Physiological and Proteomic Responses of Diploid and Tetraploid Black Locust (Robinia pseudoacacia L.) Subjected to Salt Stress

Abstract
:1. Introduction
2. Results and Discussion
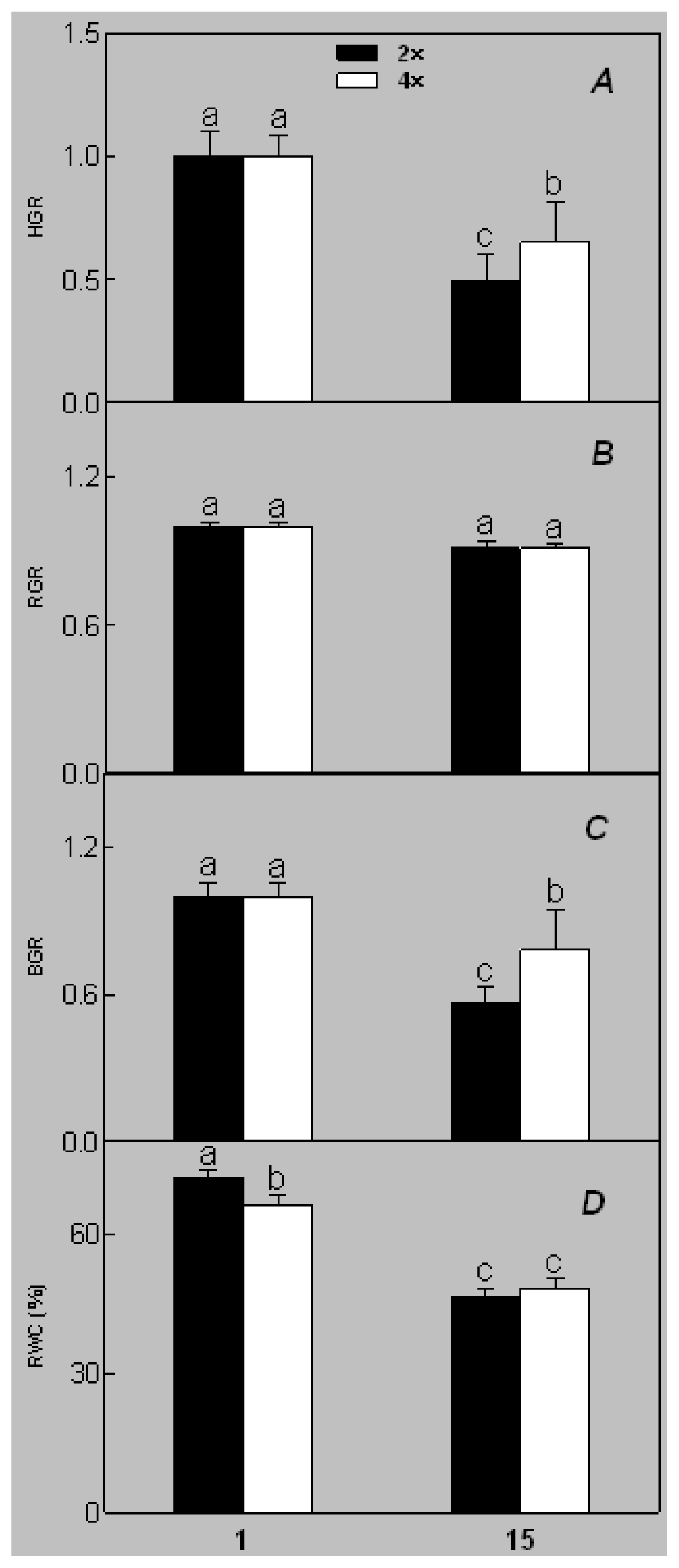
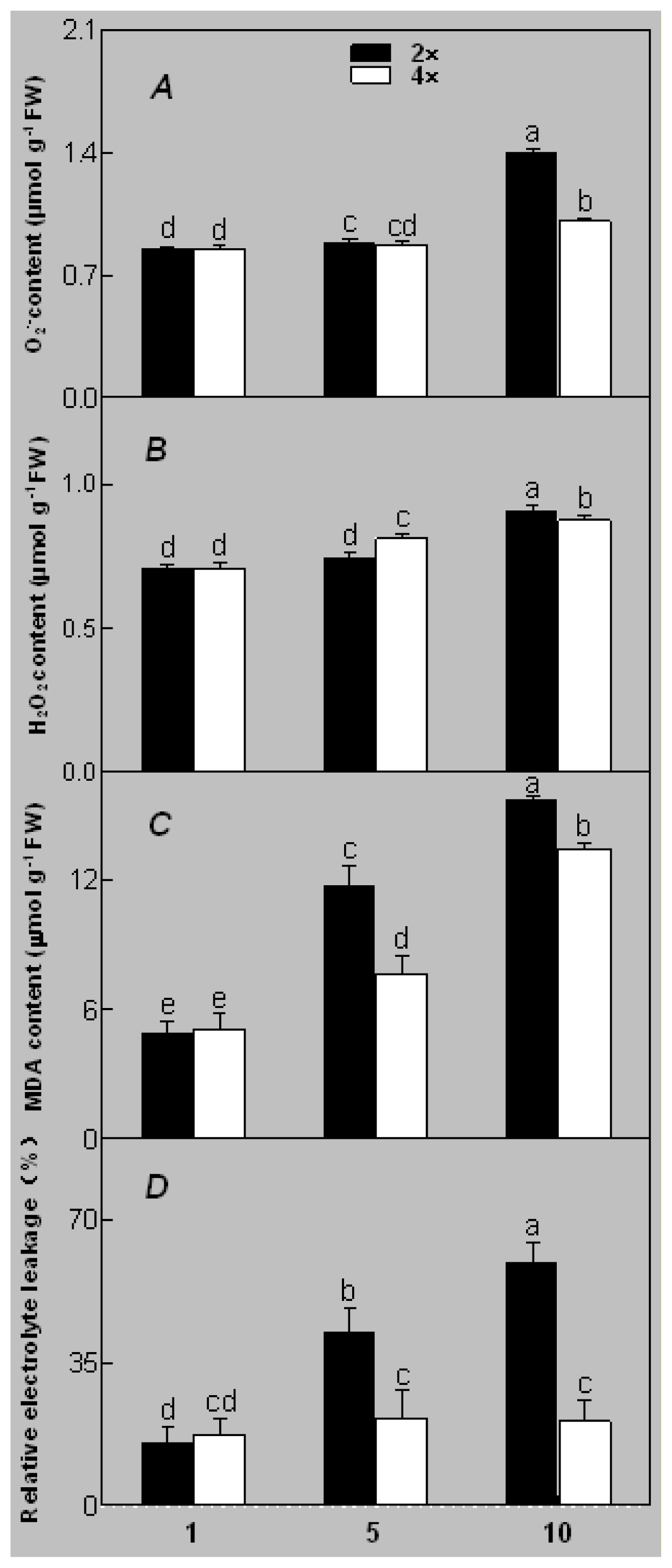
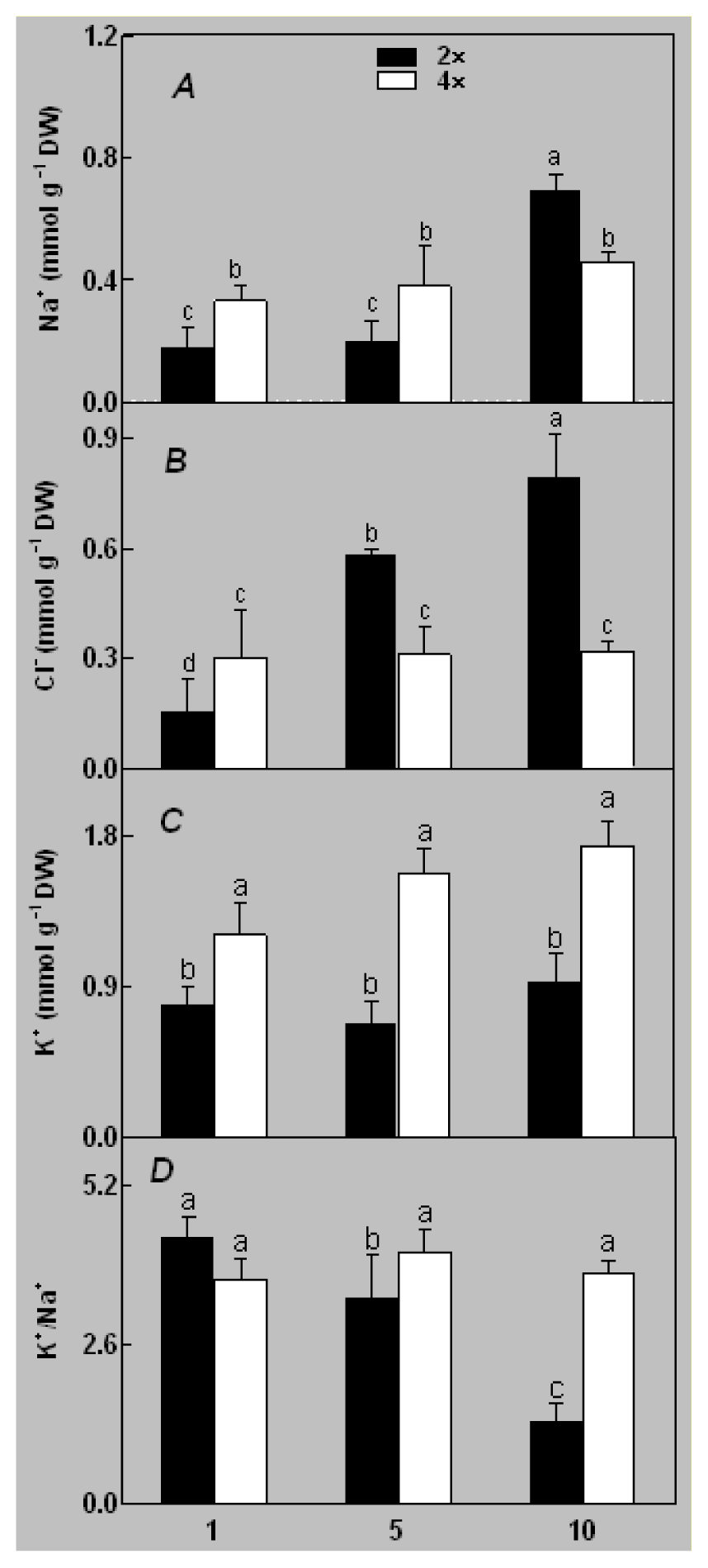
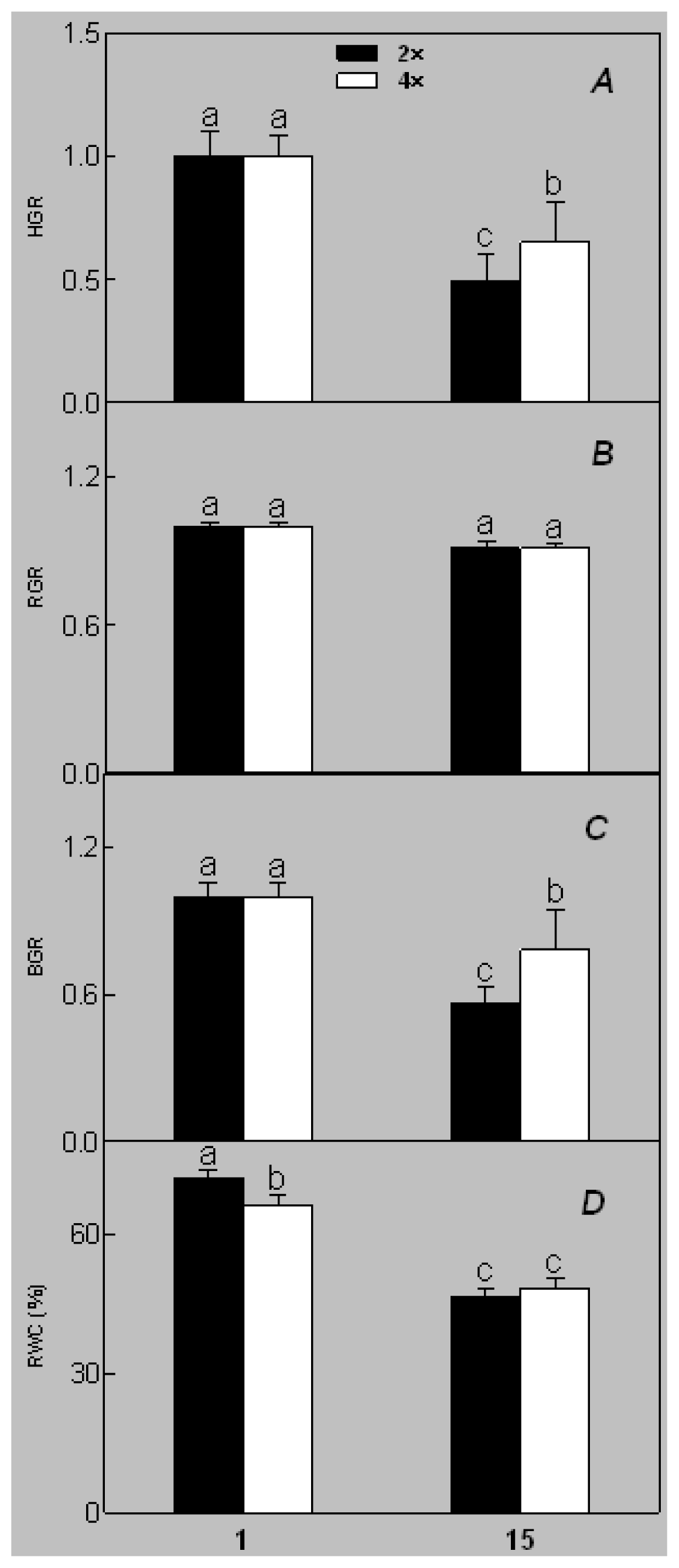
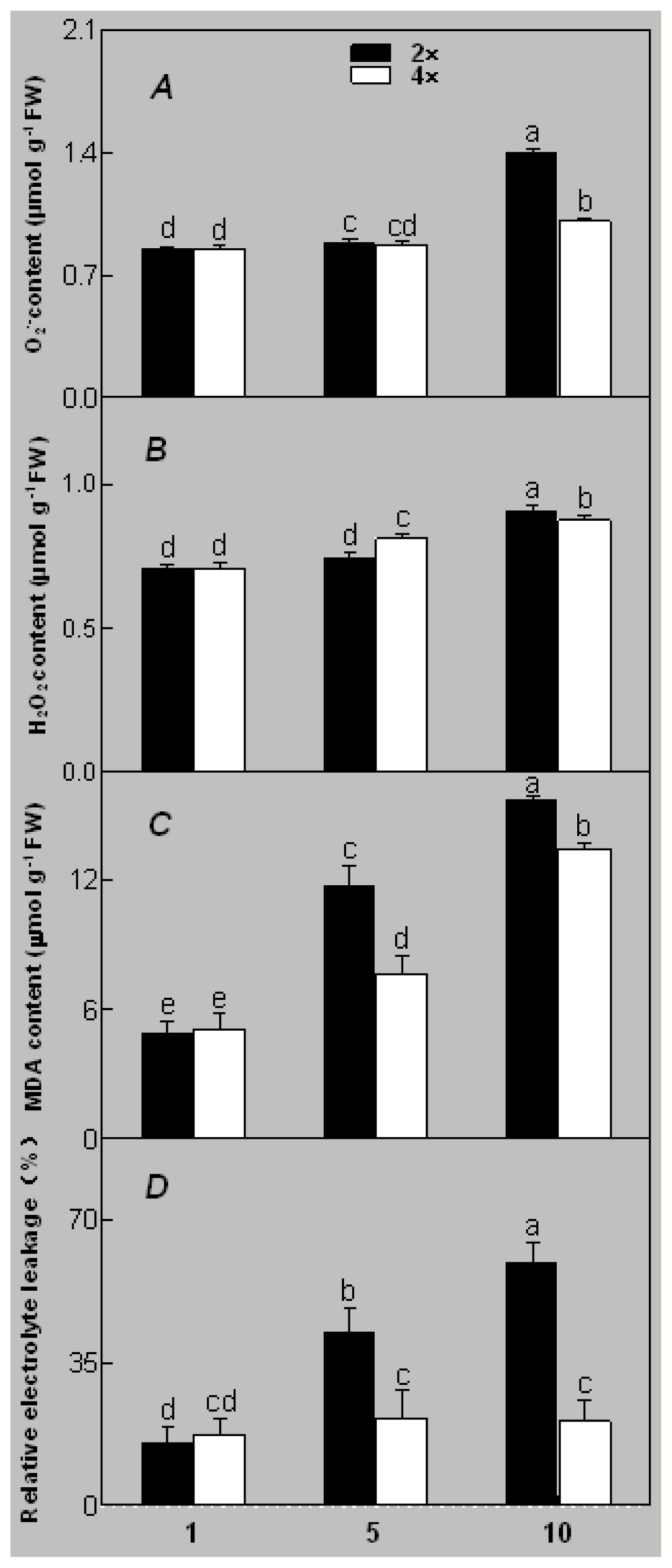
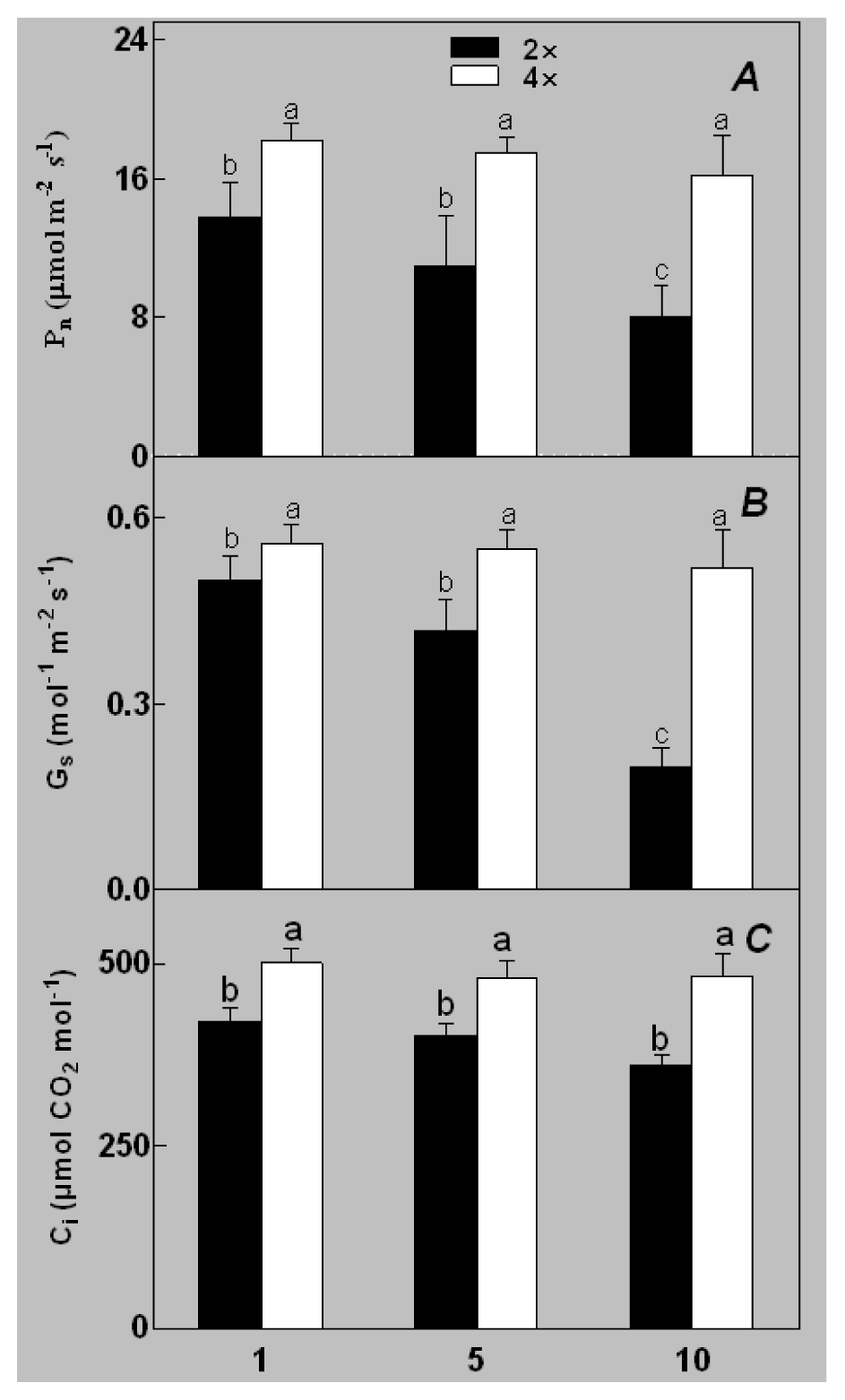
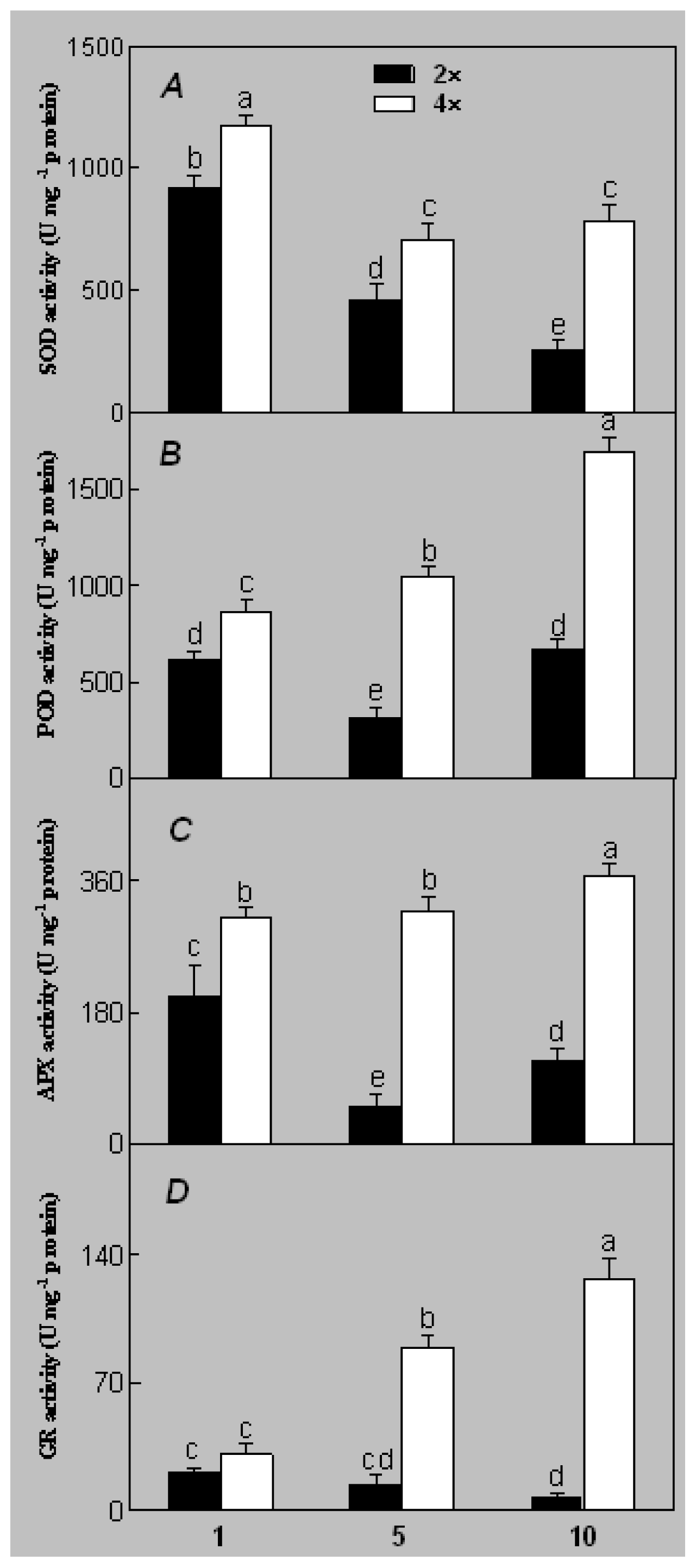
2.1. Plant Growth and Physiological Response of Leaves of 2× and 4× Plants to Salt Stress
2.2. Ultrastructural Responses of 2× and 4× Plants to Salt Stress
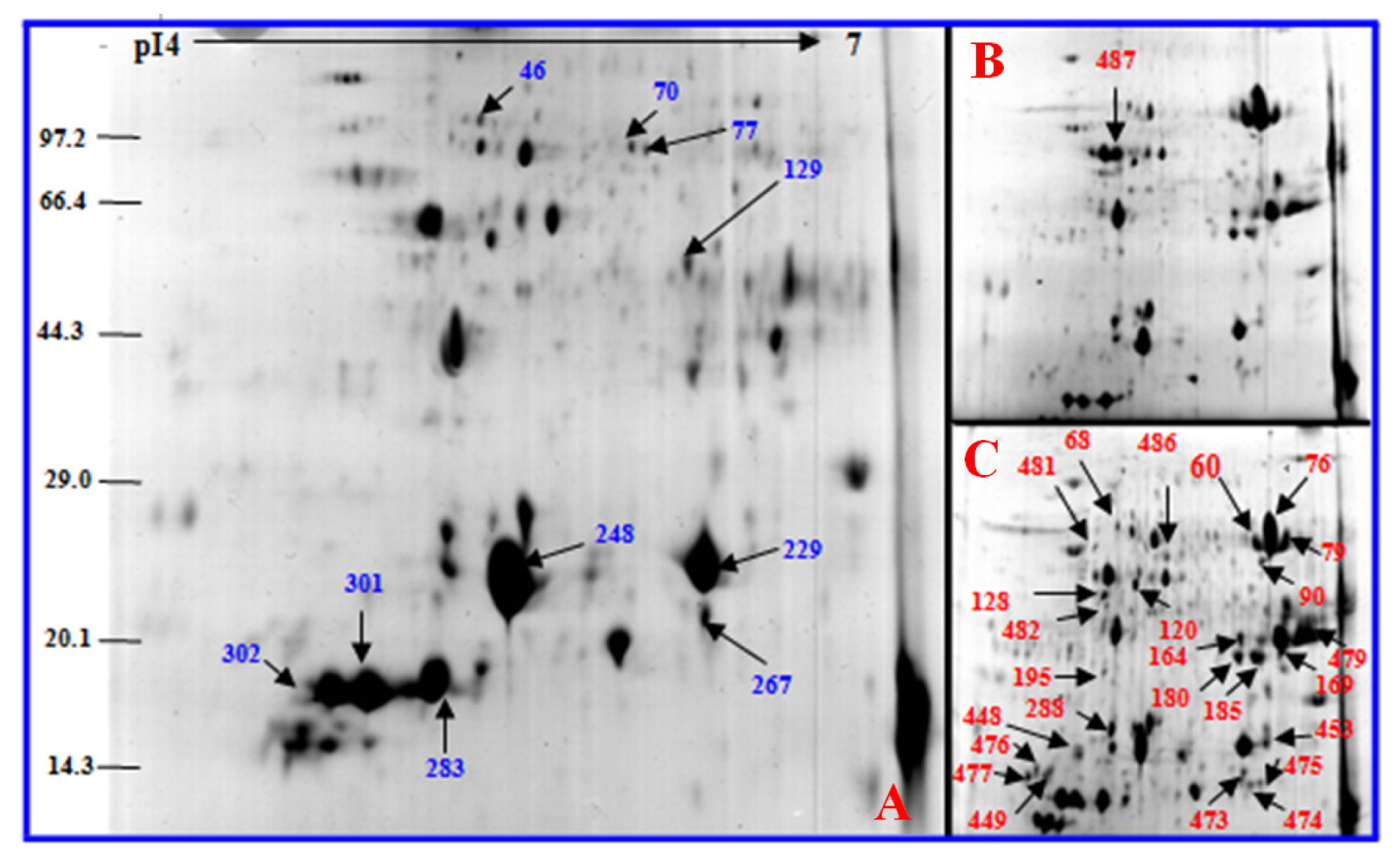
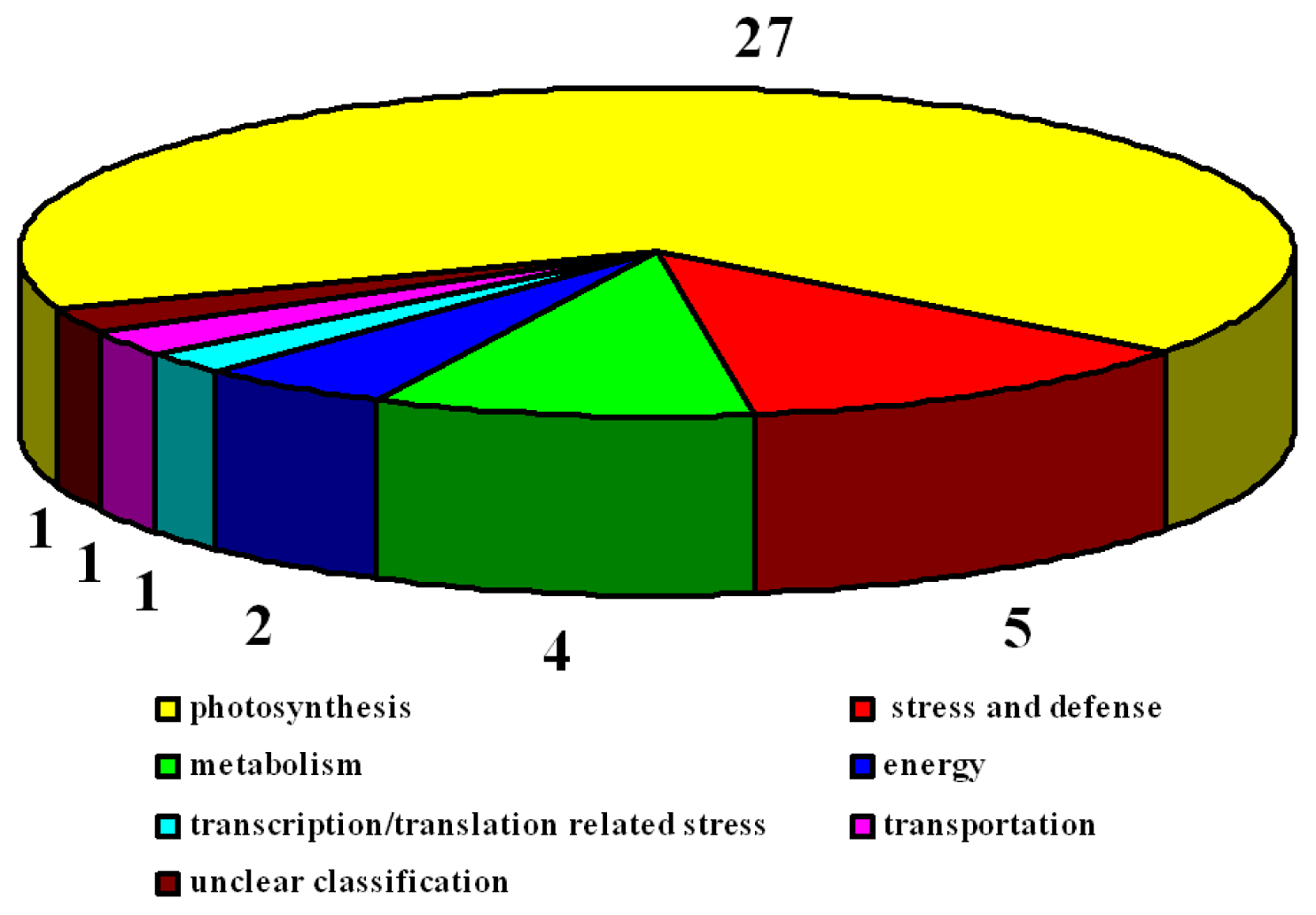
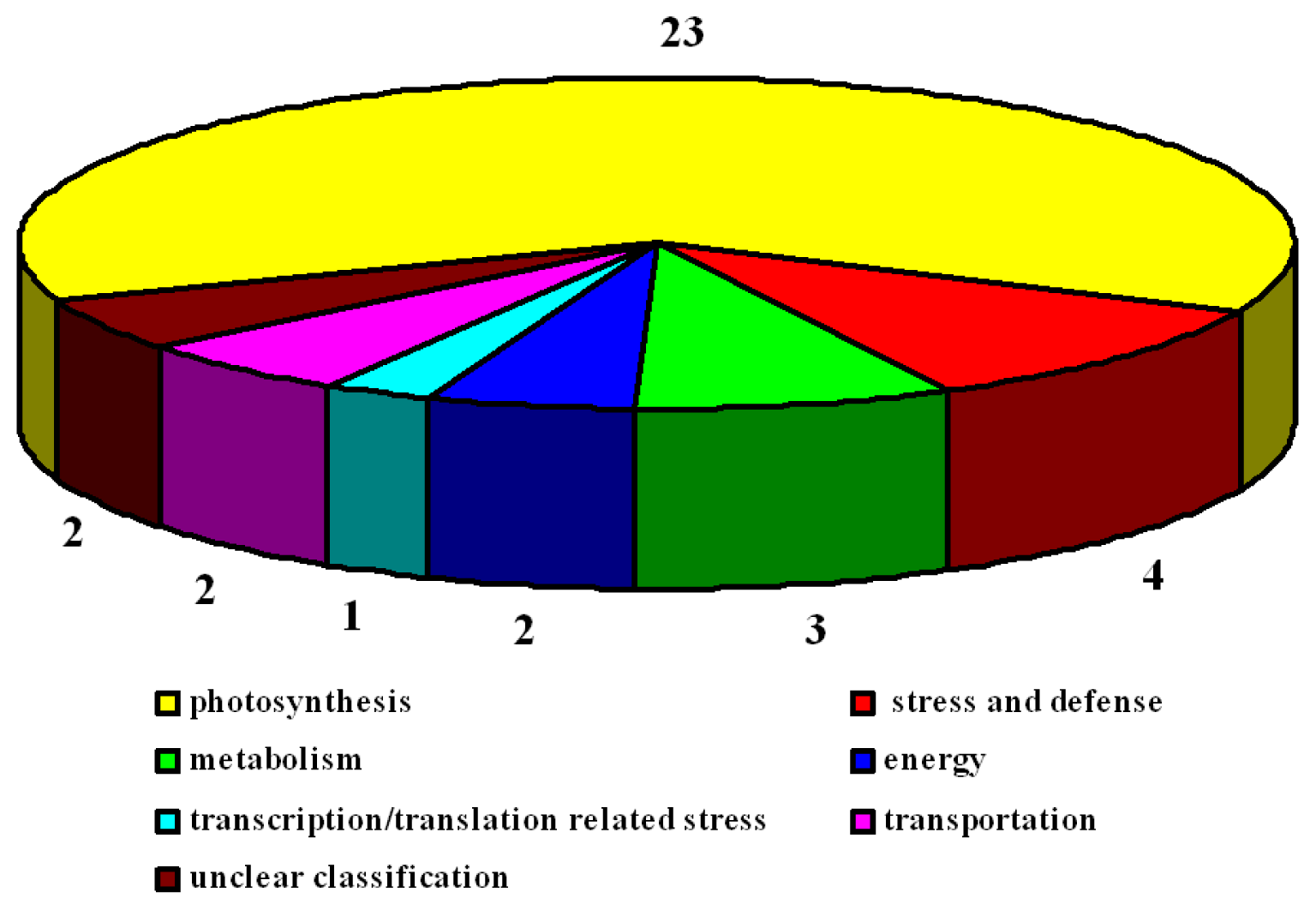
2.3. Identification of Differentially Expressed Proteins after Salt Treatment by Two-Dimensional Electrophoresis (2-DE)
2.3.1. Photosynthesis-Related Proteins
2.3.2. Stress and Defense Proteins
2.3.3. Energy Proteins
2.3.4. Metabolic Proteins
3. Experimental Section
3.1. Plant Materials and Growth Conditions
3.2. Morphological and Biomass Measurements
3.3. Ion Content Analysis
3.4. Determination of Superoxide Radical, Hydrogen Peroxide, Lipid Peroxidation and Relative Electrolyte Leakage
3.5. Photosynthesis Analysis
3.6. Antioxidant Enzyme Activity Analysis
3.7. Ultrastructural Leaf Analysis
3.8. Extraction of Total Leaf Protein
3.9. 2-DE Image Analysis and Gel Staining
3.10. In-Gel Digestion and Matrix-Assisted Time of Flight Mass Spectroscopy (MALDI-TOF-MS) Analysis
3.11. Statistical Analyses
4. Conclusions







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. a | Protein Name | Species | gi Number b | Theoretical MW(Da)/pI c | Experimental MW(Da)/pI d | Score e | M f | C (%) g | V% ± SE h (1/5/10 days) |
|---|---|---|---|---|---|---|---|---|---|
| Energy | |||||||||
| 494 | phosphoglycerate kinase | N. benthamiana | 313585890 | 50.05/7.66 | 58.00/6.34 | 172 | 10 | 19 | 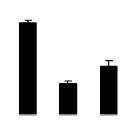 |
| 537 | phosphoglycerate kinase | R. pseudoacacia | 2257598 | 23.67/6.49 | 45.66/6.32 | 479 | 34 | 100 |  |
| Metabolism | |||||||||
| 498 | putative plastidic glutamine synthetase | O. sativa (japonica group) | 115461066 | 47.56/5.96 | 51.16/5.52 | 98 | 13 | 25 |  |
| 508 | chalcone synthase | R. pseudoacacia | 194740616 | 32.58/5.75 | 50.19/5.09 | 115 | 13 | 46 |  |
| 789 | aconitate hydratase domain protein | A. cellulolyticus CD2 | 303239527 | 19.84/4.86 | 28.47/4.45 | 91 | 11 | 58 |  |
| 536 | pyruvate kinase | C. reinhardtii | 159485206 | 23.201/6.2 | 40.20/6.22 | 86 | 23 | 15 |  |
| Photosynthesis | |||||||||
| 506 | Rubisco large subunit | G. subaequalis | 24634972 | 52.37/6.34 | 50.37/5.13 | 215 | 31 | 45 |  |
| 538 | Rubisco large subunit | Caesalpinia sp. SH-2010 | 306481385 | 43.78/7.38 | 45.71/6.67 | 310 | 32 | 49 | 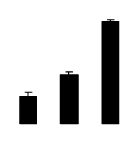 |
| 554 | Rubisco large subunit | Parkia multijuga | 148590322 | 51.78/6.23 | 34.27/6.41 | 190 | 19 | 27 |  |
| 557 | Rubisco large subunit | Merremia hastata | 21634009 | 47.96/6.42 | 32.77/6.30 | 216 | 18 | 25 |  |
| 575 | Rubisco large subunit | Dendrobium aphyllum | 300250366 | 22.77/6.2 | 22.67/5.39 | 217 | 13 | 54 |  |
| 582 | Rubisco large subunit | Loxocarya gigas | 5737828 | 49.48/6.43 | 21.50/5.29 | 258 | 11 | 26 |  |
| 584 | Rubisco large subunit | Ipomoea purpurea | 157325538 | 53.35/6.41 | 37.47/6. 26 | 182 | 18 | 31 |  |
| 593 | Rubisco large subunit | Millettia lenneoides | 18032763 | 51.53/6.04 | 24.01/6.22 | 239 | 18 | 24 |  |
| 603 | Rubisco large subunit | R. pseudoacacia | 2343004 | 49.20/6.13 | 19.78/5.88 | 336 | 31 | 54 |  |
| 610 | Rubisco large subunit | Haematoxylum brasiletto | 66735773 | 18.40/6.05 | 17.95/5.17 | 179 | 14 | 46 | 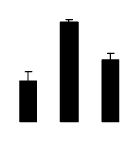 |
| 612 | Rubisco large subunit | Marila laxiflora | 49823207 | 21.44/5.91 | 18.190/4.98 | 282 | 12 | 35 |  |
| 615 | Rubisco large subunit | R.pseudoacacia | 340511916 | 50.20/6.14 | 18.69/5.28 | 305 | 32 | 53 |  |
| 621 | Rubisco large subunit | Prunus salicina | 15987094 | 51.31/6.99 | 16.61/4.30 | 147 | 14 | 20 |  |
| 622 | Rubisco large subunit | Cecropia palmata | 6983898 | 52.00/6.23 | 16.72/4.54 | 126 | 13 | 16 |  |
| 623 | Rubisco large subunit | R. pseudoacacia | 2342974 | 52.01/6.14 | 15.83/5.07 | 116 | 15 | 28 | 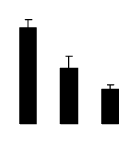 |
| 624 | Rubisco large subunit | Stigmaphyllon paralias | 14599610 | 52.24/6.23 | 15.23/4.41 | 175 | 21 | 31 | 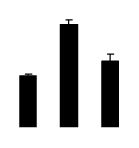 |
| 626 | Rubisco large subunit | Codonopsis dicentrifolia | 194400582 | 50.82/6.19 | 15.03/4.96 | 118 | 15 | 30 | 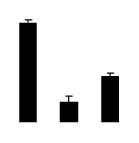 |
| 702 | Rubisco large subunit | Millettia lenneoides | 18032763 | 51.53/6.04 | 22.37/5.72 | 271 | 23 | 47 |  |
| 721 | Rubisco large subunit | Parthenocissus himalayana | 16973408 | 51.68/6.34 | 31.78/4.26 | 196 | 15 | 40 | 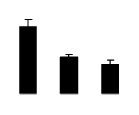 |
| 795 | Rubisco large subunit | R. pseudoacacia | 67079090 | 25.32/6.23 | 16.84/5.84 | 354 | 25 | 55 | 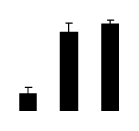 |
| 567 | Ribulose-bisphosphate carboxylase | Mangifera indica | 7261036 | 24.43/6.71 | 27.20/6.56 | 227 | 19 | 64 |  |
| 597 | Ribulose-bisphosphate carboxylase | Centrosema sp. SH-2010 | 306481395 | 49.81/6.44 | 22794/6.23 | 167 | 18 | 27 | 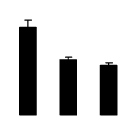 |
| 548 | Ribulose-biphosphate carboxylase oxygenase | Liparia genistoides | 146188483 | 27.28/6.36 | 37.36/6.06 | 263 | 25 | 60 | 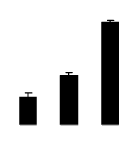 |
| 533 | photosystem II protein 33kD | Spinacia oleracea | 224916 | 26.65/5.01 | 38.28/5.34 | 294 | 22 | 78 | 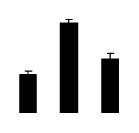 |
| 578 | polypeptide of the oxygen evolving complex of photosystem II | Sonneratia apetala | 146454492 | 24.99/5.61 | 24.59/5.03 | 95 | 6 | 26 | 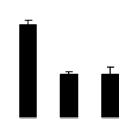 |
| 475 | ATP synthase CF1 alpha subunit | R. communis | 339516150 | 55.52/5.22 | 70.28/5.03 | 340 | 26 | 31 | 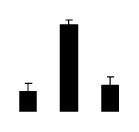 |
| 476 | putative ATP synthase beta subunit | O. sativa (japonica group) | 56784991 | 45.94/5.33 | 69.32/5.49 | 106 | 22 | 51 |  |
| Transportation | |||||||||
| 480 | aspartyl/glutamyl-tRNA(asn/gln) amidotransferase subunit b | Stigmatella aurantiaca DW4/3-1 | 310819540 | 53.54/5.56 | 59.82/6.02 | 74 | 11 | 23 |  |
| Transcription/translation related | |||||||||
| 625 | maturase-like protein | Coursetia weberbaueri | 23664381 | 60.78/9.42 | 15.06/4.94 | 193 | 18 | 39 | 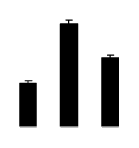 |
| Stress and defense | |||||||||
| 455 | heat shock protein 70 | Cucumis sativus | 1143427 | 75.37/5.15 | 100.32/4.83 | 332 | 21 | 25 |  |
| 478 | agglutinin I polypeptide B | R. pseudoacacia | 4033451 | 31.19/6.14 | 64.35/5.50 | 117 | 14 | 69 |  |
| 482 | agglutinin I polypeptide B | R. pseudoacacia | 4033451 | 31.19/6.14 | 60.65/6.19 | 488 | 19 | 69 | 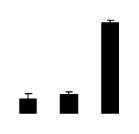 |
| 525 | plastidic aldolase | N. paniculata | 4827253 | 43.07/6.38 | 46.90/6.47 | 202 | 25 | 48 | 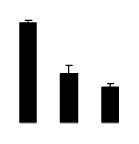 |
| 800 | phenylalanine ammonia lyase | R. pseudoacacia | 194740604 | 78.41/6.31 | 58.78/5.46 | 274 | 31 | 55 |  |
| Unclear classification | |||||||||
| 611 | predicted protein | Micromonas sp. RCC299 | 171910308 | 31.90/5.47 | 18.68/5.22 | 280 | 31 | 62 | 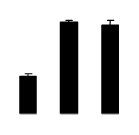 |
| Spot No. a | Protein Name | Species | gi Number b | Theoretical MW(Da)/pI c | Experimental MW(Da)/pI d | Score e | M f | C (%) g | V% ± SE h (1/5/10 days) |
|---|---|---|---|---|---|---|---|---|---|
| Energy | |||||||||
| 60 | phosphoglycerate kinase | N. benthamiana | 313585890 | 50.05/7.66 | 98.76/6.24 | 169 | 13 | 23 | 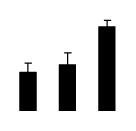 |
| 476 | mitochondrial F1-ATPase beta subunit | Dimocarpus longan | 269914683 | 59.87/6.18 | 22.47/4.62 | 274 | 23 | 38 | 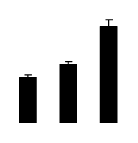 |
| Metabolism | |||||||||
| 229 | chalcone synthase | R. pseudoacacia | 194740620 | 36.76/6.22 | 27.61/6.24 | 608 | 29 | 32 |  |
| 486 | chalcone synthase | R. pseudoacacia | 194740616 | 32.58/5.75 | 68.39/5.71 | 175 | 15 | 48 | 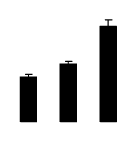 |
| 70 | enolase | Glycine max | 42521309 | 47.69/5.31 | 92.57/5.73 | 130 | 12 | 20 |  |
| Photosynthesis | |||||||||
| 79 | Rubisco large subunit | R. pseudoacacia | 340511916 | 50.71/6.14 | 98.81/6.59 | 299 | 34 | 62 |  |
| 90 | Rubisco large subunit | Daviesia rhizomata | 18032753 | 51.67/6.14 | 67.60/6.28 | 418 | 37 | 63 | 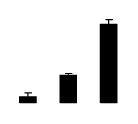 |
| 164 | Rubisco large subunit | Gironniera subaequalis | 24634972 | 52.37/6.34 | 44.69/6.22 | 340 | 35 | 47 | 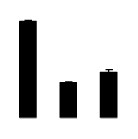 |
| 180 | Rubisco large subunit | Wisteria sp. | 2343020 | 51.49/6.13 | 39.61/6.19 | 294 | 20 | 32 | 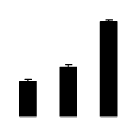 |
| 185 | Rubisco large subunit | Canavalia rosea | 18157259 | 52.11/6.14 | 39.79/6.24 | 209 | 17 | 30 |  |
| 267 | Rubisco large subunit | R. pseudoacacia | 2343004 | 49.23/6.13 | 22.59/6.28 | 156 | 20 | 47 | 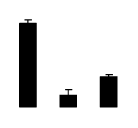 |
| 301 | Rubisco large subunit | Mascagnia stannea | 14599586 | 51.57/6.14 | 19.08/4.81 | 347 | 29 | 56 | 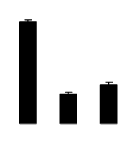 |
| 302 | Rubisco large subunit | Mascagnia stannea | 14599586 | 51.57/6.14 | 18.99/4.64 | 308 | 31 | 54 |  |
| 448 | Rubisco large subunit | Floerkea proserpinacoides | 38147280 | 51.57/5.87 | 24.53/4.89 | 120 | 14 | 23 |  |
| 449 | Rubisco large subunit | Aspicarpa sericea | 331690047 | 49.79/6.13 | 22.68/4.81 | 175 | 16 | 27 | 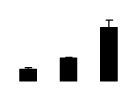 |
| 453 | Rubisco large subunit | Wisteria sp. | 2343020 | 51.49/6.13 | 27.71/6.32 | 311 | 26 | 42 | 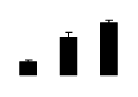 |
| 473 | Rubisco large subunit | Centrosema sp. SH-2010 | 306481395 | 49.81/6.44 | 22.49/6.20 | 265 | 27 | 31 | 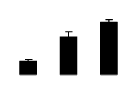 |
| 474 | Rubisco large subunit | Sassafras albidum | 283558279 | 20.64/6.05 | 21.07/6.24 | 132 | 14 | 32 |  |
| 477 | Rubisco large subunit | Diospyros pentamera | 221078519 | 51.98/6.14 | 21.06/4.35 | 179 | 15 | 20 |  |
| 479 | Rubisco large subunit | Mucuna macrocarpa | 18157295 | 52.11/6.14 | 44.05/6.82 | 284 | 33 | 43 |  |
| 272 | Rubisco large subunit | Pachynema junceum | 9909908 | 19.07/5.33 | 20.07/5.68 | 226 | 14 | 36 | 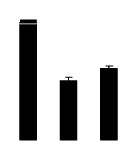 |
| 283 | Rubisco large subunit | Pachynema junceum | 9909908 | 19.19/5.33 | 19.14/5.18 | 199 | 14 | 36 | 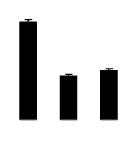 |
| 481 | Rubisco activase | Glycine max | 290766481 | 52.64/5.54 | 98.57/5.11 | 211 | 29 | 43 |  |
| 487 | Rubisco activase | Glycine max | 290766485 | 48.64/6.28 | 53.18/5.47 | 172 | 14 | 21 |  |
| 129 | Rubisco activase | Zantedeschia aethiopica | 13430334 | 37.25/6.7 | 55.76/6.03 | 157 | 18 | 35 | 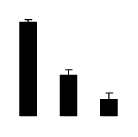 |
| 68 | ATP synthase CF1 alpha subunit | R. communis | 339516150 | 55.52/5.22 | 93.78/5.40 | 472 | 28 | 36 | 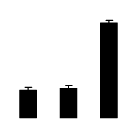 |
| 475 | ATP synthase CF1 alpha subunit | Vigna radiata | 289066833 | 55.68/5.21 | 21.06/6.24 | 366 | 28 | 35 | 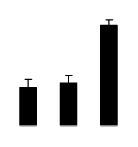 |
| 120 | Phosphoribulokinase (PPK) | Pisum sativum | 1885326 | 39.00/5.41 | 58.67/5.57 | 129 | 10 | 25 | 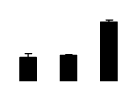 |
| Transportation | |||||||||
| 46 | Rubisco subunit binding-protein beta subunit | R. communis | 255564820 | 64.15/5.65 | 100.08/5.40 | 127 | 13 | 23 | 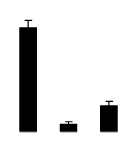 |
| 248 | General secretion pathway protein D precursor, putative | R. communis | 255619353 | 30.81/4.67 | 27.01/5.53 | 174 | 17 | 54 |  |
| Transcription/translation related | |||||||||
| 77 | maturase-like protein | Olneya tesota | 23477700 | 60.67/9.45 | 91.49/6.01 | 409 | 27 | 52 | 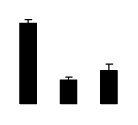 |
| Stress and defense | |||||||||
| 76 | Chain A, legume lectin sf the bark of robinia pseudoacacia | R. pseudoacacia | 15826665 | 25.58/4.48 | 99.77/6.42 | 121 | 13 | 57 | 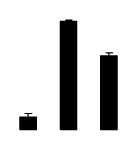 |
| 128 | sedoheptulose-1,7-bisphosphatase(SBPase) | Cucumis sativus | 229597543 | 42.08/5.96 | 55.09/5.22 | 99 | 11 | 23 | 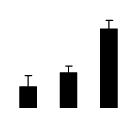 |
| 169 | phenylalanine ammonia lyase | R. pseudoacacia | 194740606 | 34.40/5.4 | 44.64/6.59 | 300 | 21 | 72 | 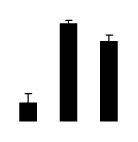 |
| 195 | ascorbate peroxidase | Medicago sativa | 16304410 | 20.14/5.33 | 32.49/5.26 | 110 | 6 | 27 | 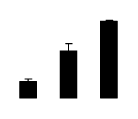 |
| Unclear classification | |||||||||
| 482 | PREDICTED: ADP-ribosylation factor 1-like | Amphimedon queenslandica | 340369230 | 20.47/6.15 | 49.38/5.26 | 89 | 2 | 4 | 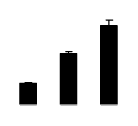 |
| 288 | Em protein | R. pseudoacacia | 1754977 | 12.22/6.21 | 28.51/5.30 | 356 | 22 | 98 | 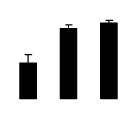 |
Acknowledgments
Conflicts of Interest
Abbreviations
| APX | ascorbate peroxidase |
| BGR | the relative growth rate of stem basal diameter |
| BSA | bovine serum albumin |
| CHS | chalcone synthase |
| 2-DE | Two-dimensional electrophoresis |
| HGR | The relative growth rate of height |
| H2O2 | hydrogen peroxide |
| Hsp70 | heat shock protein 70 |
| MDA | Malondialdehyde |
| PGK | phosphoglycerate kinase |
| PPK | phosphoribulokinase |
| RGR | the relative growth rate of range |
| ROS | Reactive Oxygen Species |
| RuBPCase | ribulose-bisphosphate carboxylase |
| RWC | relative water content |
| SBPase | sedoheptulose-1,7-bisphosphatase |
| TCA | tricarboxylic acid |
References
- Horie, T.; Schroeder, J.I. Sodium transporters in plants. Diverse genes and physiological functions. Plant Physiol 2004, 136, 2457–2462. [Google Scholar]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci 2001, 6, 66–71. [Google Scholar]
- Srivastava, A.K.; Ramaswamy, N.K.; Mukopadhyaya, R.; Chiramal, J.M.G.; D’Souza, S.F. Thiourea modulates the expression and activity profile of mtATPase under salinity stress in seeds of Brassica juncea. Ann. Bot 2009, 103, 403–410. [Google Scholar]
- Yang, A.; Dai, X.; Zhang, W.H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot 2012, 63, 2541–2556. [Google Scholar]
- Breyne, P.; Zabeau, M. Genome-wide expression analysis of plant cell cycle modulated genes. Curr. Opin. Plant Biol 2001, 4, 136–142. [Google Scholar]
- Salekdeh, G.H.; Siopongco, J.; Wade, L.J.; Ghareyazie, B.; Bennett, J. Proteomic analysis of rice leaves during drought stress and recovery. Proteomics 2002, 2, 1131–1145. [Google Scholar]
- Zhang, H.; Han, B.; Wang, T.; Chen, S.; Li, H.; Zhang, Y.; Dai, S. Mechanisms of plant salt response: Insights from proteomics. J. Proteome Res 2012, 11, 49–67. [Google Scholar]
- Romero-Aranda, R.; Bondada, B.R.; Syvertsen, J.P.; Grosser, J.W. Leaf characteristics and net gas exchange of diploid and autotetraploid citrus. Ann. Bot 1997, 79, 153–160. [Google Scholar]
- Wang, Q.L.; Yu, M.D.; Lu, C.; Wu, C.R.; Jing, C.R. Study on breeding and photosynthetic characteristics of new polyploidy variety for leaf and fruit-producing mulberry (Morus L). Sci. Agric. Sin 2011, 44, 562–569. [Google Scholar]
- Masterson, J. Stomatal size in fossil plants: Evidence for polyploidy in majority of angiosperms. Science 1994, 264, 421–424. [Google Scholar]
- Stupar, R.M.; Bhaskar, P.; Yandell, B.; Rensink, W.A.; Hart, A.L.; Ouyang, S.; Veilleux, R.E.; Busse, J.S.; Erhardt, R.J.; Buell, C.R.; et al. Phenotypic and transcriptomic changes associated with potato autopolyploidization. Genetics 2007, 176, 2055–2067. [Google Scholar]
- Riddle, N.C.; Jiang, H.; An, L.; Doerge, R.W.; Birchler, J.A. Gene expression analysis at the intersection of ploidy and hybridity in maize. Theor. Appl. Genet 2010, 120, 341–353. [Google Scholar]
- Ramsey, J. Polyploidy and ecological adaptation in wild yarrow. Proc. Natl. Acad. Sci. USA 2011, 108, 6697–6669. [Google Scholar]
- Li, F.Z.; Ning, X.M.; Qiu, X.M.; Su, C.F.; Yao, J.Q.; Tian, L.W. Genetic mapping of the dark brown fiber Lc1 gene in tetraploid cotton. Sci. Agric. Sin 2012, 45, 4109–4114. [Google Scholar]
- Allario, T.; Brumos, J.; Colmenero-Flores, J.M.; Tadeo, F.; Froelicher, Y.; Talon, M.; Navarro, L.; Ollitrault, P.; Morillon, R. Large changes in anatomy and physiology between diploid Rangpur lime (Citrus limonia) and its autotetraploid are not associated with large changes in leaf gene expression. J. Exp. Bot 2013, 62, 2507–2519. [Google Scholar]
- Podda, A.; Checcucci, G.; Mouhaya, W.; Centeno, D.; Rofidal, V.; del Carratore, R.; Luro, F.; Morillon, R.; Ollitrault, P.; Maserti, B.E. Salt-stress induced changes in the leaf proteome of diploid and tetraploid mandarins with contrasting Na+ and Cl− accumulation behaviour. J. Plant Physiol 2013, 170, 1101–1112. [Google Scholar]
- Beest, M.; le Roux, J.J.; Richardson, D.M.; Brysting, A.K.; Suda, J.; Kubešová, M.; Pyšek, P. The more the better? The role of polyploidy in facilitating plant invasions. Ann. Bot 2012, 109, 19–45. [Google Scholar]
- Lu, C.; Cui, B.; Huang, L.; Sun, P.; Zhang, G.; Li, Y. Phenotypic observation and analysis of inflorescence variation of Autotetraploid Robinia pseudoacacia. Sci. Silvae Sin 2012, 48, 63–68. [Google Scholar]
- Al, H.A.; Monneveaux, P.; Nachit, M.M. Direct and indirect selection for drought tolerance in alien tetraploid wheat durum wheat crosses. Euphytica 1998, 100, 287–294. [Google Scholar]
- Fock, I.; Collonnier, C.; Purwito, A.; Luisetti, J.; Souvannavong, V.; Vedel, F.; Servaes, A.; Ambroise, A.; Kodja, H.; Ducreux, G.; et al. Resistance to bacterial wilt in somatic hybrids between Solanum tuberosum and Solanum phureja. Plant Sci 2000, 160, 165–176. [Google Scholar]
- Huang, S.; Sirikhachornkit, A.; Su, X.; Faris, J.; Gill, B.S.; Haselkorn, R.; Gornicki, P. Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of the Triticum/Aegilops complex and the evolutionary history of polyploid wheat. Proc. Natl. Acad. Sci. USA 2002, 99, 8133–8138. [Google Scholar]
- Zhang, X.Y.; Hu, C.G.; Yao, J.L. Tetraploidization of diploid Dioscorea results in activation of the antioxidant defense system and increased heat tolerance. J. Plant Physiol 2010, 167, 88–94. [Google Scholar]
- Xiong, Y.C.; Li, F.M.; Zhang, T. Performance of wheat crops with different chromosome ploidy: Root-sourced signals, drought tolerance, and yield performance. Planta 2006, 224, 710–718. [Google Scholar]
- Meng, F.J.; Huang, F.L. Changes of function and ultrastructure of mitochondria in Robinia pseudoacacia leaves under salt stress. Nonwood For. Res 2010, 28, 18–23. [Google Scholar]
- Saleh, B.; Allario, T.; Dambier, D.; Ollitrault, P.; Morillon, R. Tetraploid citrus rootstocks are more tolerant to salt stress than diploid. C. R. Biol 2008, 331, 703–710. [Google Scholar]
- Xiao, X.; Yang, F.; Zhang, S.; Korpelainen, H.; Li, C. Physiological and proteomic responses of two contrasting Populus cathayana populations to drought stress. Physiol. Plant 2009, 136, 150–168. [Google Scholar]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J 2006, 45, 523–539. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Tuna, A.L.; Kaya, C.; Dikilitas, M.; Higgs, D. The combined effects of gibberellid acid and salinity on some antioxidant enzyme activities, plant growth parameters and nutritional status in maize plants. Environ. Exp. Bot 2008, 62, 1–9. [Google Scholar]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv 2009, 27, 84–93. [Google Scholar]
- Barhoumi, Z.; Djebali, W.; Chaïbi, W.; Abdelly, C.; Smaoui, A. Salt impact on photosynthesis and leaf ultrastructure of Aeluropus littoralis. J. Plant Res 2007, 120, 529–537. [Google Scholar]
- Zapata, J.M.; Guéra, A.; Esteban-Carrasco, A.; Martín, M.; Sabater, B. Chloroplasts regulate leaf senescence: Delayed senescence in transgenic ndhF-defective tobacco. Cell Death Differ 2005, 12, 1277–1284. [Google Scholar]
- Ishikawa, T.; Yoshimura, K.; Tamoi, M.; Takeda, T.; Shigeoka, S. Alternative mRNA splicing of 3′-terminal exons generates ascorbate peroxidase isoenzymes in spinach (Spinacia oleracea) chloroplasts. Biochem. J 1997, 328, 795–800. [Google Scholar]
- Ndimba, B.K.; Chivasa, S.; Simon, W.J.; Slabas, A.R. Identification of Arabidopsis salt and osmotic stress responsive proteins using two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 4185–4196. [Google Scholar]
- Portis, J.A.R. Rubisco activase: Rubisco’s catalytic chaperone. Photosynth. Res 2003, 751, 11–27. [Google Scholar]
- Parker, R.; Flowers, T.J.; Moore, A.L.; Harpham, N.V.J. An accurate and reproducible method for proteome profiling of the effects of salt stress in the rice leaf lamina. J. Exp. Bot 2006, 57, 1109–1118. [Google Scholar]
- Sudhir, P.R.; Pogoryelov, D.; Kovacs, L.; Garab, G.; Murthy, S.D. The effects of salt stress on photosynthetic electron transport and thylakoid membrane proteins in the cyanobacterium Spirulina platensis. J. Biochem. Mol. Biol 2005, 38, 481–485. [Google Scholar]
- Ma, H.; Song, L.; Shu, Y.; Wang, S.; Niu, J.; Wang, Z.; Yu, T.; Gu, W.; Ma, H.; Baker, N.R. Comparative proteomic analysis of seedling leaves of different salt tolerant soybean genotypes. J. Proteomics 2012, 75, 1529–1546. [Google Scholar]
- Davletova, S.; Rizhsky, L.; Liang, H.; Sheng, A.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic ascorbate peroxidase 1 is a central component of the reactive oxygen gene network of Arabidopsisi. Plant Cell 2005, 17, 268–281. [Google Scholar]
- Shakirova, F.M.; Bezrukova, M.V.; Khairullin, R.M. The increase in lectin level in wheat shoots under the action of salt stress. Izv. Russ. Acad. Sci 1993, 1, 142–145. [Google Scholar]
- Kosová, K.; Vítámvás, P.; Prášil, L.T.; Renaut, J. Plant proteome changes under abiotic stress-contribution of proteomics studies to understanding plant stress response. J. Proteomics 2011, 74, 1301–1322. [Google Scholar]
- Wolf, B. A comprehensive system of leaf analyses and its use fore diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal 1982, 13, 1035–1059. [Google Scholar]
- Chen, S.; Li, J.; Wang, S.; Hüttermann, A.; Altman, A. Salt, nutrient uptake and transport, and ABA of Populus euphratica; a hybrid in response to increasing soil NaCl. Trees 2001, 15, 186–194. [Google Scholar]
- Roth, E.F.; Gilbert, J.H.S. Pyrogallol assay for SOD: Absence of a glutathione artifact. Anal. Biochem 1984, 137, 50–53. [Google Scholar]
- Nickel, R.S.; Cunningham, B.A. Improved peroxidase assay method using leuco 2,3,6-trichloroindophenol and application to comparative measurements of peroxidase catalysis. Anal. Biochem 1969, 27, 292–299. [Google Scholar]
- Nordhoff, A.; Bucheler, U.S.; Werner, D.; Schirmer, R.H. Folding of the four domains and dimerization are impaired by the Gly446→Glu exchange in human glutathione reductase. implications for the design of antiparasitic drugs. Biochemistry 1993, 32, 4060–4066. [Google Scholar]
- Nakano, Y.; Asad, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplast. Plant Cell Physiol 1981, 22, 867–880. [Google Scholar]
- Hurkman, W.J.; Tanaka, C.K. Solubilization of plant membrane proteins for analysis by two-dimensional gel electrophoresis. Plant Physiol 1986, 81, 802–806. [Google Scholar]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-due binding. Ann. Biochem 1976, 72, 248–254. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Z.; Wang, M.; Liu, L.; Meng, F. Physiological and Proteomic Responses of Diploid and Tetraploid Black Locust (Robinia pseudoacacia L.) Subjected to Salt Stress. Int. J. Mol. Sci. 2013, 14, 20299-20325. https://doi.org/10.3390/ijms141020299
Wang Z, Wang M, Liu L, Meng F. Physiological and Proteomic Responses of Diploid and Tetraploid Black Locust (Robinia pseudoacacia L.) Subjected to Salt Stress. International Journal of Molecular Sciences. 2013; 14(10):20299-20325. https://doi.org/10.3390/ijms141020299
Chicago/Turabian StyleWang, Zhiming, Mingyue Wang, Likun Liu, and Fanjuan Meng. 2013. "Physiological and Proteomic Responses of Diploid and Tetraploid Black Locust (Robinia pseudoacacia L.) Subjected to Salt Stress" International Journal of Molecular Sciences 14, no. 10: 20299-20325. https://doi.org/10.3390/ijms141020299