Characterization of 42 Microsatellite Markers from Poison Ivy, Toxicodendron radicans (Anacardiaceae)

Abstract
:1. Introduction
2. Results and Discussion
2.1. Enrichment Microsatellite Library and Sequencing Results
2.2. Development of Microsatellite Markers
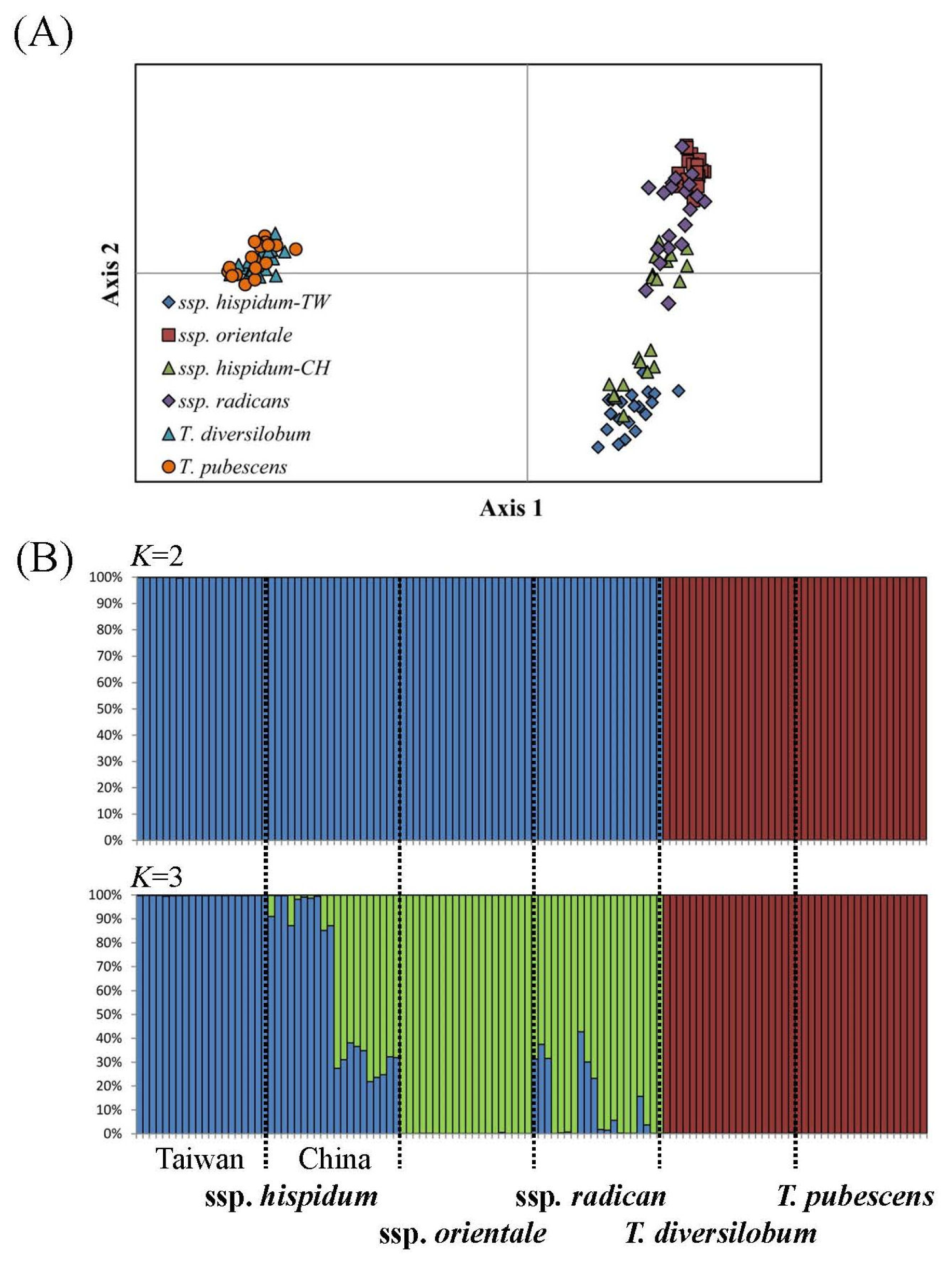
2.3. Genotyping and Population Genetics Analysis
3. Experimental Section
3.1. Sampling and DNA Extractions
3.2. Isolation of Microsatellite DNA Loci and Identification
3.3. DNA Amplification and Genotyping
3.4. Data Analysis
4. Conclusions


{kind=link}
| Locus | Primer sequence (5′–3′ ) | Repeat motif | Allelic size (bp) | Ta (°C) | GenBank Accession No. |
|---|---|---|---|---|---|
| AC3 | F: GCGCAAATACGAAAGCGAGA | (AG)27 | 104~146 | 55 | HF680270 |
| R: AAAAATGGGCTCAAGCGATC | |||||
| AC6 | F: CGGGATCGATGATGAGTCCTGA | (ATT)7(TTC)2N(CTT)13 | 299~337 | 55 | HF680271 |
| R: ATCAGAGGAGCGAGTCAGC | |||||
| AC11 | F: GTGAAGAAACTGAAGAGCCAC | (AG)24 | 194~218 | 55 | HF680272 |
| R: TCACCAAAACTTAAGGGTGG | |||||
| AC19 | F: CCACTCCACCCGTAACAACG | (AGAAAA)5N(CT)14N(ATG)7 | 324~340 | 55 | HF680273 |
| R: TCGTCCGTCATCGCTGCCCT | |||||
| AC20 | F: CGTGCGTTACTTCTGCTCAC | (ATG)12(AAG)9(ATG)9 | 237~245 | 55 | HF680274 |
| R: ACTGTGAATCACCTGACCACG | |||||
| AC139 | F: GAGGTGATATTGGTACTTGG | (TA)9(GA)10 | 112~128 | 55 | HF680275 |
| R: TTCCTCTCACTTTTACGTTC | |||||
| AG28 | F: TATCGCATCAGGGGTTCCCA | (GGA)15 | 222~230 | 55 | HF680276 |
| R: CGGGATGGAGCCGCCAATGA | |||||
| AG153 | F: GATGAGTCCTGAGTAAACCA | (TTTC)19 | 165–169 | 51 | HF680277 |
| R: TGCATATTTCATGATAATGG | |||||
| M8 | F: TTCTTCTTCATTGTGCCGTC | (GA)23 | 136~140 | 55 | HF680278 |
| R: ATGTAGGCATGAATGAGGTG | |||||
| M18 | F: AGGCTCCAAATCCATGCCTC | (AAGA)27 | 187~195 | 55 | HF680279 |
| R: CAAGAGCAAGAACATAGAATATAA | |||||
| M19 | F: AGTGAATAGGTAGAATTCTCC | (AG)22 | 129~129 | 55 | HF680280 |
| R: CGGATTTTAGCTCAATTCCATC | |||||
| M22 | F: AAGGATCAAGAAGGAAGGTG | (AG)30 | 155~159 | 55 | HF680281 |
| R: CCCTTCTCTTTCTTCTTCCC | |||||
| M24 | F: GATTCATCTGGGTCACCTGG | (GAGTGA)14 | 166~178 | 55 | HF680282 |
| R: GACAATAGACTCCGACAACG | |||||
| M27 | F: CATTCTTCTTCATTGTGCCG | (GA)27 | 110~112 | 55 | HF680283 |
| R: CCAATTTACCGAATCCAAGC | |||||
| M30 | F: AAAGTTCATCATGGGTGTTTG | (TG)16 | 124~148 | 55 | HF680284 |
| R: AAACAAATCAGCCCTTCCAC | |||||
| M31 | F: AGTTGTGTATGTCTGTGTTG | (GT)92 | 218~244 | 55 | HF680285 |
| R: AAACAAAGATGATGTAAAACGC | |||||
| M452 | F: GACCAAGTGAAGCTGAATAG | (GA)12 | 75~105 | 55 | HF680286 |
| R: CTCACCAACTCAGCTAAGC | |||||
| M493 | F: GCATCCTTCATTTTCTTATGG | (AAGA)25 | 221~223 | 55 | HF680287 |
| R: CGTTATCCAAACAACTCCAC | |||||
| M54 | F: AAAACGTTAGCCGATAAGG | (GA)15 | 108~132 | 55 | HF680288 |
| R: TCAGCCTCTCCCCTCTTTTC | |||||
| M56 | F: TGGAGATGGAGATGAAGAGG | (AG)12 | 93~123 | 55 | HF680289 |
| R: GCGTAAGATAGTCACTGTAC | |||||
| M60 | F: AACTGAAGAGGTGCAATGGG | (TGA)17 | 122~144 | 55 | HF680290 |
| R: AGAGACTCTTCATCTTCTCC | |||||
| M61 | F: CCGTTCACTGATTTTGCTAG | (AG)11 | 169~207 | 55 | HF680291 |
| R: CTGGCTACTAGATGATCCAG | |||||
| M64 | F: ATAGTGAGTGCATGGTGGCG | (AG)17 | 114~128 | 55 | HF680292 |
| R: CTCCTCTTGAAACTGAGCTG | |||||
| M66 | F: TGGAGCACTCATTTGTAACG | (AG)11N(AG)9N(AG)9 | 116~132 | 55 | HF680293 |
| R: CTGGATCTATACTCAATTCC | |||||
| M67 | F: AGTGTGCTCTAAGAGTAAGG | (GAAT)14 | 153 | 55 | HF680294 |
| R: TATCCTACTAGGACTCTACC | |||||
| M68 | F: CTGGTGTTGGGAAAGAAGG | (TGGTGA)10 | 120 | 51 | HF680295 |
| R: TTATTACCATATTATCCTTTACAT | |||||
| M821 | F: TTGTCATCGTCGTCCAAACC | (TTG/A)11 | 158~160 | 55 | HF680296 |
| R: AAATCTCCTCATCCAACGCC | |||||
| M822 | F: GGTGGATTGAAGAAATGACG | (GA)4(GAGAA)4N(GA)12 | 127~149 | 55 | HF680297 |
| R: AAATTCATTCGCTTTCACCTC | |||||
| M83 | F: CATTCAACGCCGACAATTCC | (AAT/C)16 | 124~126 | 55 | HF680298 |
| R: TCCATATTCAGCCCAAGTGC | |||||
| M85 | F: TTTGCTTTGGTTGAGAGTGC | (AG)11 | 118~122 | 55 | HF680299 |
| R: AATGTAATGTTCCTCCAACG | |||||
| M97 | F: AGTTCTGGAGCTCAACATGG | (GT)12 | 163~179 | 55 | HF680300 |
| R: TCGAAGCTCTGATACCACTG | |||||
| M99 | F: CCTTCCGGAGAGGTAGATTG | (AG)10 | 140~152 | 55 | HF680301 |
| R: TCTATAAGTACACCTTCTCC | |||||
| M104 | F: TGGATTAGGCGAGTCACACC | (AG)15 | 149~157 | 55 | HF680302 |
| R: GTTTCACAGCATCCACGTGC | |||||
| M120 | F: CGACTCATAATTGACGAGCC | (TG)10 | 119~143 | 55 | HF680303 |
| R: CTGTAAAATTACTATAGCCC | |||||
| M121 | F: TGATTCTTTTGTGGTTTGCG | (AG)14 | 210~216 | 55 | HF680304 |
| R: TGTGTAGTGATTATAGAAGG | |||||
| M123 | F: GTAATGTGTTTCAGTGCGTC | (AG)12 | 138~154 | 55 | HF680305 |
| R: CTTTTGGGCTATCATGGATG | |||||
| M124 | F: AAGTACAGTTCCCGAAACTG | (AAAG)10N(AG)11 | 296~320 | 55 | HF680306 |
| R: TATTTTCACTAACCCTACCC | |||||
| M137 | F: AGTGAGCTATCCAGCTATCG | (AG)22 | 124 | 52 | HF680307 |
| R: TCGTGTCAGTTTCGAGTAGC | |||||
| M148 | F: GATCTGAATTTTCCGAAAGCG | (AG)10 | 197 | 53 | HF680308 |
| R: AGTGGGAGTTACAGTATACC | |||||
| M154 | F: AAGAACTTCATTCACCGTCC | (TGG)102 | 417~445 | 55 | HF680309 |
| R: GTACTGCCTTCAAGGAAGTC | |||||
| M155 | F: TCTAACCCTTCCAAAATTGG | (AG)12 | 130~147 | 55 | HF680310 |
| R: AAATTATGGGCCTGTTACTG | |||||
| M156 | F: AAGCTAGCAAATACACATAGG | (CA)14(CT)9N(AAT/C)16 | 120~152 | 55 | HF680311 |
| R: CTGACAAGTTCCAGACAGGG | |||||
| Species | Locality | Sample size | Longitude | Latitude | Voucher Specimens Number |
|---|---|---|---|---|---|
| T. radicans ssp. hispidum | Yilan Co., Taiwan | 7 | N 24°30′26.2″ | E121°31′00.7″ | Hsu18286 |
| Hsinchu, Taiwan | 7 | N 24°30′17.0″ | E121°07′05.6″ | Hsu18285 | |
| Nantou, Taiwan | 6 | N 24°06′39.8″ | E121°12′50.5″ | Hsu18287 | |
| ssp. hispidum | Daguan, Yunnan, China | 7 | N 28°12′28.6″ | E103°56′26.8″ | Hsu18290 |
| Leibo, Sichuan, China | 6 | N 28°20′50.4″ | E103°43′49.8″ | Hsu18289 | |
| Pingshan, Sichuan, China | 7 | N 28°43′31.1″ | E103°58′09.7″ | Hsu18295 | |
| ssp. orientale | Kochi, Shikoku, Japan | 7 | N 33°46′02.4″ | E134°02′11.0″ | Hsu18281 |
| Okayama, Honshu, Japan | 7 | N 35°05′18.1″ | E133°31′35.6″ | Hsu18282 | |
| Nagano, Honshu, Japan | 6 | N 36°10′59.6″ | E137°31′30.0″ | Hsu18284 | |
| ssp. radicans | Washington Co., MO, USA | 7 | N 38°04′20.1″ | W90°41′57.6″ | Hsu18300 |
| Montgomery Co., MO, USA | 7 | N 38°51′25.9″ | W91°30′57.6″ | Hsu18296 | |
| Monroe Co., MO, USA | 6 | N 39°30′50.3″ | W91°47′24.1″ | Hsu18298 | |
| T. diversilobum | Butte Co., CA, USA | 7 | N 39°32′08.5″ | W121°25′24.4″ | Hsu18302 |
| Chico, CA, USA | 7 | N 39°44′06.9″ | W121°49′38.1″ | Hsu18303 | |
| Medford, OR, USA | 6 | N 42°17′34.9″ | W122°49′55.3″ | Hsu18305 | |
| T. pubescens | Carter Co., MO, USA | 7 | N 36°55′40.5″ | W91°07′12.5″ | Hsu18304 |
| Oregon Co., MO, USA | 7 | N 36°48′29.8″ | W91°07′45.3″ | Hsu18306 | |
| Howell Co., MO, USA | 6 | N 36°32′23.9″ | W91°50′29.7″ | Hsu18307 | |
| Locus | ssp. hispidum (Taiwan) | ssp. hispidum (China) | ssp. orientale | ssp. radicans | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Na | Ne | HO | HE | H | Na | Ne | HO | HE | H | Na | Ne | HO | HE | H | Na | Ne | HO | HE | H | |
| AC3 | 6 | 3.77 | 0.65 | 0.74 | 1.45 | 10 | 6.78 | 0.70 | 0.85 | 2.04 | 8 | 4.94 | 0.80 | 0.80 | 1.76 | 8 | 5.23 | 0.60 | 0.81 | 1.78 |
| AC6 | 4 | 2.62 | 0.60 | 0.62 | 1.14 | 9 | 5.44 | 0.80 | 0.82 | 1.88 | 7 | 4.60 | 0.80 | 0.78 | 1.68 | 12 | 7.55 | 0.85 | 0.87 | 2.19 |
| AC11 | 3 | 2.23 | 0.35 | 0.55 | 0.89 | 6 | 3.76 | 0.50 | 0.73 | 1.49 | 7 | 4.08 | 0.55 | 0.76 | 1.58 | 6 | 3.46 | 0.45 | 0.71 * | 1.39 |
| AC19 | 3 | 2.04 | 0.30 | 0.51 | 0.78 | 5 | 3.49 | 0.45 | 0.71 | 1.37 | 6 | 4.62 | 0.60 | 0.78 | 1.62 | 7 | 4.44 | 0.85 | 0.78 | 1.64 |
| AC20 | 3 | 2.07 | 0.25 | 0.52 | 0.78 | 3 | 2.35 | 0.50 | 0.57 | 0.94 | 3 | 2.35 | 0.50 | 0.57 | 0.94 | 4 | 2.69 | 0.60 | 0.63 | 1.11 |
| AC139 | 2 | 1.66 | 0.35 | 0.40 | 0.59 | 3 | 1.97 | 0.45 | 0.49 | 0.85 | 3 | 2.57 | 0.65 | 0.61 | 1.02 | 3 | 2.56 | 0.55 | 0.61 | 1.02 |
| AG28 | 4 | 3.24 | 0.50 | 0.69 * | 1.23 | 3 | 2.17 | 0.40 | 0.54 | 0.90 | 3 | 2.27 | 0.45 | 0.56 * | 0.90 | 3 | 2.51 | 0.45 | 0.60 | 0.98 |
| AG153 | 3 | 1.87 | 0.30 | 0.47 * | 0.82 | 2 | 1.72 | 0.30 | 0.42 | 0.61 | - | - | - | - | - | - | - | - | - | - |
| M8 | 2 | 1.47 | 0.20 | 0.32 | 0.50 | 2 | 1.22 | 0.10 | 0.18 * | 0.33 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M18 | 4 | 3.52 | 0.85 | 0.72 | 1.32 | 3 | 2.33 | 0.60 | 0.57 | 0.96 | 1 | 1.00 | - | - | - | 3 | 1.68 | 0.40 | 0.41 | 0.74 |
| M19 | 2 | 1.72 | 0.30 | 0.42 | 0.61 | 1 | 1.00 | - | - | 0.00 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M22 | 2 | 1.66 | 0.25 | 0.40 | 0.59 | 3 | 1.68 | 0.25 | 0.41 | 0.74 | 2 | 1.88 | 0.35 | 0.47 | 0.66 | 2 | 1.96 | 0.35 | 0.49 | 0.68 |
| M24 | 2 | 1.78 | 0.25 | 0.44 | 0.63 | 3 | 2.85 | 0.50 | 0.65 | 1.07 | 4 | 3.77 | 0.70 | 0.74 | 1.35 | 5 | 4.65 | 0.60 | 0.79 | 1.57 |
| M27 | 2 | 1.78 | 0.35 | 0.44 | 0.63 | 2 | 1.28 | 0.15 | 0.22 | 0.38 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M30 | 4 | 3.85 | 0.90 | 0.74 | 1.37 | 6 | 4.02 | 0.85 | 0.72 | 1.52 | 4 | 3.83 | 0.80 | 0.74 | 1.36 | 8 | 4.82 | 0.75 | 0.79 | 1.77 |
| M31 | 5 | 3.90 | 0.55 | 0.74 * | 1.45 | 8 | 5.63 | 0.65 | 0.82 | 1.88 | 1 | 1.00 | - | - | - | 4 | 1.80 | 0.35 | 0.44 | 0.86 |
| M452 | 3 | 2.30 | 0.40 | 0.57 | 0.93 | 3 | 1.11 | 0.10 | 0.10 | 0.23 | 4 | 1.78 | 0.35 | 0.44 | 0.82 | 5 | 1.68 | 0.60 | 0.41 | 0.80 |
| M493 | 2 | 1.72 | 0.30 | 0.42 | 0.61 | 2 | 1.60 | 0.30 | 0.38 | 0.56 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M54 | 6 | 4.79 | 0.75 | 0.79 | 1.67 | 6 | 3.88 | 0.70 | 0.74 | 1.56 | 6 | 3.56 | 0.70 | 0.72 | 1.45 | 6 | 3.54 | 0.50 | 0.72 * | 1.47 |
| M56 | 1 | 1.00 | - | - | 0.00 | 5 | 2.42 | 0.45 | 0.59 | 1.16 | 5 | 3.62 | 0.50 | 0.72 | 1.42 | 6 | 3.16 | 0.55 | 0.68 | 1.41 |
| M60 | 6 | 4.65 | 1.00 | 0.79 | 1.65 | 6 | 3.98 | 0.75 | 0.75 | 1.59 | 2 | 1.83 | 0.30 | 0.46 | 0.65 | 5 | 2.17 | 0.50 | 0.54 | 1.09 |
| M61 | 2 | 1.92 | 0.40 | 0.48 | 0.67 | 6 | 4.12 | 0.75 | 0.76 | 1.57 | 7 | 3.90 | 0.85 | 0.74 | 1.60 | 8 | 5.52 | 0.80 | 0.82 | 1.86 |
| M64 | 4 | 3.38 | 0.55 | 0.70 | 1.27 | 3 | 2.60 | 0.60 | 0.62 | 1.01 | 3 | 2.06 | 0.40 | 0.52 | 0.82 | 3 | 1.80 | 0.45 | 0.45 | 0.75 |
| M66 | 2 | 1.88 | 0.45 | 0.47 | 0.66 | 3 | 2.46 | 0.55 | 0.59 | 0.97 | 1 | 1.00 | - | - | - | 2 | 1.78 | 0.35 | 0.44 | 0.63 |
| M67 | 1 | 1.00 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| M68 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M821 | 2 | 1.78 | 0.45 | 0.44 | 0.63 | 2 | 1.47 | 0.30 | 0.32 | 0.50 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M822 | 1 | 1.00 | - | - | 0.00 | 2 | 1.16 | 0.15 | 0.14 | 0.27 | 2 | 1.91 | 0.35 | 0.48 | 0.81 | 4 | 1.95 | 0.40 | 0.49 | 0.89 |
| M83 | 2 | 1.98 | 0.50 | 0.50 | 0.69 | 2 | 1.41 | 0.25 | 0.29 | 0.46 | 2 | 1.28 | 0.15 | 0.22 | 0.38 | 2 | 1.28 | 0.15 | 0.22 | 0.38 |
| M85 | 3 | 2.97 | 0.40 | 0.66 | 1.09 | 3 | 2.52 | 0.50 | 0.60 | 1.00 | - | - | - | - | - | - | - | - | - | - |
| M97 | 5 | 3.52 | 0.75 | 0.72 | 1.37 | 5 | 3.76 | 0.60 | 0.73 | 1.45 | 1 | 1.00 | - | - | - | 4 | 2.03 | 0.35 | 0.51 | 0.98 |
| M99 | 6 | 4.08 | 0.50 | 0.76 | 1.51 | 4 | 3.29 | 0.60 | 0.70 | 1.28 | 2 | 1.47 | 0.20 | 0.32 | 0.50 | 2 | 1.96 | 0.35 | 0.49 | 0.68 |
| M104 | 5 | 3.86 | 0.60 | 0.74 | 1.46 | 3 | 2.63 | 0.40 | 0.62 * | 1.03 | 1 | 1.00 | - | - | - | 2 | 1.72 | 0.30 | 0.42 | 0.61 |
| M120 | 7 | 4.82 | 0.70 | 0.79 | 1.73 | 6 | 5.06 | 0.70 | 0.80 | 1.71 | 4 | 2.79 | 0.60 | 0.64 | 1.15 | 5 | 2.56 | 0.55 | 0.61 | 1.19 |
| M121 | 3 | 2.47 | 0.50 | 0.60 | 1.00 | 3 | 2.69 | 0.55 | 0.63 | 1.04 | 2 | 1.72 | 0.30 | 0.42 | 0.61 | 3 | 2.38 | 0.70 | 0.58 | 0.94 |
| M123 | 3 | 2.69 | 0.55 | 0.63 | 1.04 | 6 | 3.24 | 0.65 | 0.69 | 1.46 | 7 | 4.76 | 0.60 | 0.79 | 1.70 | 7 | 3.96 | 0.80 | 0.75 | 1.63 |
| M124 | 6 | 3.13 | 0.65 | 0.68 | 1.35 | 4 | 3.01 | 0.65 | 0.67 | 1.17 | 3 | 2.35 | 0.40 | 0.57 | 0.94 | 4 | 2.78 | 0.60 | 0.64 | 1.17 |
| M137 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M148 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M154 | 3 | 2.97 | 0.60 | 0.66 | 1.09 | 4 | 2.91 | 0.60 | 0.66 | 1.18 | 5 | 3.03 | 0.45 | 0.67 | 1.27 | 6 | 3.94 | 0.65 | 0.75 | 1.48 |
| M155 | 3 | 2.75 | 0.40 | 0.64 | 1.06 | 4 | 2.68 | 0.45 | 0.63 | 1.15 | 4 | 2.91 | 0.50 | 0.66 | 1.19 | 4 | 2.67 | 0.50 | 0.63 | 1.17 |
| M156 | 7 | 1.83 | 0.40 | 0.45 | 1.01 | 7 | 2.95 | 0.65 | 0.66 * | 1.39 | 6 | 1.47 | 0.65 | 0.74 | 1.47 | 7 | 3.15 | 0.65 | 0.68 | 1.42 |
| Mean | 3.26 | 2.49 | 0.49 | 0.59 | 0.98 | 3.93 | 2.73 | 0.50 | 0.59 | 1.07 | 3.18 | 2.27 | 0.52 | 0.61 | 0.78 | 4.05 | 2.60 | 0.53 | 0.60 | 0.96 |
| Locus | T. diversilobum | T. pubescens | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Na | Ne | HO | HE | H | Na | Ne | HO | HE | H | |
| AC3 | 6 | 3.94 | 0.70 | 0.75 | 1.54 | 6 | 3.94 | 0.65 | 0.75 | 1.54 |
| AC6 | 5 | 4.62 | 0.80 | 0.78 | 1.56 | 5 | 4.71 | 0.80 | 0.79 | 1.58 |
| AC11 | 8 | 5.80 | 0.85 | 0.83 | 1.90 | 8 | 5.56 | 0.90 | 0.82 | 1.87 |
| AC19 | 5 | 3.69 | 0.80 | 0.73 | 1.45 | 6 | 3.08 | 0.65 | 0.68 | 1.43 |
| AC20 | 2 | 1.96 | 0.35 | 0.49 | 0.68 | 2 | 1.96 | 0.45 | 0.49 | 0.68 |
| AC139 | - | - | - | - | - | - | - | - | - | - |
| AG28 | 4 | 3.29 | 0.50 | 0.70 | 1.28 | 3 | 2.82 | 0.40 | 0.65 | 1.07 |
| AG153 | - | - | - | - | - | - | - | - | - | - |
| M8 | - | - | - | - | - | - | - | - | - | - |
| M18 | 5 | 2.83 | 0.60 | 0.65 | 1.25 | 4 | 3.92 | 0.50 | 0.75 * | 1.38 |
| M19 | 3 | 1.94 | 0.35 | 0.48 | 0.83 | 1 | 1.00 | - | - | - |
| M22 | 3 | 1.94 | 0.35 | 0.48 | 0.83 | 3 | 2.52 | 0.45 | 0.60 | 1.00 |
| M24 | - | - | - | - | - | - | - | - | - | - |
| M27 | 2 | 2.00 | 0.40 | 0.50 | 0.69 | 2 | 1.98 | 0.40 | 0.50 | 0.69 |
| M30 | 6 | 2.74 | 0.60 | 0.64 | 1.26 | 3 | 2.20 | 0.55 | 0.55 | 0.86 |
| M31 | - | - | - | - | - | - | - | - | - | - |
| M452 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M493 | - | - | - | - | - | - | - | - | - | - |
| M54 | - | - | - | - | - | - | - | - | - | - |
| M56 | - | - | - | - | - | - | - | - | - | - |
| M60 | 7 | 5.30 | 0.80 | 0.81 | 1.77 | 4 | 2.71 | 0.65 | 0.63 * | 1.15 |
| M61 | 5 | 3.40 | 0.60 | 0.71 | 1.39 | 2 | 1.34 | 0.30 | 0.26 | 0.42 |
| M64 | - | - | - | - | - | - | - | - | - | - |
| M66 | 2 | 1.60 | 0.30 | 0.38 | 0.56 | 2 | 1.98 | 0.30 | 0.50 | 0.69 |
| M67 | - | - | - | - | - | - | - | - | - | - |
| M68 | - | - | - | - | - | - | - | - | - | - |
| M821 | 2 | 1.98 | 0.40 | 0.50 | 0.69 | 2 | 1.98 | 0.40 | 0.50 | 0.69 |
| M822 | 2 | 1.60 | 0.50 | 0.38 | 0.56 | 2 | 1.60 | 0.40 | 0.38 | 0.56 |
| M83 | 2 | 1.96 | 0.45 | 0.49 | 0.68 | 2 | 1.98 | 0.50 | 0.50 | 0.69 |
| M85 | 1 | 1.00 | - | - | - | 1 | 1.00 | - | - | - |
| M97 | - | - | - | - | - | - | - | - | - | - |
| M99 | - | - | - | - | - | - | - | - | - | - |
| M104 | 6 | 4.85 | 0.80 | 0.79 | 1.67 | 5 | 4.19 | 0.60 | 0.76 | 1.49 |
| M120 | 3 | 2.06 | 0.45 | 0.52 | 0.89 | 2 | 1.83 | 0.30 | 0.46 | 0.65 |
| M121 | - | - | - | - | - | - | - | - | - | - |
| M123 | 5 | 2.29 | 0.55 | 0.56 | 1.13 | 5 | 2.32 | 0.55 | 0.57 | 1.15 |
| M124 | 5 | 4.28 | 0.65 | 0.77 | 1.53 | 5 | 4.19 | 0.70 | 0.76 | 1.52 |
| M137 | - | - | - | - | - | - | - | - | - | - |
| M148 | - | - | - | - | - | - | - | - | - | - |
| M154 | 3 | 2.38 | 0.45 | 0.58 | 0.94 | 3 | 2.22 | 0.40 | 0.55 | 0.92 |
| M155 | 3 | 2.69 | 0.60 | 0.63 | 1.04 | 3 | 2.52 | 0.60 | 0.60 | 1.00 |
| M156 | - | - | - | - | - | - | - | - | - | - |
| Mean | 3.84 | 2.85 | 0.56 | 0.62 | 0.69 | 3.28 | 2.58 | 0.52 | 0.58 | 0.61 |
Acknowledgments
Conflicts of Interest
References
- Gladman, A.C. Toxicodendron dermatitis: Poison ivy, oak, and sumac. Wilderness Environ. Med 2006, 17, 120–128. [Google Scholar]
- Billets, S.; Craig, J.C.; Corbett, M.D.; Vickery, J.F. Component analysis of the urushiol content of poison ivy and poison oak. Phytochemistry 1976, 15, 533–535. [Google Scholar]
- Dawson, C.R. The chemistry of poison ivy. Trans. N. Y. Acad. Sci 1956, 18, 427–443. [Google Scholar]
- Gillis, W.T. The systematics and ecology of poison-ivy and the poison-oaks (Toxicodendron, Anacardiaceae). Rhodora 1971, 73, 370–443. [Google Scholar]
- Gillis, W.T. Poison-ivy and its kin. Arnoldia 1975, 35, 93–123. [Google Scholar]
- Liao, P.C.; Chao, Y.S.; Chiang, Y.C. Chapter 2-Species Occurrence, Hybridization and Speciation in Postglacial East and South Asia. In The Molecular Basis of Plant Genetic Diversity; Caliskan, M., Ed.; InTech–Open Access Publisher: Rijeka, Croatia, 2012; pp. 33–60. [Google Scholar]
- Milne, R.I. Phylogeny and biogeography of Rhododendron subsection Pontica, a group with a tertiary relict distribution. Mol. Phylogenet. Evol 2004, 33, 389–401. [Google Scholar]
- Wen, J. Evolution of eastern Asian and eastern North American biogeographic disjunctions: A few additional issues. Int. J. Plant Sci 2001, 162, S117–S122. [Google Scholar]
- Barkley, F.A. A monographic study of Rhus and its immediate allies in North and Central America, including the West Indies. Ann. MO Bot. Gard 1937, 24, 265–498. [Google Scholar]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res 1999, 27, 573–580. [Google Scholar]
- Kalendar, R.; Lee, D.; Schulman, A.H. Java web tools for PCR, in silico PCR, and oligonucleotide assembly and analysis. Genomics 2011, 98, 137–144. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar]
- Ge, X.J.; Hsu, T.W.; Hung, K.H.; Lin, C.J.; Huang, C.C.; Huang, C.C.; Chiang, Y.C.; Chiang, T.Y. Inferring mutiple refugia and phylogeographical patterns in Pinus massoniana based on nucleotide sequence variation and fingerprinting. PLoS One 2012, 7, e43717. [Google Scholar]
- Chiang, Y.C.; Huang, B.H.; Chang, C.W.; Wan, Y.T.; Lai, S.J.; Huang, S.; Liao, P.C. Asymmetric introgression in the horticultural living fossil Cycas Sect. Asiorientales using a genome-wide scanning approach. Int. J. Mol. Sci 2013, 14, 8228–8251. [Google Scholar]
- Liao, P.C.; Tsai, C.C.; Chou, C.H.; Chiang, Y.C. Introgression between cultivars and wild populations of Momordica charantia L. (Cucurbitaceae) in Taiwan. Int. J. Mol. Sci 2012, 13, 6469–6491. [Google Scholar]
- Zane, L.; Bargelloni, L.; Patarnello, T. Strategies for microsatellite isolation: A review. Mol. Ecol 2002, 11, 1–6. [Google Scholar]
- Liao, P.C.; Gong, X.; Shih, H.C.; Chiang, Y.C. Isolation and characterization of eleven polymorphic microsatellite loci from an endemic species, Piper polysyphonum (Piperaceae). Conserv. Genet 2009, 10, 1911–1914. [Google Scholar]
- Chiang, Y.C.; Shih, H.C.; Chang, L.W.; Li, W.R.; Lin, H.Y.; Ju, L.P. Isolation of 16 polymorphic microsatellite markers from an endangered and endemic species, Podocarpus nakaii (Podocarpaceae). Am. J. Bot 2011, 98, e306–e309. [Google Scholar]
- Chiang, Y.C.; Shih, H.C.; Huang, M.C.; Ju, L.P.; Hung, K.H. The characterization of microsatellite loci from an endemic tree Litsea hypophaea (Lauraceae) in Taiwan. Am. J. Bot 2012, 99, e251–e254. [Google Scholar]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour 2010, 10, 564–567. [Google Scholar]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour 2009, 9, 1322–1332. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol 2005, 14, 2611–2620. [Google Scholar]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resourc 2012, 4, 359–361. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hsu, T.-W.; Shih, H.-C.; Kuo, C.-C.; Chiang, T.-Y.; Chiang, Y.-C. Characterization of 42 Microsatellite Markers from Poison Ivy, Toxicodendron radicans (Anacardiaceae). Int. J. Mol. Sci. 2013, 14, 20414-20426. https://doi.org/10.3390/ijms141020414
Hsu T-W, Shih H-C, Kuo C-C, Chiang T-Y, Chiang Y-C. Characterization of 42 Microsatellite Markers from Poison Ivy, Toxicodendron radicans (Anacardiaceae). International Journal of Molecular Sciences. 2013; 14(10):20414-20426. https://doi.org/10.3390/ijms141020414
Chicago/Turabian StyleHsu, Tsai-Wen, Huei-Chuan Shih, Chia-Chi Kuo, Tzen-Yuh Chiang, and Yu-Chung Chiang. 2013. "Characterization of 42 Microsatellite Markers from Poison Ivy, Toxicodendron radicans (Anacardiaceae)" International Journal of Molecular Sciences 14, no. 10: 20414-20426. https://doi.org/10.3390/ijms141020414