Enhancement of Palmarumycins C12 and C13 Production in Liquid Culture of Endophytic Fungus Berkleasmium sp. Dzf12 after Treatments with Metal Ions


Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of Metal Ions on Mycelia Growth and Palmarumycin Production
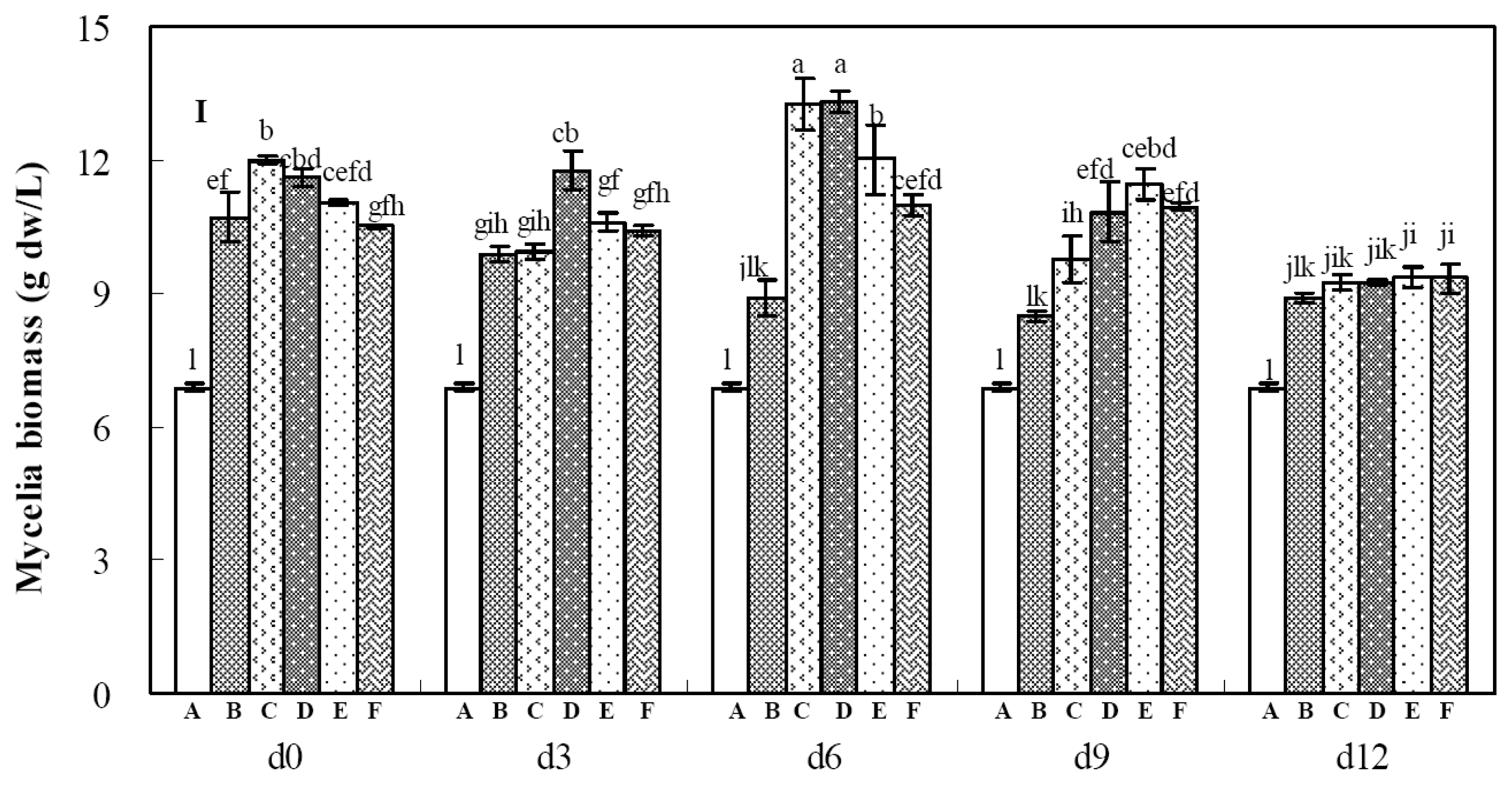
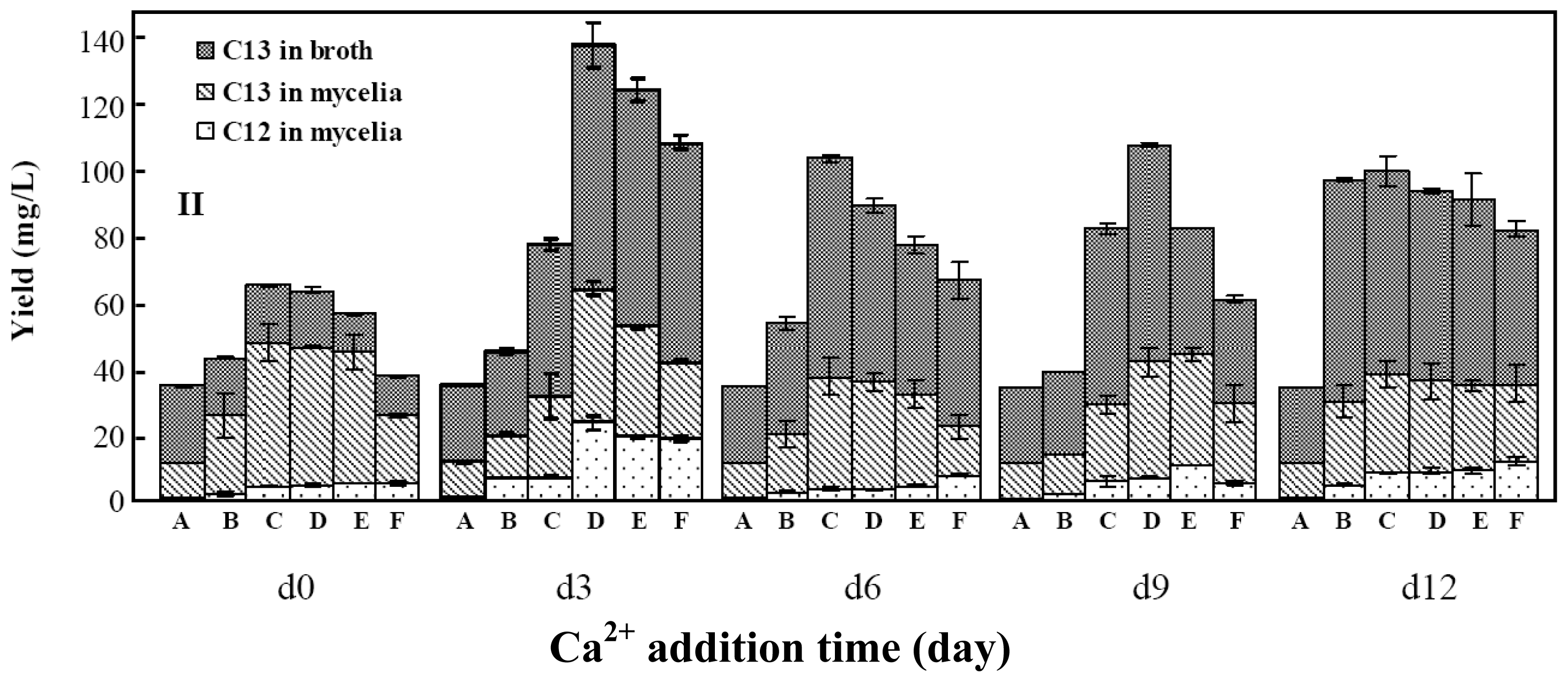
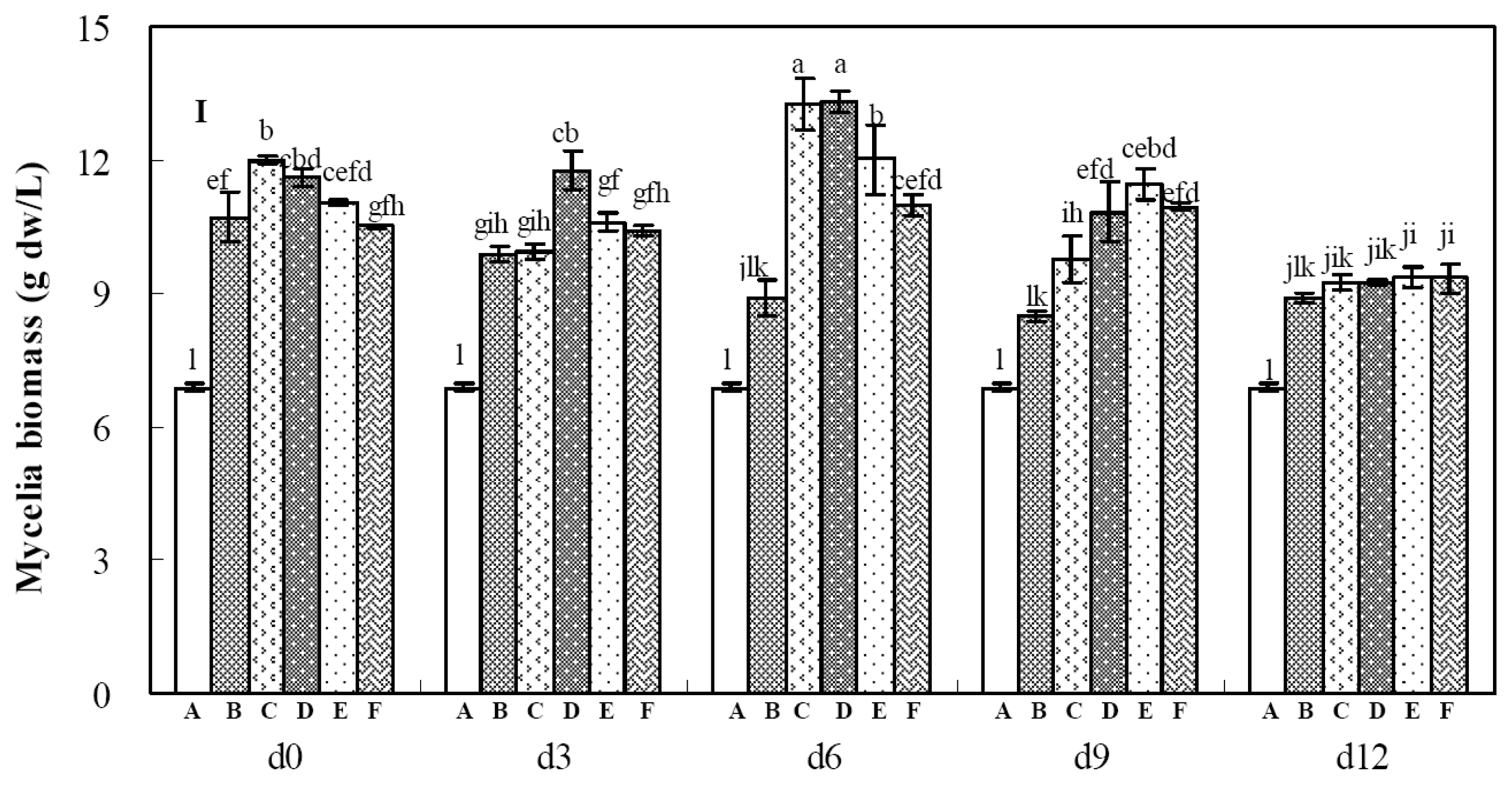
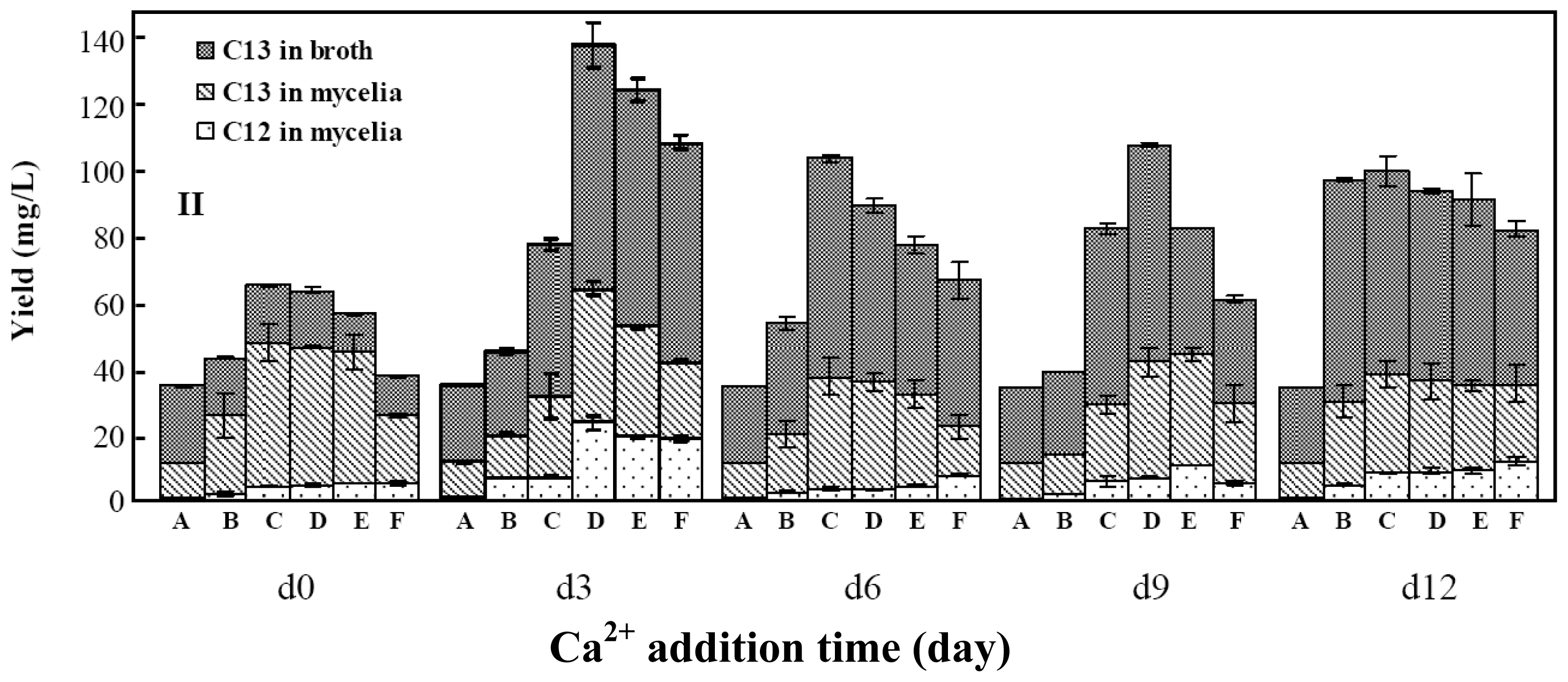
2.2. Effects of Calcium Ion Addition Time on Mycelia Growth and Palmarumycin Production
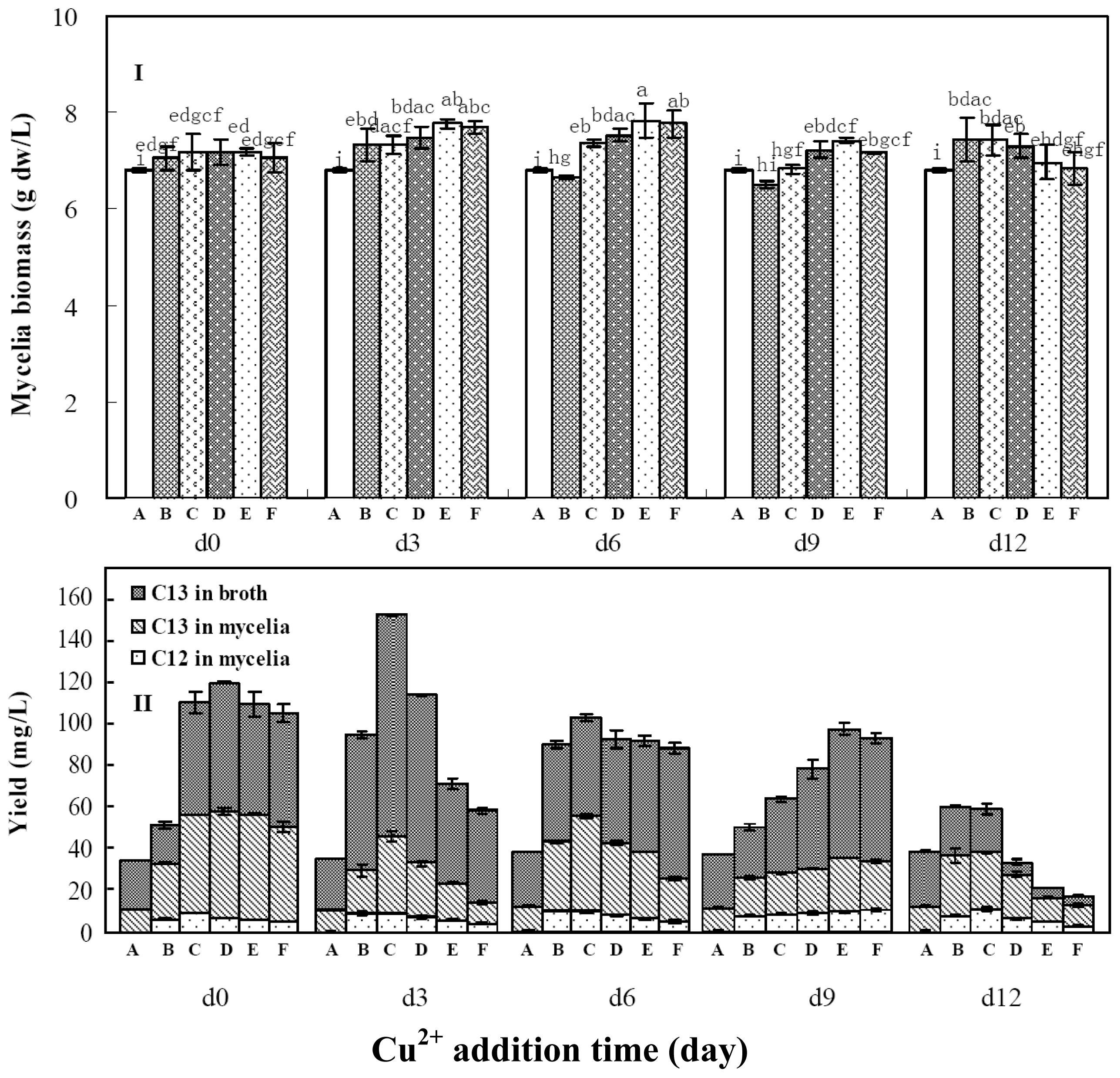
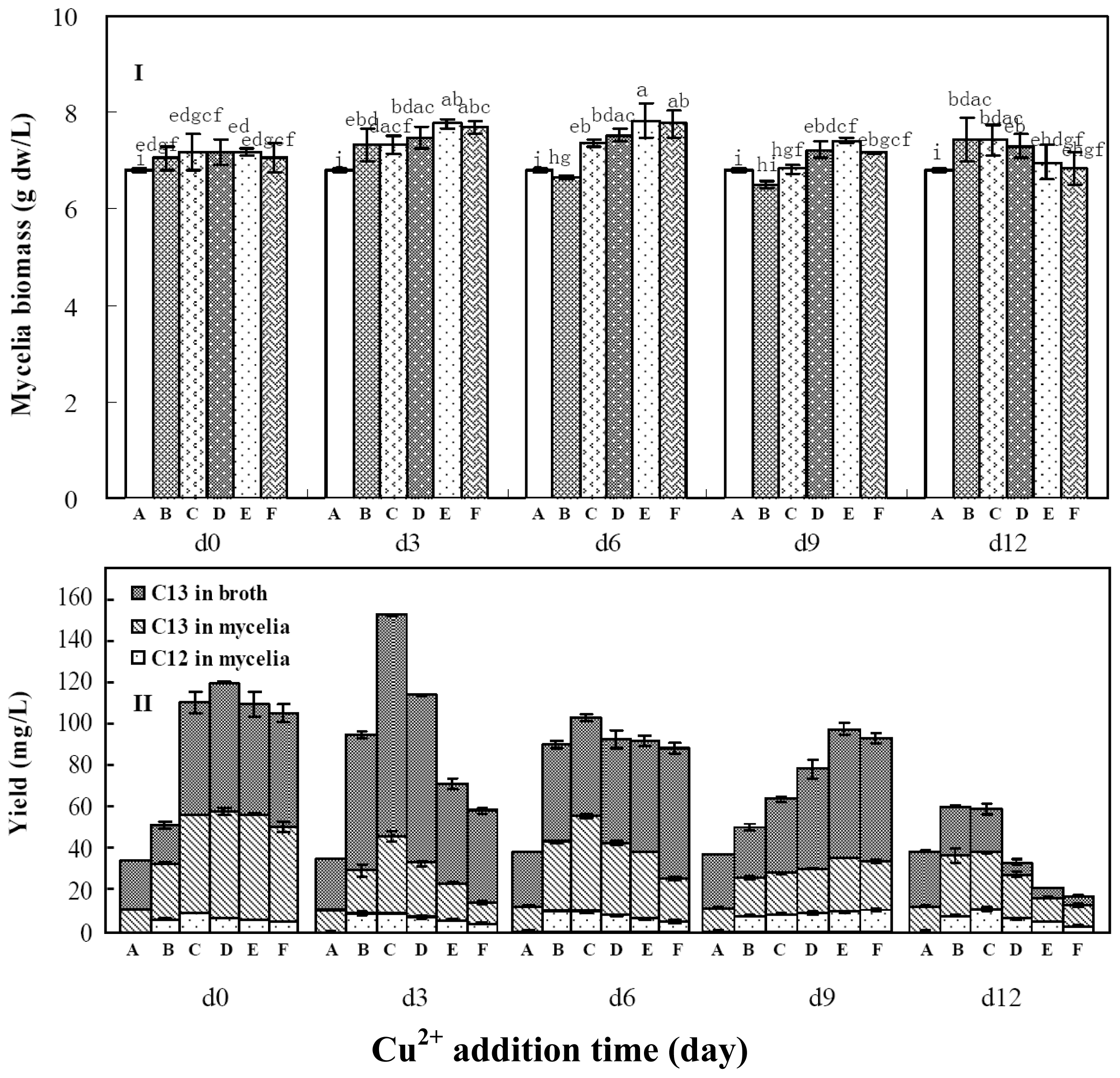
2.3. Effects of Copper Ion Addition Time on Mycelia Growth and Palmarumycin Production
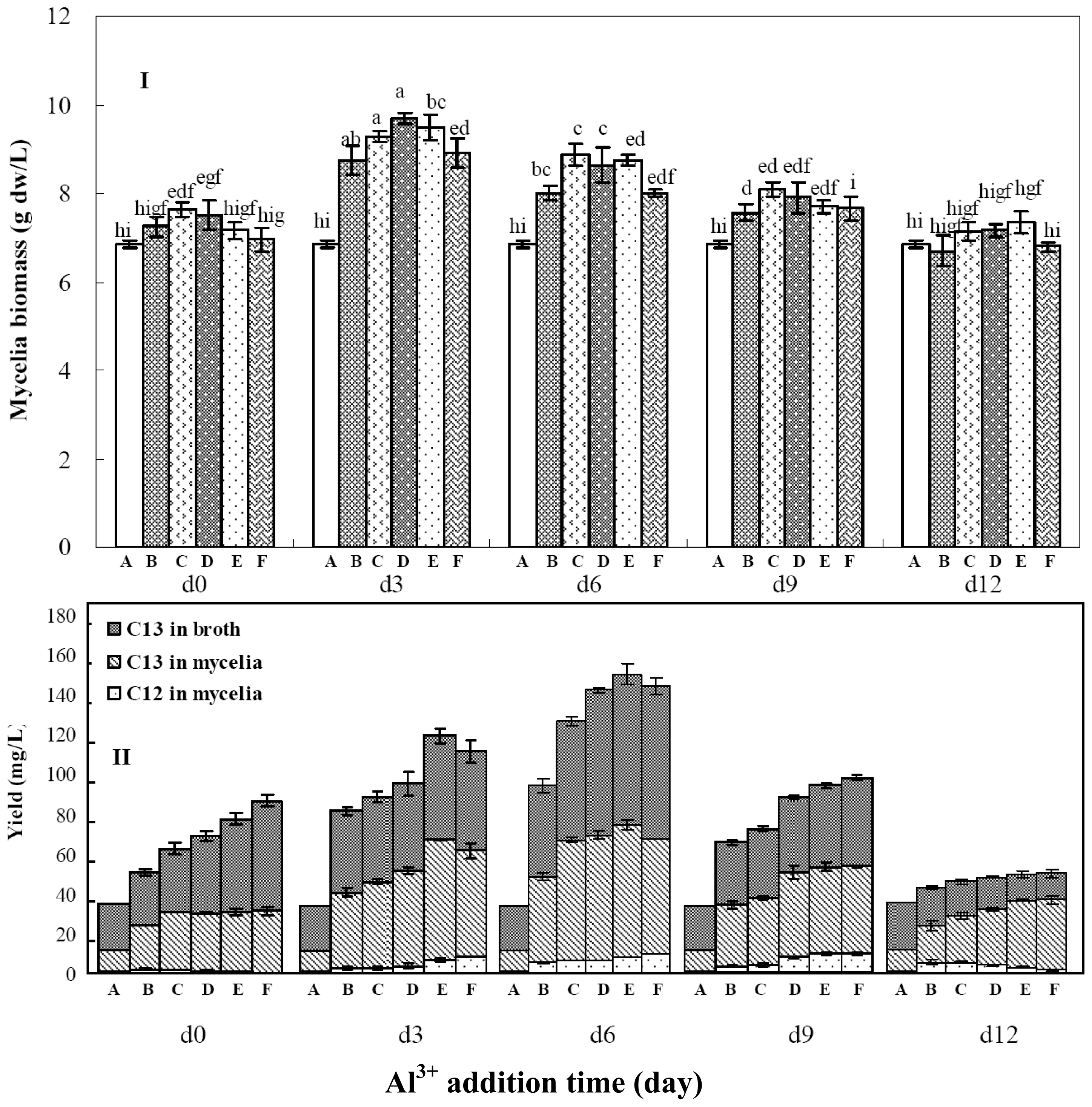
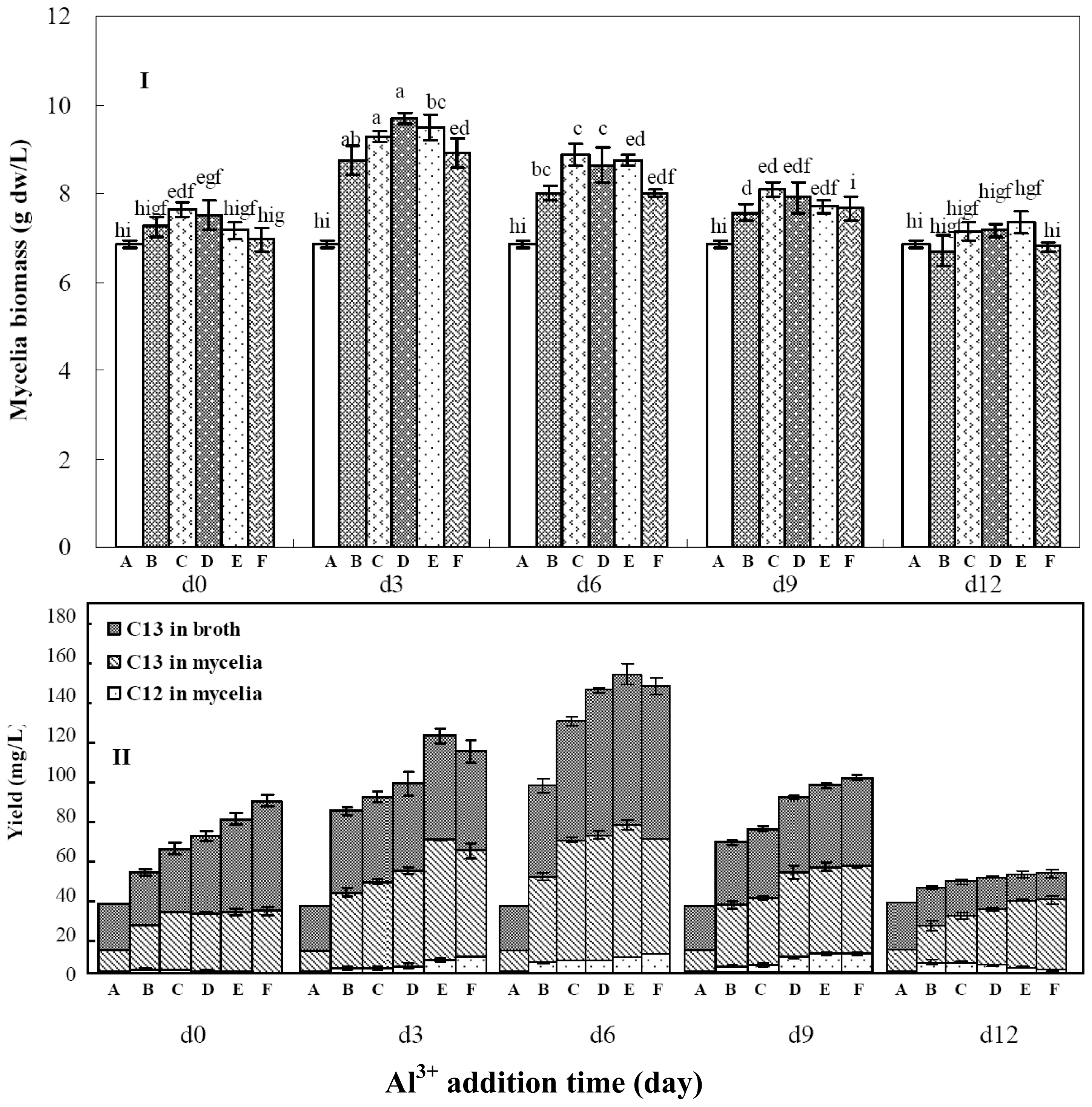
2.4. Effects of Aluminum Ion Addition Time on Mycelia Growth and Palmarymycin Production
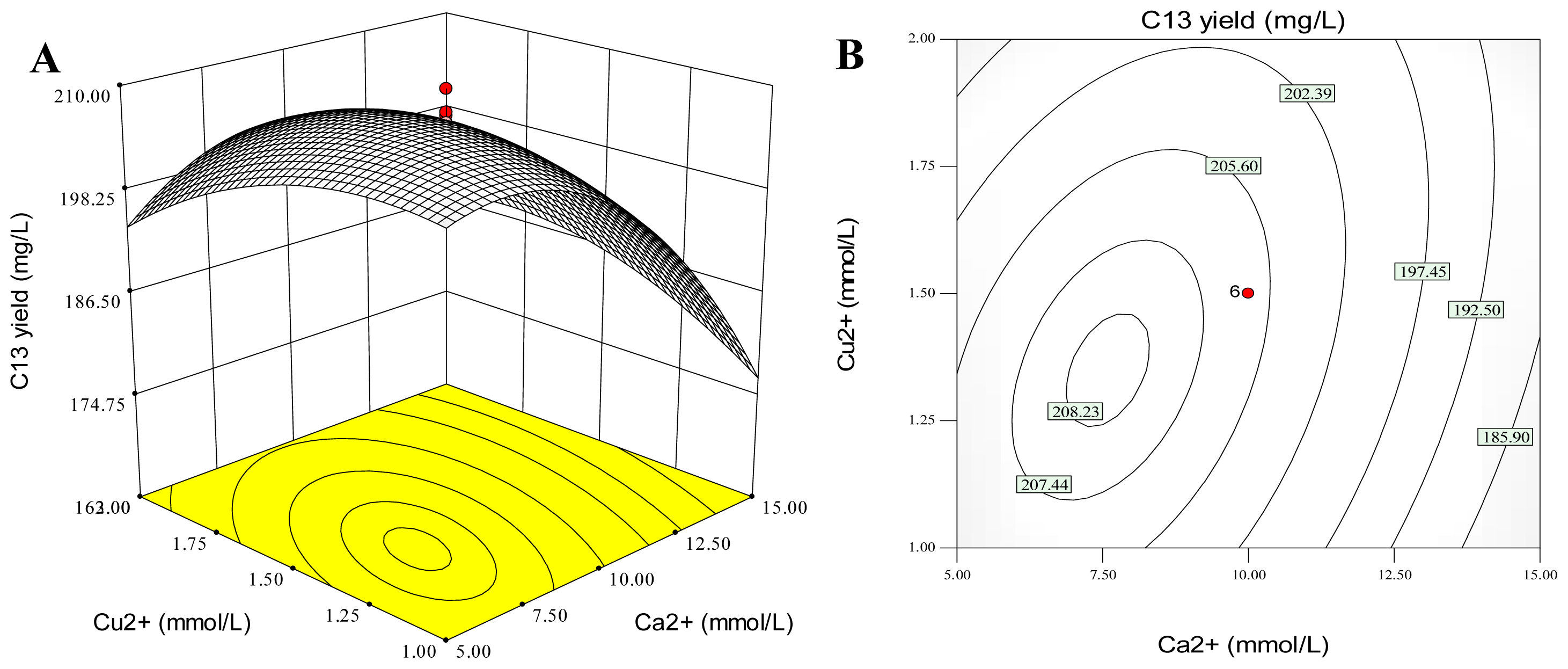
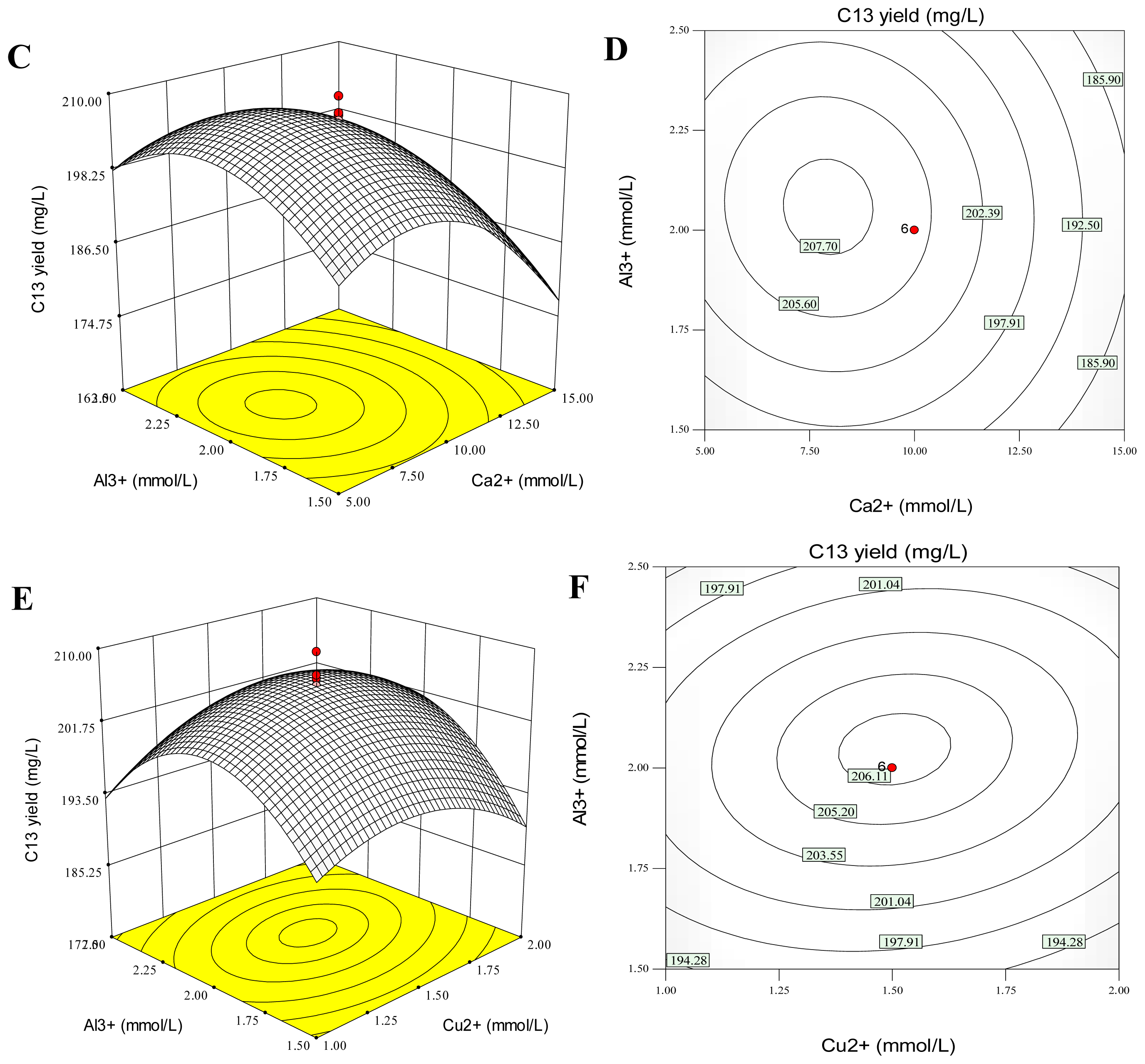
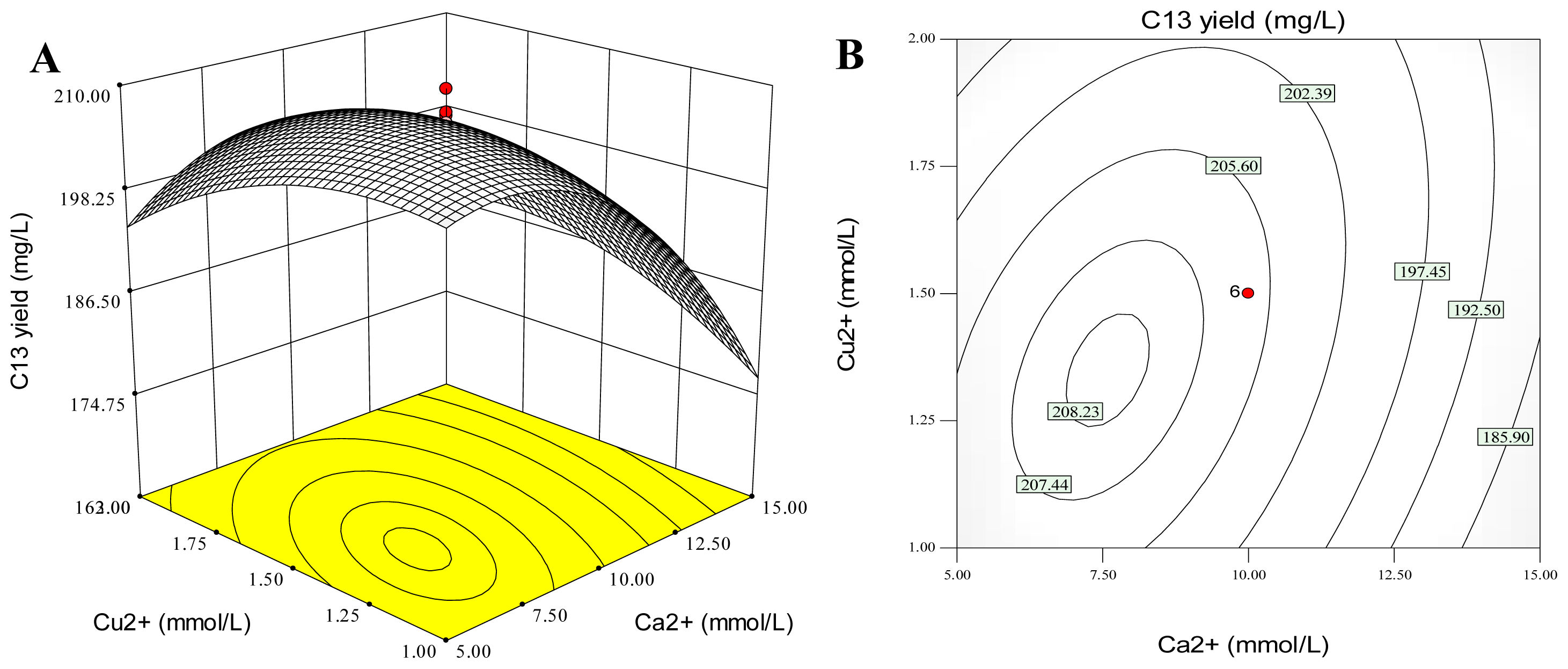
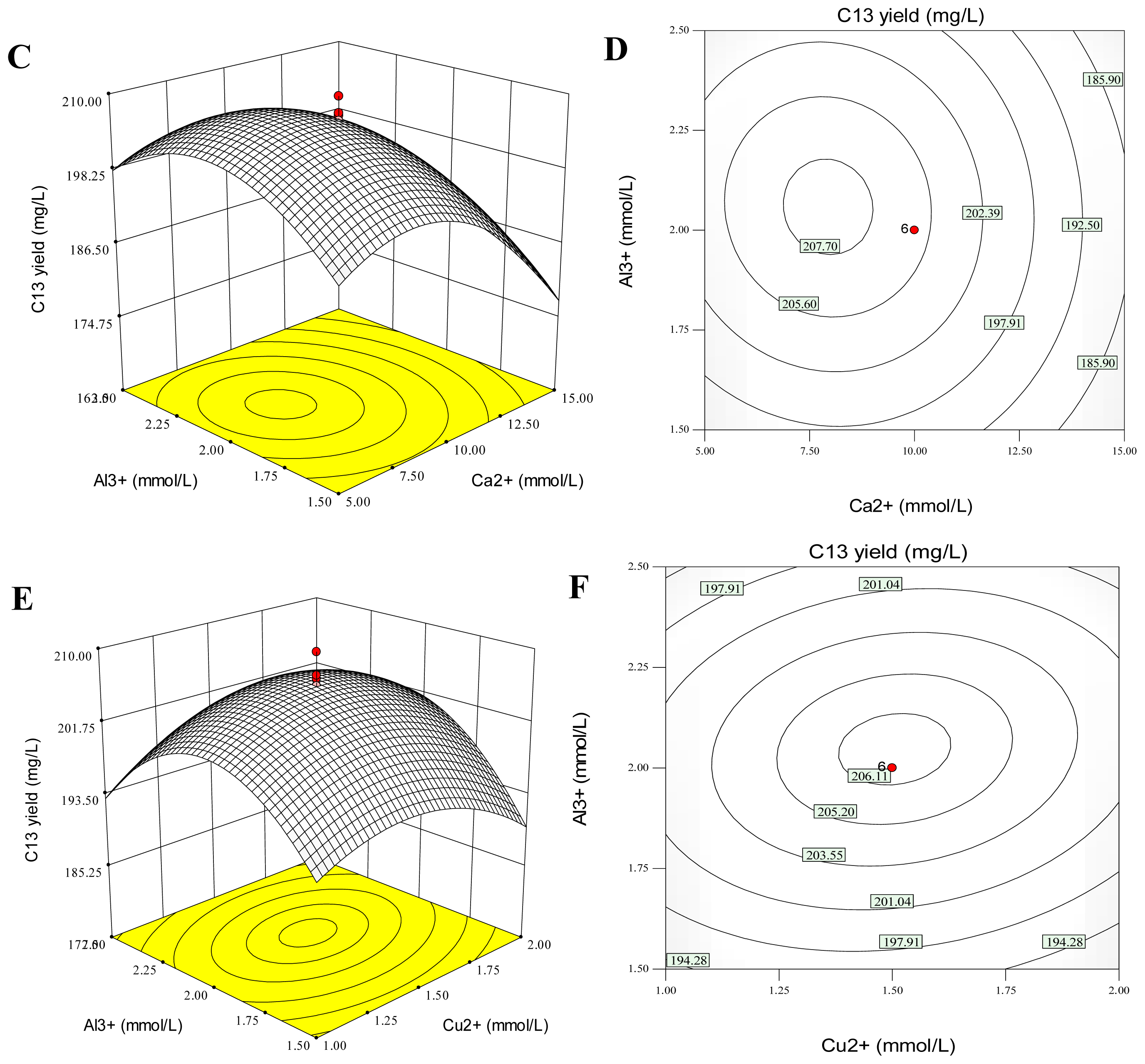
2.5. Combination Effects of Calcium, Copper and Aluminum Ions on Palmarumycin C13 Production
3. Experimental Section
3.1. Endophytic Fungus and Culture Conditions
3.2. Application of the Metal Ions
3.3. Determination of Mycelia Biomass
3.4. Extraction and Quantification of Palmarumycins C12 and C13
3.5. Experimental Design for the Combination Effects of the Metal Ions
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Divers 2010, 41, 1–16. [Google Scholar]
- Gunatilaka, A.A.L. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity, and implications of their occurrence. J. Nat. Prod 2006, 69, 509–526. [Google Scholar]
- Schulz, B.; Boyle, C.; Draeger, S.; Rommert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res 2002, 106, 996–1004. [Google Scholar]
- Kharwar, R.N.; Mishra, A.; Gond, S.K.; Stierle, A.; Stierle, D. Anticancer compounds derived from fungal endophytes: their importance and future challenges. Nat. Prod. Rep 2011, 28, 1208–1228. [Google Scholar]
- Wang, L.-W.; Zhang, Y.-L.; Lin, F.-C.; Hu, Y.-Z.; Zhang, C.-L. Natural products with antitumor activity from endophytic fungi. Mini-Rev. Med. Chem 2011, 11, 1056–1074. [Google Scholar]
- Gutierrez, R.M.P.; Gonzalez, A.M.N.; Ramirez, A.M. Compounds derived from endophytes: A review of phytochemistry and pharmacology. Curr. Med. Chem 2012, 19, 2992–3030. [Google Scholar]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini-Rev. Med. Chem 2011, 11, 159–168. [Google Scholar]
- Cai, X.; Shan, T.; Li, P.; Huang, Y.; Xu, L.; Zhou, L.; Wang, M.; Jiang, W. Spirobisnaphthalenes from the endophytic fungus Dzf12 of Dioscorea zingiberensis and their antimicrobial activities. Nat. Prod. Commun 2009, 4, 1469–1472. [Google Scholar]
- Zhang, R.; Li, P.; Zhao, J.; Yin, C.; Zhou, L. Endophytic fungi from Dioscorea zingiberensis and their effects on the growth and diosgenin production of the host plant cultures. Nat. Prod. Res. Dev 2010, 22, 11–15. [Google Scholar]
- Li, Y.; Shan, T.; Mou, Y.; Li, P.; Zhao, J.; Zhao, W.; Peng, Y.; Zhou, L.; Ding, C. Enhancement of palmarumycin C12 and C13 production in liquid culture of the endophytic fungus Berkleasmium sp. Dzf12 by oligosaccharides from its host plant Dioscorea zingiberensis. Molecules 2012, 17, 3761–3773. [Google Scholar]
- Krohn, K.; Michel, A.; Floerke, U.; Aust, H.-J.; Draeger, S.; Schulz, B. Palmarumycins C1–C16 from Coniothyrium sp.: Isolation, structure elucidation, and biological activity. Liebigs. Ann. Chem 1994, 11, 1099–1108. [Google Scholar]
- Schlingmann, G.; West, R.R.; Milne, L.; Pearce, C.J.; Carter, G.T. Diepoxins, novel fungal metabolites with antibiotic activity. Tetrahedron Lett 1993, 34, 7225–7228. [Google Scholar]
- Chu, M.; Truumees, I.; Patel, M.G.; Gullo, V.P.; Blood, C.; King, I.; Pai, J.K.; Puar, M.S. A novel class of anti-tumor metabolites from the fungus Nattrassia mangiferae. Tetrahedron Lett 1994, 35, 1343–1346. [Google Scholar]
- Zhou, L.; Zhao, J.; Shan, T.; Cai, X.; Peng, Y. Spirobisnaphthalenes from fungi and their biological activities. Mini-Rev. Med. Chem 2010, 10, 977–989. [Google Scholar]
- Cai, Y.-S.; Guo, Y.-W.; Krohn, K. Structure, bioactivities, biosynthetic relationships and chemical synthesis of the spirodioxynaphthalenes. Nat. Prod. Rep 2010, 27, 1840–1870. [Google Scholar]
- Zhou, L.G.; Wu, J.Y. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Nat. Prod. Rep 2006, 23, 789–810. [Google Scholar]
- Zhou, L.; Cao, X.; Zhang, R.; Peng, Y.; Zhao, S.; Wu, J. Stimulation of saponin production in Panax ginseng hairy roots by two oligosaccharides from Paris polyphylla var. yunnanensis. Biotechnol. Lett 2007, 29, 631–634. [Google Scholar]
- Schneider, P.; Misiek, M.; Hoffmeister, D. In vivo and in vitro production options for fungal secondary metabolites. Mol. Pharmaceut 2008, 5, 234–242. [Google Scholar]
- Zhang, R.; Li, P.; Xu, L.; Chen, Y.; Sui, P.; Zhou, L.; Li, J. Enhancement of diosgenin production in Dioscorea zingiberensis cell culture by oligosaccharide elicitor from its endophytic fungus Fusarium oxysporum Dzf17. Nat. Prod. Commun 2009, 4, 1459–1462. [Google Scholar]
- Xu, L.; Liu, Y.; Zhou, L.; Wu, J. Enhanced beauvericin production with in situ adsorption in mycelial liquid culture of Fusarium redolens Dzf2. Process Biochem 2009, 44, 1063–1067. [Google Scholar]
- Xu, L.; Liu, Y.; Zhou, L.; Wu, J. Optimization of a liquid medium for beauvericin production in Fusarium redolens Dzf2 mycelial culture. Biotechnol. Bioproc. Eng 2010, 15, 460–466. [Google Scholar]
- Zhao, J.; Zhou, L.; Wu, J. Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures. Appl. Microbiol. Biotechnol 2010, 87, 137–144. [Google Scholar]
- Li, P.; Mou, Y.; Shan, T.; Xu, J.; Li, Y.; Lu, S.; Zhou, L. Effects of polysaccharide elicitors from endophytic Fusarium oxysporum Dzf17 on growth and diosgenin production in cell suspension culture of Dioscorea zingiberensis. Molecules 2011, 16, 9003–9016. [Google Scholar]
- Li, P.; Mao, Z.; Lou, J.; Li, Y.; Mou, Y.; Lu, S.; Peng, Y.; Zhou, L. Enhancement of diosgenin production in Dioscorea zingiberensis cell cultures by oligosaccharides from its endophytic fungus Fusarium oxysporum Dzf17. Molecules 2011, 16, 10631–10644. [Google Scholar]
- Metwally, M.; Abou-Zeid, A. Effects of toxic heavy metals on growth and metabolic activity of some fungi. Egypt. J. Microbiol 1996, 31, 115–127. [Google Scholar]
- Flores-Cotera, L.B.; Sanchez, S. Copper but not iron limitation increases astaxanthin production by Phaffia rhodozyma in a chemically defined medium. Biotechnol. Lett 2001, 23, 793–797. [Google Scholar]
- Zhou, Y.; Du, J.; Tsao, G.T. Mycelial pellet formation by Rhizopus oryzae ATCC 20344. Appl. Biochem. Biotechnol 2000, 84, 779–789. [Google Scholar]
- Cui, Y.-H.; Zhang, K.-C. Effect of metal ions on the growth and metabolites production of Ganodenma lucidum in submerged culture. Afr. J. Biotechnol 2011, 10, 11983–11989. [Google Scholar]
- Zhao, J.; Zheng, B.; Li, Y.; Shan, T.; Mou, Y.; Lu, S.; Li, P.; Zhou, L. Enhancement of diepoxin ζ production by yeast extract and its fractions in liquid culture of Berkleasmium-like endophytic fungus Dzf12 from Dioscorea zingiberensis. Molecules 2011, 16, 847–856. [Google Scholar]
- Zhao, J.; Li, Y.; Shan, T.; Mou, Y.; Zhou, L. Enhancement of diepoxin ζ production with in situ resin adsorption in mycelial liquid culture of the endophytic fungus Berkleasmium sp. Dzf12 from Dioscorea zingiberensis. World J. Microbiol. Biotechnol 2011, 27, 2753–2758. [Google Scholar]
- Li, Y.; Li, P.; Mou, Y.; Zhao, J.; Shan, T.; Ding, C.; Zhou, L. Enhancement of diepoxin ζ production in liquid culture of endophytic fungus Berkleasmium sp. Dzf12 by polysaccharides from its host plant Dioscorea zingiberensis. World J. Microbiol. Biotechnol 2012, 28, 1407–1413. [Google Scholar]
- Bode, H.B.; Wegner, B.; Zeeck, A. Biosynthesis of cladospirone bisepoxide, a member of the spirobisnaphthalene family. J. Antibiot 2000, 53, 153–157. [Google Scholar]
- Bode, H.B.; Zeeck, A. UV mutagenesis and enzyme inhibitors as tools to elucidate the late biosynthesis of the spirobisnaphthalenes. Phytochemistry 2000, 55, 311–316. [Google Scholar]
- Cuero, R.; Ouellet, T. Metal ions modulate gene expression and accumulation of the mycotoxins aflatoxin and zearalenone. J. Appl. Microbiol 2005, 98, 598–605. [Google Scholar]
- Wei, Q.; Liu, F.; Zhang, S.; You, T.; Chen, G.; Wen, L. Study on the relation between several metal ions and the growth metabolism of a symbiotic fungus of Sesarma bidens. Guangdong Weiliang Yuansu Kexue 2012, 19, 16–21. [Google Scholar]
- Chardonnet, C.O.; Sams, C.E.; Conway, W.S. Calcium effect on the mycelial cell walls of Botrytis cinerea. Phytochemistry 1999, 52, 967–973. [Google Scholar]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signaling: Dynamics, homeostasis and remodeling. Nat. Rev. Mol. Cell Biol 2003, 4, 517–529. [Google Scholar]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar]
- Cavinder, B.; Hamam, A.; Lew, R.R.; Trail, F. Mid1, a mechanosensitive calcium ion channel, affects growth, development, and ascospore discharge in the filamentous fungus Gibberella zeae. Eukaryotic Cell 2011, 10, 832–841. [Google Scholar]
- Tychanowicz, G.K.; Souza, D.F.; Souza, C.G.M.; Kadowaki, M.K.; Peralta, R.M. Copper improves the production of laccase by the white-rot fungus Pleurotus pulmonarius in solid state fermentation. Braz. Arch. Biol. Technol 2006, 49, 699–704. [Google Scholar]
- Tsunoda, K. Toxicity of aluminum and its speciation analysis. Biomed. Res. Trace Elem 2005, 16, 276–280. [Google Scholar]
- Berthon, G. Aluminum speciation in relation to aluminum bioavailability, metabolism and toxicity. Coord. Chem. Rev 2002, 228, 319–341. [Google Scholar]
- Haque, R.M.; Kuo, Y.-H.; Lambein, F.; Hussain, M. Effect of environmental factors on the biosynthesis of the neuro-excitatory amino acid β-ODAP (β-N-oxalyl-α,β-diaminopropionic acid) in callus tissue of Lathyrus sativus. Food Chem. Toxicol 2011, 49, 583–588. [Google Scholar]
- Bejan, C.; Visoiu, E. Resveratrol biosynthesis on in vitro conditions in grapevine (cv. Feteasca Neagra and Cabernet Sauvignon). Rom. Biotechnol. Lett 2011, 16, S97–S101. [Google Scholar]
- Karimi, F.; Khataee, E. Aluminum elicits tropane alkaloid production and antioxidant system activity in micropropagated Datura innoxia plantlets. Acta Physiol. Plant 2012, 34, 1035–1041. [Google Scholar]
- Pujari, V.; Chandra, T.S. Statistical optimization of medium components for enhanced riboflavin production by a UV-mutant of Eremothecium ashbyii. Process Biochem 2000, 36, 31–37. [Google Scholar]
- Gao, H.; Gu, W.-Y. Optimization of polysaccharide and ergosterol production from Agaricus brasiliensis by fermentation process. Biochem. Eng. J 2007, 33, 202–210. [Google Scholar]
- Liu, J.-Z.; Weng, L.-P.; Zhan, Q.-L.; Xu, H.; Ji, L.N. Optimization of glucose oxidase production by Aspergillus niger in a benchtop bioreactor using response surface methodology. World J. Microbiol. Biotechnol 2003, 19, 317–323. [Google Scholar]
- Box, G.E.P.; Wilson, K.B. On the experimental attainment of optimum conditions. J. Roy. Stat. Soc. B 1951, 13, 1–45. [Google Scholar]
- Kiran, B.; Kaushik, A.; Kaushik, C.P. Response surface methodological approach for optimizing removal of Cr (VI) from aqueous solution using immobilized cyanobacterium. Chem. Eng. J 2007, 126, 147–153. [Google Scholar]
- Muralidhar, R.V.; Chirumamila, R.R.; Marchant, R.; Nigam, P. A response surface approach for the comparison of lipase production by Candida cylindracea using two different carbon sources. Biochem. Eng. J 2001, 9, 17–23. [Google Scholar]
- Wu, Y.; Cui, S.W.; Tang, J.; Gu, X. Optimizaiton of extraction process of crude polysaccharides from boat-fruited sterculia seeds by response surface methodology. Food Chem 2007, 105, 1599–1605. [Google Scholar]
- Soto-Cruz, O.; Saucedo-Castaneda, G.; Pablos-Hach, J.L.; Gutierrez-Rojas, M.; Favela-Torres, E. Effect of substrate composition on the mycelial growth of Pleurotus ostreatus. An analysis by mixture and response surface methodologies. Process Biochem 1999, 35, 127–133. [Google Scholar]
- Triveni, R.; Shamala, T.R.; Rastogi, N.K. Optimized production and utilization of exopolysaccharide from Agrobacterium radiobacter. Process Biochem 2001, 36, 787–795. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal ion | Conc. (mmol/L) | Mycelia biomass (g dw/L) | C12 yield (mg/L) | C13 yield (mg/L) | C12 plus C13 yield (mg/L) |
|---|---|---|---|---|---|
| CK | 0.00 | 6.79 ± 0.08 cde | 1.29 ± 0.03 fgh | 31.03 ± 0.06 h | 32.32 ± 0.13 g |
| Na+ | 5.00 | 5.81 ± 0.18 fg | 0.66 ± 0.01 ghi | 16.10 ± 0.21 ij | 16.76 ± 0.32 ij |
| 10.00 | 5.83 ± 0.13 fg | 3.00 ± 0.17 e | 19.25 ± 0.34 i | 22.25 ± 1.51 hi | |
| 20.00 | 5.96 ± 0.25 efg | 2.06 ± 0.19 e | 19.39 ± 0.15 i | 21.45 ± 0.38 hi | |
| Ca2+ | 5.00 | 7.26 ± 0.17 bcd | 4.98 ± 0.21 d | 91.57 ± 1.76 d | 96.55 ± 2.66 c |
| 10.00 | 7.32 ± 0.44 bcd | 14.76 ± 0.31 a | 114.48 ± 6.76 a | 129.24 ± 7.12 a | |
| 20.00 | 7.58 ± 0.10 bc | 13.79 ± 0.90 a | 83.48 ± 2.37 e | 97.27 ± 3.63 c | |
| Ag+ | 0.05 | 3.21 ± 0.29 k | 0.00 ± 0.00 i | 0.67 ± 0.13 | l 0.67 ± 0.13 k |
| 0.10 | 3.17 ± 0.26 k | 0.00 ± 0.00 i | 0.44 ± 0.16 l | 0.44 ± 0.16 k | |
| 0.50 | 2.99 ± 0.31 k | 0.00 ± 0.00 i | 0.32 ± 0.09 l | 0.32 ± 0.09 k | |
| Co2+ | 0.05 | 4.35 ± 0.50 ij | 0.00 ± 0.00 i | 0.30 ± 0.02 l | 0.30 ± 0.02 k |
| 0.10 | 4.29 ± 0.18 ij | 0.00 ± 0.00 i | 0.39 ± 0.09 l | 0.39 ± 0.09 k | |
| 0.50 | 3.52 ± 0.76 jk | 0.00 ± 0.00 i | 0.21 ± 0.10 l | 0.21 ± 0.10 k | |
| Cu2+ | 0.50 | 7.51 ± 0.33 b | 1.42 ± 0.18 fg | 78.97 ± 0.26 e | 80.39 ± 4.13 d |
| 1.50 | 7.67 ± 0.22 b | 6.65 ± 0.06 c | 105.61 ± 1.70 b | 112.26 ± 3.07 b | |
| 3.00 | 7.82 ± 0.13 b | 4.62 ± 0.21d | 63.84 ± 0.86 f | 68.46 ± 1.06 e | |
| Al3+ | 0.50 | 7.08 ± 0.33 bcd | 2.20 ± 0.86 ef | 51.49 ± 2.16 g | 53.69 ± 4.77 f |
| 1.50 | 8.09 ± 0.29 ab | 8.38 ± 0.85 b | 97.83 ± 3.67 cd | 106.21 ± 5.50 bc | |
| 3.00 | 8.82 ± 0.33 a | 9.00 ± 0.62 d | 103.82 ± 5.80 bc | 112.82 ± 9.41 b | |
| Zn2+ | 2.00 | 7.09 ± 0.27 bcd | 0.32 ± 0.15 ghi | 15.85 ± 1.97 ij | 16.17 ± 1.57 ij |
| 4.00 | 7.66 ± 0.18 bc | 0.63 ± 0.13 ghi | 31.42 ± 2.54 h | 32.05 ± 1.63 g | |
| 8.00 | 6.49 ± 0.39 def | 0.59 ± 0.14 ghi | 29.67 ± 1.58 h | 30.26 ± 2.01 gh | |
| Mn4+ | 2.00 | 4.79 ± 0.28 hi | 0.14 ± 0.01 hi | 7.23 ± 0.42 kl | 7.37 ± 0.77 jk |
| 4.00 | 5.43 ± 0.51 gh | 0.15 ± 0.02 hi | 7.43 ± 0.81 kl | 7.58 ± 0.52 jk | |
| 8.00 | 5.82 ± 0.49 fg | 0.20 ± 0.00 hi | 10.10 ± 0.19 kj | 10.30 ± 1.01 jk | |
| Aluminum ion conc. (mmol/L) | Mycelia biomass (g dw/L) | C12 yield (mg/L) | C13 yield (mg/L) | C12 plus C13 yield (mg/L) | Ratio of C12 yield to C13 yield |
|---|---|---|---|---|---|
| 0 | 7.35 ± 0.17 d | 1.83 ± 0.48 c | 38.14 ± 3.21 d | 39.97 ± 3.07 d | 0.05 |
| 2 | 11.16 ± 0.24 abc | 9.98 ± 1.47 c | 117.26 ± 5.70 b | 127.24 ± 4.25 c | 0.09 |
| 4 | 11.85 ± 0.41 ab | 14.94 ± 1.25 c | 166.27 ± 3.42 a | 181.21 ± 2.87 b | 0.09 |
| 6 | 11.76 ± 1.21 ab | 106.59 ± 5.31 b | 162.93 ± 4.08 a | 269.52 ± 5.19 a | 0.65 |
| 8 | 13.17 ± 0.39 a | 224.33 ±10.08 a | 59.23 ± 2.72 c | 283.56 ± 8.08 a | 3.79 |
| 10 | 10.51 ± 1.01 bc | 118.85 ± 3.93 b | 4.19 ± 0.93 e | 123.04 ± 3.91 c | 28.36 |
| 12 | 9.49 ± 0.98 cd | 115.55 ± 2.97 b | 3.58 ± 0.13 e | 119.13 ± 2.88 c | 32.28 |
| Variable (mmol/L) | Symbol | Coded level | |||||
|---|---|---|---|---|---|---|---|
| Uncoded | Coded | −1.682 | −1 | 0 | 1 | +1.682 | |
| Ca2+ | X1 | x1 | 1.59 | 5 | 10 | 15 | 18.41 |
| Cu2+ | X2 | x2 | 0.65 | 1.0 | 1.5 | 2.0 | 2.34 |
| Al3+ | X3 | x3 | 1.16 | 1.5 | 2.0 | 2.5 | 2.84 |
| Run | x1 | x2 | x3 | Palmarumycin C13 yield (mg/L) | ||
|---|---|---|---|---|---|---|
| Experimental Ye | Predicted Yp | Ye − Yp | ||||
| 1 | 1.68 | 0 | 0 | 163.22 | 161.47 | 1.75 |
| 2 | 0 | 0 | −1.68 | 177.98 | 179.53 | −1.55 |
| 3 | 0 | 0 | 0 | 202.40 | 206.31 | −3.91 |
| 4 | −1 | 1 | 1 | 189.29 | 190.14 | −0.85 |
| 5 | 0 | 0 | 0 | 206.74 | 206.31 | 0.43 |
| 6 | 1 | −1 | 1 | 166.33 | 167.58 | −1.25 |
| 7 | −1 | 1 | −1 | 182.75 | 181.70 | 1.05 |
| 8 | 0 | 0 | 0 | 209.64 | 206.31 | 3.33 |
| 9 | 1 | 1 | 1 | 179.77 | 181.50 | −1.73 |
| 10 | 0 | 0 | 0 | 205.98 | 206.31 | −0.33 |
| 11 | 0 | 0 | 0 | 207.10 | 206.31 | 0.79 |
| 12 | 0 | 1.68 | 0 | 194.98 | 193.70 | 1.28 |
| 13 | 0 | 0 | 1.68 | 186.67 | 184.83 | 1.84 |
| 14 | 1 | −1 | −1 | 170.37 | 169.72 | 0.65 |
| 15 | −1.68 | 0 | 0 | 189.26 | 190.72 | −1.46 |
| 16 | 0 | −1.68 | 0 | 192.91 | 193.90 | −0.99 |
| 17 | −1 | −1 | −1 | 197.38 | 195.86 | 1.52 |
| 18 | −1 | −1 | 1 | 198.36 | 197.78 | 0.58 |
| 19 | 1 | 1 | −1 | 176.34 | 177.12 | −0.78 |
| 20 | 0 | 0 | 0 | 205.94 | 206.31 | −0.37 |
| Source | Sum of squares | d.f. | Mean square | F value | Probability p > | F |
|---|---|---|---|---|---|---|
| Model | 3890.94 | 9 | 432.33 | 84.65 | <0.0001 | |
| Residual | 51.08 | 10 | 5.12 | |||
| Lack of fit | 23.61 | 5 | 4.72 | 0.86 | 0.56 | |
| Pure error | 27.46 | 5 | 5.49 | |||
| Corrected total | 3942.02 | 19 |
| Model Term | Coefficient estimate | Standard error | Sum of Squares | d.f. | Mean square | F value | Probability p > F |
|---|---|---|---|---|---|---|---|
| Intercept | 206.31 | 0.92 | |||||
| x1 | −8.70 | 0.61 | 1032.61 | 1 | 1032.61 | 202.17 | <0.0001 |
| x2 | −0.060 | 0.61 | 0.049 | 1 | 0.049 | 0.0095 | 0.9243 |
| x3 | 1.58 | 0.61 | 33.92 | 1 | 33.92 | 6.64 | 0.0275 |
| x1x2 | 5.39 | 0.80 | 232.41 | 1 | 232.41 | 45.50 | <0.0001 |
| x1x3 | −1.017 | 0.80 | 8.27 | 1 | 8.27 | 1.62 | 0.2320 |
| x2x3 | 1.63 | 0.80 | 21.23 | 1 | 21.23 | 4.16 | 0.0688 |
| x12 | −10.68 | 0.60 | 1644.34 | 1 | 1644.34 | 321.94 | <0.0001 |
| x22 | −4.42 | 0.60 | 281.79 | 1 | 281.79 | 55.17 | <0.0001 |
| x32 | −8.53 | 0.60 | 1048.77 | 1 | 1048.77 | 205.34 | <0.0001 |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mou, Y.; Luo, H.; Mao, Z.; Shan, T.; Sun, W.; Zhou, K.; Zhou, L. Enhancement of Palmarumycins C12 and C13 Production in Liquid Culture of Endophytic Fungus Berkleasmium sp. Dzf12 after Treatments with Metal Ions. Int. J. Mol. Sci. 2013, 14, 979-998. https://doi.org/10.3390/ijms14010979
Mou Y, Luo H, Mao Z, Shan T, Sun W, Zhou K, Zhou L. Enhancement of Palmarumycins C12 and C13 Production in Liquid Culture of Endophytic Fungus Berkleasmium sp. Dzf12 after Treatments with Metal Ions. International Journal of Molecular Sciences. 2013; 14(1):979-998. https://doi.org/10.3390/ijms14010979
Chicago/Turabian StyleMou, Yan, Haiyu Luo, Ziling Mao, Tijiang Shan, Weibo Sun, Kaiyi Zhou, and Ligang Zhou. 2013. "Enhancement of Palmarumycins C12 and C13 Production in Liquid Culture of Endophytic Fungus Berkleasmium sp. Dzf12 after Treatments with Metal Ions" International Journal of Molecular Sciences 14, no. 1: 979-998. https://doi.org/10.3390/ijms14010979