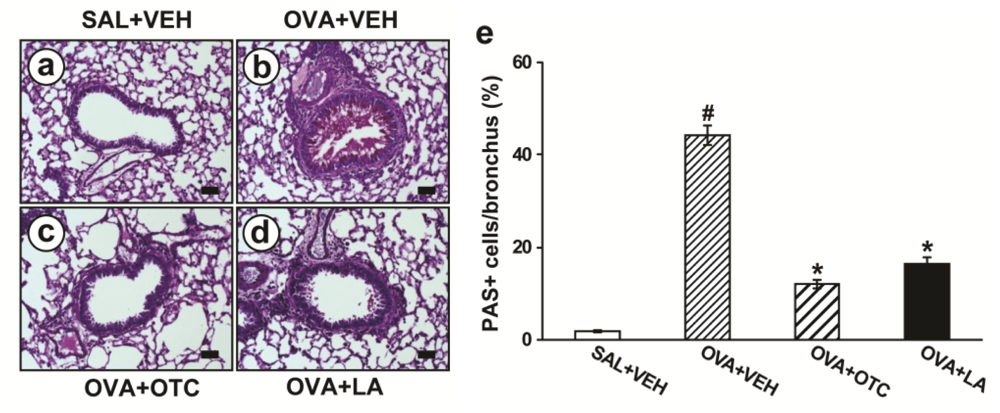
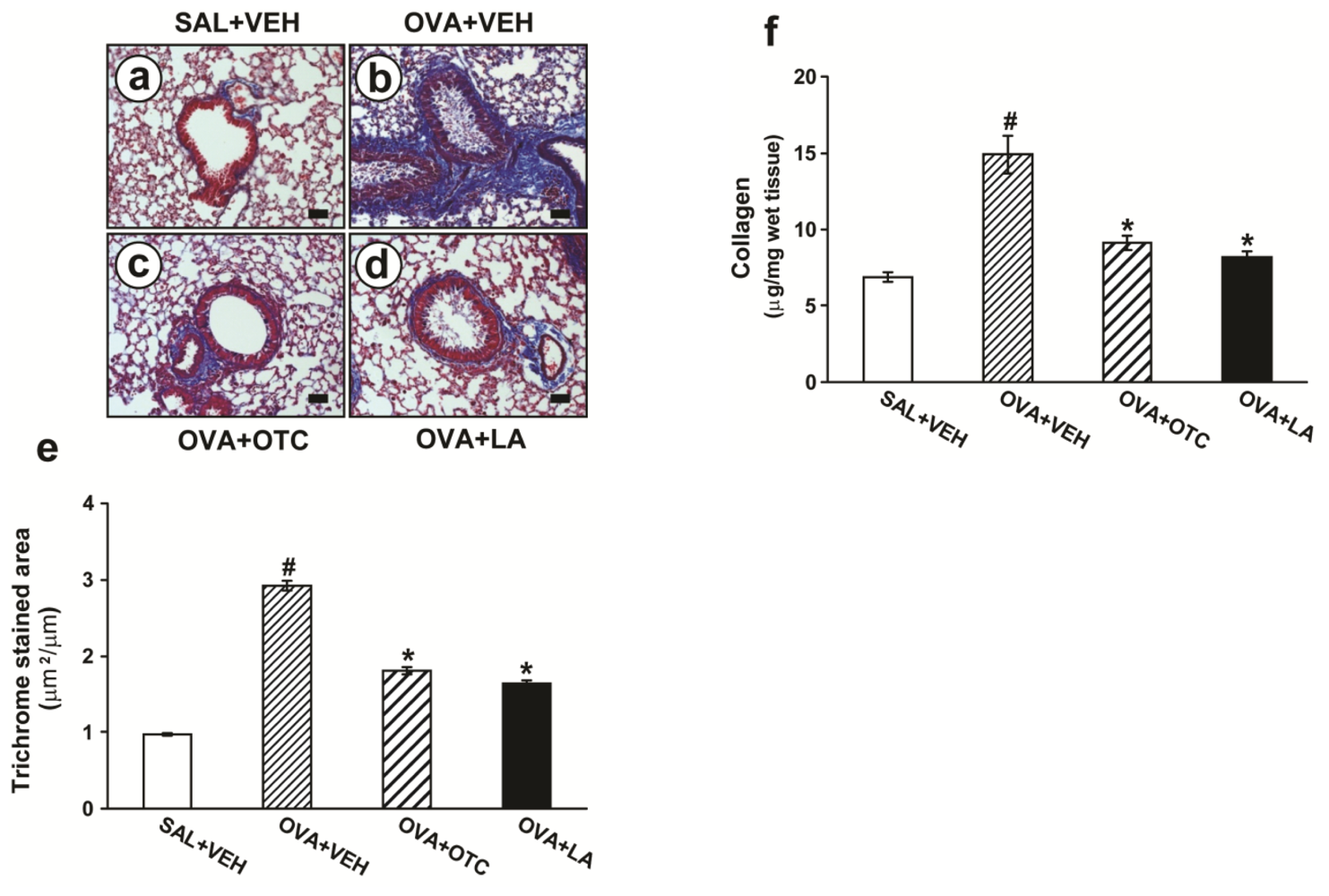
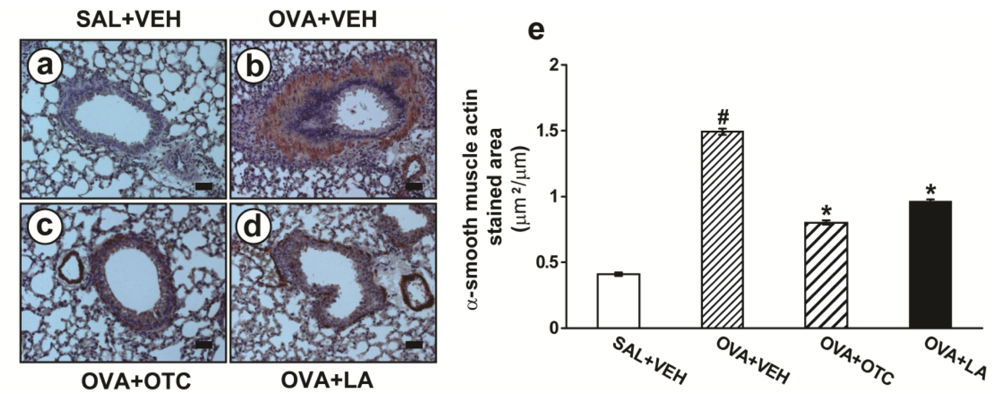
2.2. OTC and LA Inhibit Allergen-Induced Airway Remodeling
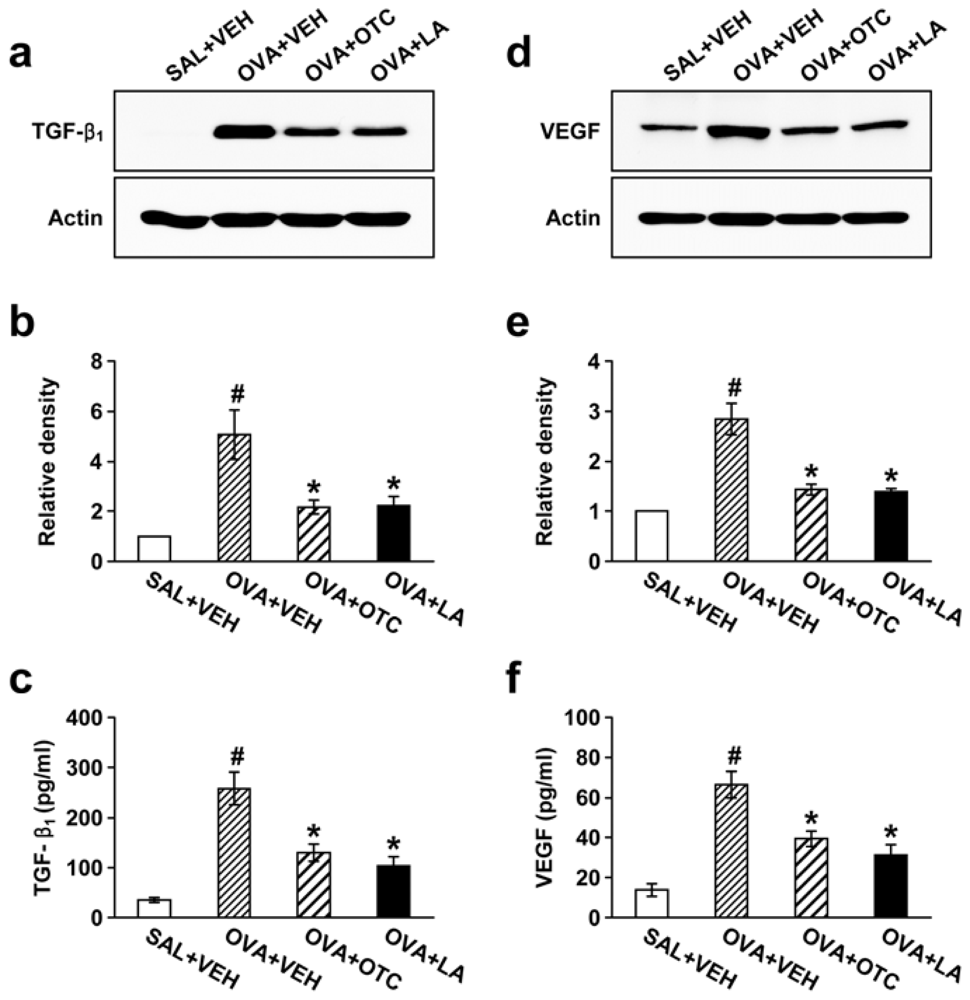
2.3. OTC and LA Suppress TGF-β1 Expression in the Lung
2.4. OTC and LA Inhibit OVA-Induced Vascular Endothelial Growth Factor (VEGF) Expression in the Lung
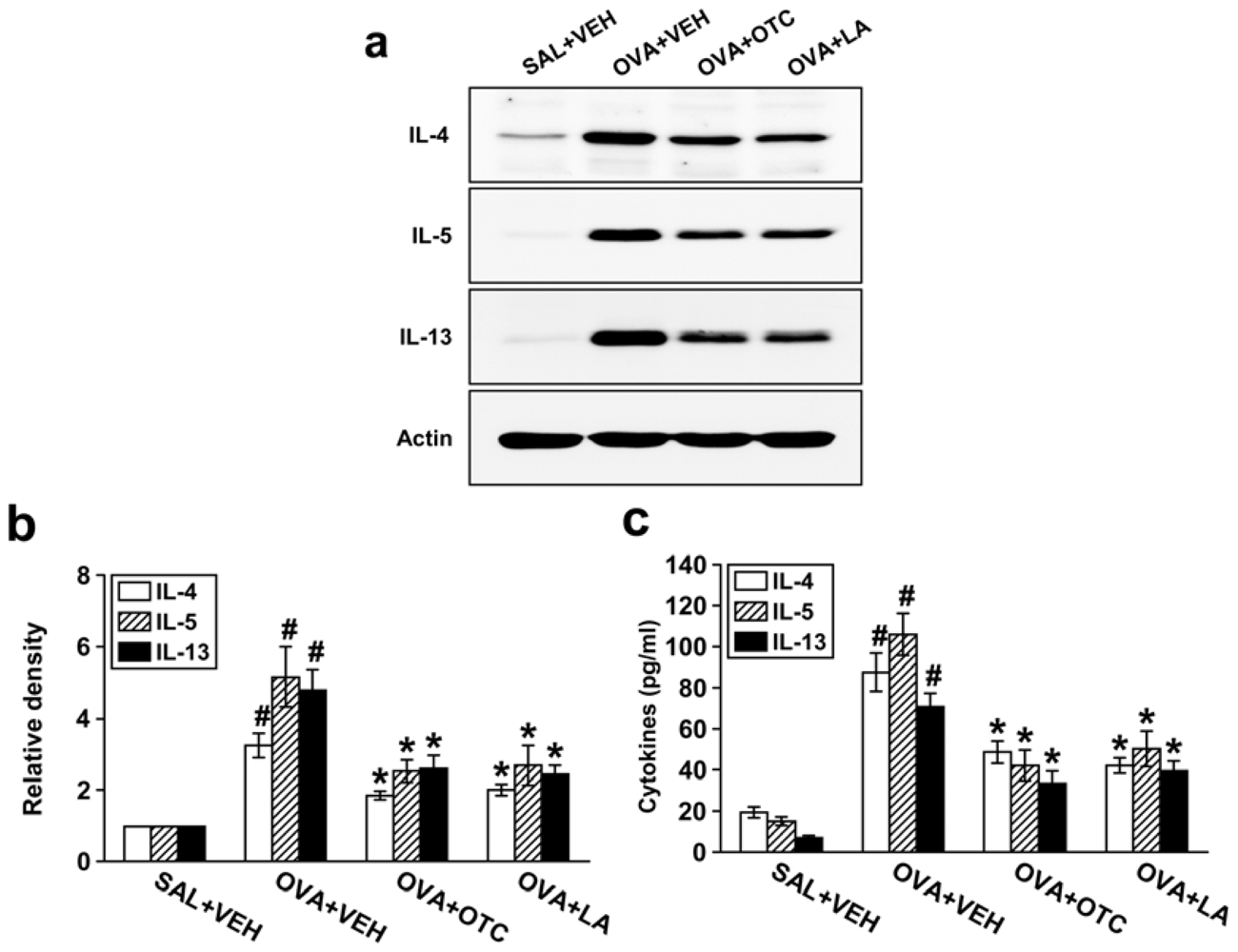
2.5. OTC and LA Suppress Expression of Th2 Cytokines
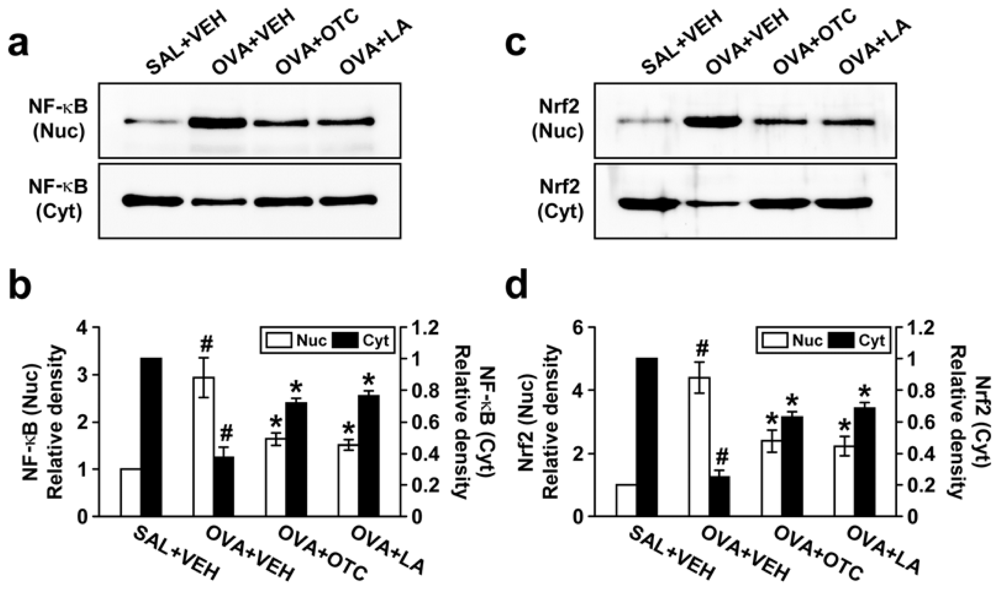
2.6. OTC and LA Negatively Modulate OVA-Induced Activation of NF-κB p65 and Nrf2
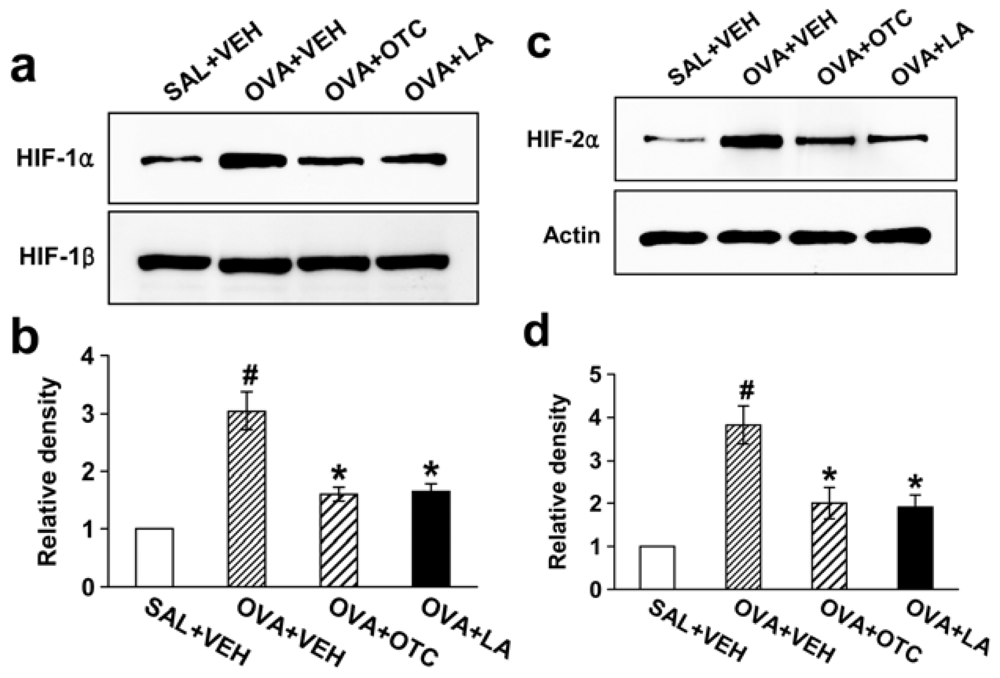
2.7. OTC and LA Decrease Levels of HIF-1α and HIF-2α
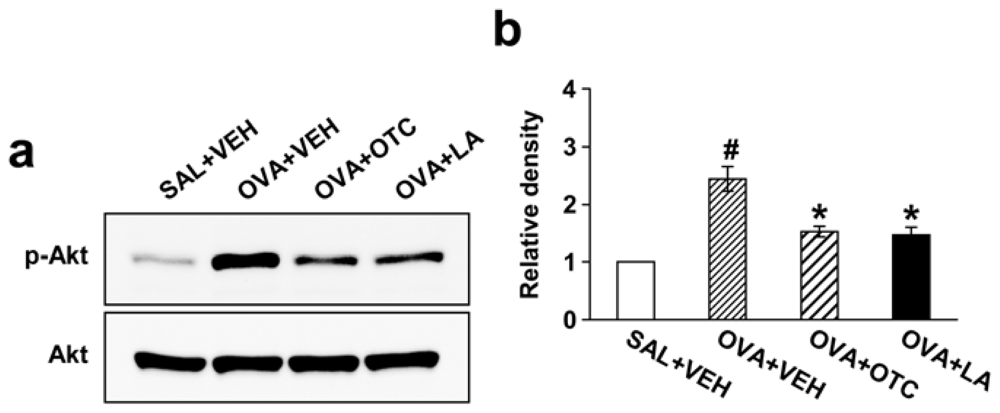
2.8. OTC and LA Down-Regulate OVA-Induced PI3K/Akt
2.9. OTC and LA Suppress Phosphorylation of p38 MAPK but Not Extracellular Signal-Regulated Kinase 1/2 (ERK1/2) and c-Jun N-Terminal Kinase (JNK)
2.10. OTC and LA Ameliorate Airway Inflammation and AHR in Chronic OVA-Challenged Mice
3. Discussion
4. Experimental Section
4.1. Animals and Experimental Protocol
4.2. Administration of OTC and LA
4.3. Measurement of Intracellular ROS
4.4. Measurement of GSH and GSSG in Lung Tissues
4.5. Western Blot Analysis
4.6. Cytosolic or Nuclear Protein Extractions for Analysis of NF-κB p65, Nrf2, HIF-1α, HIF-1β, and HIF-2α
4.7. Measurement of TGF-β1, VEGF, and Th2 Cytokines
4.8. Processing of Lungs for Histologic and Image Analysis
4.9. Quantitation of Airway Mucus Expression
4.10. Assessment of Peribronchial Fibrosis
Peribronchial trichrome staining
Determination of total lung collagen content
4.11. Quantitation of Peribronchial Smooth Muscle Layer Thickness
4.12. Determination of Airway Responsiveness
4.13. Densitometric Analyses and Statistics
5. Conclusions
Acknowledgments
References
- Chiappara, G.; Gagliardo, R.; Siena, A.; Bonsignore, M.R.; Bousquet, J.; Bonsignore, G.; Vignola, A.M. Airway remodelling in the pathogenesis of asthma. Curr. Opin. Allergy Clin. Immunol 2001, 1, 85–93. [Google Scholar]
- Park, S.J.; Lee, Y.C. Antioxidants as novel agents for asthma. Mini Rev. Med. Chem 2006, 6, 235–240. [Google Scholar]
- De Boer, W.I.; Yao, H.; Rahman, I. Future therapeutic treatment of COPD: Struggle between oxidants and cytokines. Int. J. Chron. Obstruct. Pulmon. Dis 2007, 2, 205–228. [Google Scholar]
- Comhair, S.A.; Erzurum, S.C. Redox control of asthma: Molecular mechanisms and therapeutic opportunities. Antioxid. Redox Signal 2010, 12, 93–124. [Google Scholar]
- Lee, Y.C.; Lee, K.S.; Park, S.J.; Park, H.S.; Lim, J.S.; Park, K.H.; Im, M.J.; Choi, I.W.; Lee, H.K.; Kim, U.H. Blockade of airway hyperresponsiveness and inflammation in a murine model of asthma by a prodrug of cysteine, L-2-oxothiazolidine-4-carboxylic acid. FASEB J 2004, 18, 1917–1919. [Google Scholar]
- Tanabe, T.; Fujimoto, K.; Yasuo, M.; Tsushima, K.; Yoshida, K.; Ise, H.; Yamaya, M. Modulation of mucus production by interleukin-13 receptor alpha2 in the human airway epithelium. Clin. Exp. Allergy 2008, 38, 122–134. [Google Scholar]
- Yasuo, M.; Fujimoto, K.; Tanabe, T.; Yaegashi, H.; Tsushima, K.; Takasuna, K.; Koike, T.; Yamaya, M.; Nikaido, T. Relationship between calcium-activated chloride channel 1 and MUC5AC in goblet cell hyperplasia induced by interleukin-13 in human bronchial epithelial cells. Respiration 2006, 73, 347–359. [Google Scholar]
- Cheng, S.E.; Lee, I.T.; Lin, C.C.; Kou, Y.R.; Yang, C.M. Cigarette smoke particle-phase extract induces HO-1 expression in human tracheal smooth muscle cells: Role of the c-Src/NADPH oxidase/MAPK/Nrf2 signaling pathway. Free Radic. Biol. Med 2010, 48, 1410–1422. [Google Scholar]
- Papaiahgari, S.; Zhang, Q.; Kleeberger, S.R.; Cho, H.Y.; Reddy, S.P. Hyperoxia stimulates an Nrf2-ARE transcriptional response via ROS-EGFR-PI3K-Akt/ERK MAP kinase signaling in pulmonary epithelial cells. Antioxid. Redox Signal 2006, 8, 43–52. [Google Scholar]
- Koshikawa, N.; Hayashi, J.; Nakagawara, A.; Takenaga, K. Reactive oxygen species-generating mitochondrial DNA mutation up-regulates hypoxia-inducible factor-1alpha gene transcription via phosphatidylinositol 3-kinase-Akt/protein kinase C/histone deacetylase pathway. J. Biol. Chem 2009, 284, 33185–33194. [Google Scholar]
- Polosukhin, V.V.; Cates, J.M.; Lawson, W.E.; Milstone, A.P.; Matafonov, A.G.; Massion, P.P.; Lee, J.W.; Randell, S.H.; Blackwell, T.S. Hypoxia-inducible factor-1 signalling promotes goblet cell hyperplasia in airway epithelium. J. Pathol 2011, 224, 203–211. [Google Scholar]
- Jiang, H.; Zhu, Y.S.; Xu, H.; Sun, Y.; Li, Q.F. Inflammatory stimulation and hypoxia cooperatively activate HIF-1α in bronchial epithelial cells: Involvement of PI3K and NF-κB. Am. J. Physiol. Lung Cell. Mol. Physiol 2010, 298, L660–L669. [Google Scholar]
- Vita, J.A.; Frei, B.; Holbrook, M.; Gokce, N.; Leaf, C.; Keaney, J.F. L-2-Oxothiazolidine-4-carboxylic acid reverses endothelial dysfunction in patients with coronary artery disease. J. Clin. Invest 1998, 101, 1408–1414. [Google Scholar]
- Packer, L.; Witt, E.H.; Tritschler, H.J. Alpha-lipoic acid as a biological antioxidant. Free Radic. Biol. Med 1995, 19, 227–250. [Google Scholar]
- Lee, K.S.; Kim, S.R.; Park, S.J.; Min, K.H.; Lee, K.Y.; Jin, S.M.; Yoo, W.H.; Lee, Y.C. Antioxidant down-regulates IL-18 expression in asthma. Mol. Pharmacol 2006, 70, 1184–1193. [Google Scholar]
- Fattouh, R.; Jordana, M. TGF-beta, eosinophils and IL-13 in allergic airway remodeling: A critical appraisal with therapeutic considerations. Inflamm. Allergy Drug Targets 2008, 7, 224–236. [Google Scholar]
- Siddiqui, S.; Sutcliffe, A.; Shikotra, A.; Woodman, L.; Doe, C.; McKenna, S.; Wardlaw, A.; Bradding, P.; Pavord, I.; Brightling, C. Vascular remodeling is a feature of asthma and nonasthmatic eosinophilic bronchitis. J. Allergy Clin. Immunol 2007, 120, 813–819. [Google Scholar]
- Doherty, T.; Broide, D. Cytokines and growth factors in airway remodeling in asthma. Curr. Opin. Immunol 2007, 19, 676–680. [Google Scholar]
- Huerta-Yepez, S.; Baay-Guzman, G.J.; Garcia-Zepeda, R.; Hernandez-Pando, R.; Vega, M.I.; Gonzalez-Bonilla, C.; Bonavida, B. 2-Methoxyestradiol (2-ME) reduces the airway inflammation and remodeling in an experimental mouse model. Clin. Immunol 2008, 129, 313–324. [Google Scholar]
- Liang, K.C.; Lee, C.W.; Lin, W.N.; Lin, C.C.; Wu, C.B.; Luo, S.F.; Yang, C.M. Interleukin-1beta induces MMP-9 expression via p42/p44 MAPK, p38 MAPK, JNK, and nuclear factor-kappaB signaling pathways in human tracheal smooth muscle cells. J. Cell. Physiol 2007, 211, 759–770. [Google Scholar]
- Hulsmann, A.R.; Raatgeep, H.R.; den Hollander, J.C.; Stijnen, T.; Saxena, P.R.; Kerrebijn, K.F.; de Jongste, J.C. Oxidative epithelial damage produces hyperresponsiveness of human peripheral airways. Am. J. Respir. Crit. Care Med 1994, 149, 519–525. [Google Scholar]
- Chaudiere, J.; Ferrari-Iliou, R. Intracellular antioxidants from chemical to biochemical mechanisms. Food Chem. Toxicol 1999, 37, 949–962. [Google Scholar]
- Heffner, J.A.; Repine, J.E. State of the art: Pulmonary strategies of antioxidant defense. Am. Rev. Respir. Dis 1989, 140, 531–554. [Google Scholar]
- Toth, K.M.; Clifford, D.P.; Berger, E.M.; White, C.W.; Repine, J.E. Intact human erythrocyte prevent hydrogen peroxide mediated damage to isolated perfused rat lungs and cultured bovine pulmonary artery endothelial cells. J. Clin. Invest 1984, 74, 292–295. [Google Scholar]
- Van Asbeck, B.S.; Hoidal, J.; Vercelloti, G.M.; Schwartz, B.A.; Moldow, C.F.; Jacob, H.S. Protection against lethal hyperoxia by tracheal insufflation of erythrocytes: Role of red cell glutathione. Science 1985, 227, 756–759. [Google Scholar]
- Comhair, S.A.; Xu, W.; Ghosh, S.; Thunnissen, F.B.; Almasan, A.; Calhoun, W.J.; Janocha, A.J.; Zheng, L.; Hazen, S.L.; Erzurum, S.C. Superoxide dismutase inactivation in pathophysiology of asthmatic airway remodeling and reactivity. Am. J. Pathol 2005, 166, 663–674. [Google Scholar]
- Imaoka, H.; Hoshino, T.; Okamoto, M.; Sakazaki, Y.; Sawada, M.; Takei, S.; Kinoshita, T.; Kawayama, T.; Kato, S.; Aizawa, H. Endogenous and exogenous thioredoxin 1 prevents goblet cell hyperplasia in a chronic antigen exposure asthma model. Allergol. Int 2009, 58, 403–410. [Google Scholar]
- Voynow, J.A.; Fischer, B.M.; Malarkey, D.E.; Burch, L.H.; Wong, T.; Longphre, M.; Ho, S.B.; Foster, W.M. Neutrophil elastase induces mucus cell metaplasia in mouse lung. Am. J. Physiol. Lung Cell. Mol. Physiol 2004, 287, L1293–L1302. [Google Scholar]
- De Marco, V.G.; Habibi, J.; Whaley-Connell, A.T.; Schneider, R.I.; Sowers, J.R.; Andresen, B.T.; Gutweiler, A.A.; Ma, L.; Johnson, M.S.; Ferrario, C.M.; et al. Rosuvastatin ameliorates the development of pulmonary arterial hypertension in the transgenic (mRen2)27 rat. Am. J. Physiol. Heart Circ. Physiol 2009, 297, H1128–H1139. [Google Scholar]
- Koli, K.; Myllärniemi, M.; Keski-Oja, J.; Kinnula, V.L. Transforming growth factor-beta activation in the lung: Focus on fibrosis and reactive oxygen species. Antioxid. Redox Signal 2008, 10, 333–342. [Google Scholar]
- Cho, J.Y.; Miller, M.; Baek, K.J.; Han, J.W.; Nayar, J.; Lee, S.Y.; McElwain, K.; McElwain, S.; Friedman, S.; Broide, D.H. Inhibition of airway remodeling in IL-5-deficient mice. J. Clin. Invest 2004, 113, 551–560. [Google Scholar]
- Lee, K.S.; Park, S.J.; Kim, S.R.; Min, K.H.; Lee, K.Y.; Choe, Y.H.; Hong, S.H.; Lee, Y.R.; Kim, J.S.; Hong, S.J.; Lee, Y.C. Inhibition of VEGF blocks TGF-beta1 production through a PI3K/Akt signalling pathway. Eur. Respir. J 2008, 31, 523–531. [Google Scholar]
- Cho, H.Y.; Kleeberger, S.R. Nrf2 protects against airway disorders. Toxicol. Appl. Pharmacol 2010, 244, 43–56. [Google Scholar]
- Semenza, G.L. Regulation of mammalian O2 homeostasis by hypoxia-inducible factor 1. Annu. Rev. Cell Dev. Biol 1999, 15, 551–578. [Google Scholar]
- Wiesener, M.S.; Jürgensen, J.S.; Rosenberger, C.; Scholze, C.K.; Hörstrup, J.H.; Warnecke, C.; Mandriota, S.; Bechmann, I.; Frei, U.A.; Pugh, C.W.; et al. Widespread hypoxia-inducible expression of HIF-2alpha in distinct cell populations of different organs. FASEB J 2003, 17, 271–273. [Google Scholar]
- Lambert, C.M.; Roy, M.; Robitaille, G.A.; Richard, D.E.; Bonnet, S. HIF-1 inhibition decreases systemic vascular remodelling diseases by promoting apoptosis through a hexokinase 2-dependent mechanism. Cardiovasc. Res 2010, 88, 196–204. [Google Scholar]
- Patten, D.A.; Lafleur, V.N.; Robitaille, G.A.; Chan, D.A.; Giaccia, A.J.; Richard, D.E. Hypoxia-inducible factor-1 activation in nonhypoxic conditions: The essential role of mitochondrial-derived reactive oxygen species. Mol. Biol. Cell 2010, 21, 3247–3257. [Google Scholar]
- Liu, J.; Yoshida, Y.; Yamashita, U. DNA-binding activity of NF-kappaB and phosphorylation of p65 are induced by N-acetylcysteine through phosphatidylinositol (PI) 3-kinase. Mol. Immunol 2008, 45, 3984–3989. [Google Scholar]
- Park, S.J.; Lee, K.S.; Kim, S.R.; Min, K.H.; Moon, H.; Lee, M.H.; Chung, C.R.; Han, H.J.; Puri, K.D.; Lee, Y.C. Phosphoinositide 3-kinase δ inhibitor suppresses IL-17 expression in a murine asthma model. Eur. Respir. J 2010, 36, 1448–1459. [Google Scholar]
- Boutten, A.; Goven, D.; Boczkowski, J.; Bonay, M. Oxidative stress targets in pulmonary emphysema: Focus on the Nrf2 pathway. Expert Opin. Ther. Targets 2010, 14, 329–346. [Google Scholar]
- Minet, E.; Arnould, T.; Michel, G.; Roland, I.; Mottet, D.; Raes, M.; Remacle, J.; Michiels, C. ERK activation upon hypoxia: Involvement in HIF-1 activation. FEBS Lett 2000, 468, 53–58. [Google Scholar]
- Sang, N.; Stiehl, D.P.; Bohensky, J.; Leshchinsky, I.; Srinivas, V.; Caro, J. MAPK signaling upregulates the activity of hypoxia-inducible factors by its effects on p300. J. Biol. Chem 2003, 278, 14013–14019. [Google Scholar]
- Richard, D.E.; Berra, E.; Gothie, E.; Roux, D.; Pouysségur, J. p42/p44 mitogen-activated protein kinases phosphorylate hypoxia-inducible factor 1α (HIF-1α) and enhance the transcriptional activity of HIF-1. J. Biol. Chem 1999, 274, 32631–32637. [Google Scholar]
- Lu, Y.; Azad, N.; Wang, L.; Iyer, A.K.; Castranova, V.; Jiang, B.H.; Rojanasakul, Y. Phosphatidylinositol-3-kinase/akt regulates bleomycin-induced fibroblast proliferation and collagen production. Am. J. Respir. Cell Mol. Biol 2010, 42, 432–441. [Google Scholar]
- Lee, K.S.; Kim, S.R.; Park, S.J.; Min, K.H.; Lee, K.Y.; Choe, Y.H.; Park, S.Y.; Chai, O.H.; Zhang, X.; Song, C.H.; Lee, Y.C. Mast cells can mediate vascular permeability through regulation of the PI3K-HIF-1alpha-VEGF axis. Am. J. Respir. Crit. Care Med 2008, 178, 787–797. [Google Scholar]
- Institute of Laboratory Animal Research, Guide for the Care and Use of Laboratory Animals; National Acamedy Press: Washington, DC, USA, 1996.
- Kwak, Y.G.; Song, C.H.; Yi, H.K.; Hwang, P.H.; Kim, J.S.; Lee, K.S.; Lee, Y.C. Involvement of PTEN in airway hyperresponsiveness and inflammation in bronchial asthma. J. Clin. Invest 2003, 111, 1083–1092. [Google Scholar]
- Royce, S.G.; Wainewright, J.S.; Samuel, C.S.; Tang, M.L. Comparison of airway remodeling in acute, subacute, and chronic models of allergic airways disease. Am. J. Respir. Cell Mol. Biol 2007, 36, 625–632. [Google Scholar]
- Lee, K.S.; Kim, S.R.; Park, S.J.; Park, H.S.; Min, K.H.; Jin, S.M.; Lee, M.K.; Kim, U.H.; Lee, Y.C. Peroxisome proliferator activated receptor-gamma modulates reactive oxygen species generation and activation of nuclear factor-kappaB and hypoxia-inducible factor 1alpha in allergic airway disease of mice. J. Allergy Clin. Immunol 2006, 118, 120–127. [Google Scholar]
- Lee, K.S.; Park, S.J.; Kim, S.R.; Min, K.H.; Jin, S.M.; Lee, H.K.; Lee, Y.C. Modulation of airway remodeling and airway inflammation by peroxisome proliferator-activated receptor gamma in a murine model of toluene diisocyanate-induced asthma. J. Immunol 2006, 177, 5248–5257. [Google Scholar]













© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Park, S.J.; Lee, K.S.; Lee, S.J.; Kim, S.R.; Park, S.Y.; Jeon, M.S.; Lee, H.B.; Lee, Y.C. l-2-Oxothiazolidine-4-Carboxylic Acid or α-Lipoic Acid Attenuates Airway Remodeling: Involvement of Nuclear Factor-κB (NF-κB), Nuclear Factor Erythroid 2p45-Related Factor-2 (Nrf2), and Hypoxia-Inducible Factor (HIF). Int. J. Mol. Sci. 2012, 13, 7915-7937. https://doi.org/10.3390/ijms13077915
Park SJ, Lee KS, Lee SJ, Kim SR, Park SY, Jeon MS, Lee HB, Lee YC. l-2-Oxothiazolidine-4-Carboxylic Acid or α-Lipoic Acid Attenuates Airway Remodeling: Involvement of Nuclear Factor-κB (NF-κB), Nuclear Factor Erythroid 2p45-Related Factor-2 (Nrf2), and Hypoxia-Inducible Factor (HIF). International Journal of Molecular Sciences. 2012; 13(7):7915-7937. https://doi.org/10.3390/ijms13077915
Chicago/Turabian StylePark, Seoung Ju, Kyung Sun Lee, Su Jeong Lee, So Ri Kim, Seung Yong Park, Myoung Shin Jeon, Heung Bum Lee, and Yong Chul Lee. 2012. "l-2-Oxothiazolidine-4-Carboxylic Acid or α-Lipoic Acid Attenuates Airway Remodeling: Involvement of Nuclear Factor-κB (NF-κB), Nuclear Factor Erythroid 2p45-Related Factor-2 (Nrf2), and Hypoxia-Inducible Factor (HIF)" International Journal of Molecular Sciences 13, no. 7: 7915-7937. https://doi.org/10.3390/ijms13077915