Glycosyltransferase GLT8D2 Positively Regulates ApoB100 Protein Expression in Hepatocytes

Abstract
:1. Introduction
2. Results
2.1. Amplification of GLT8D2 Gene
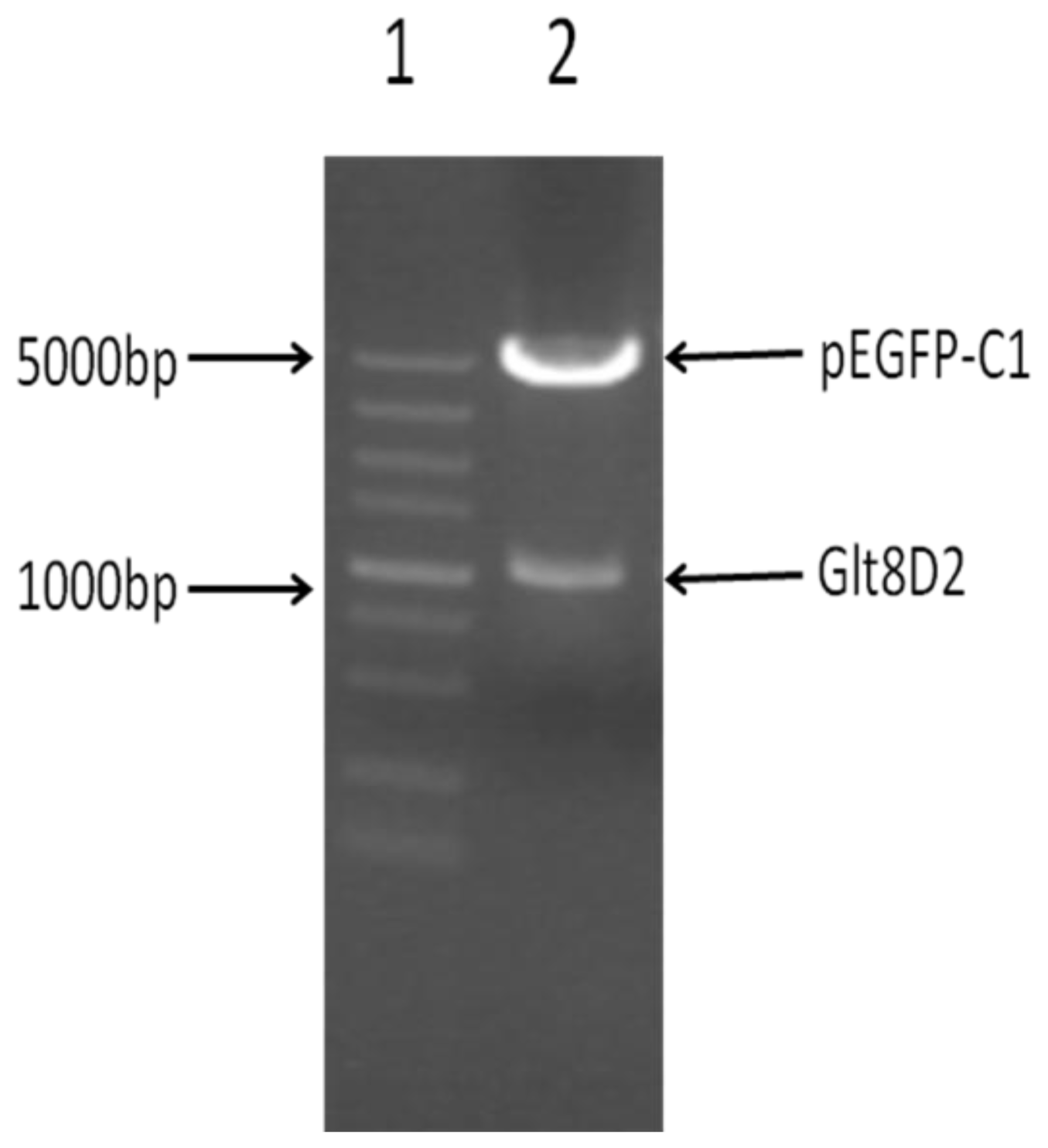
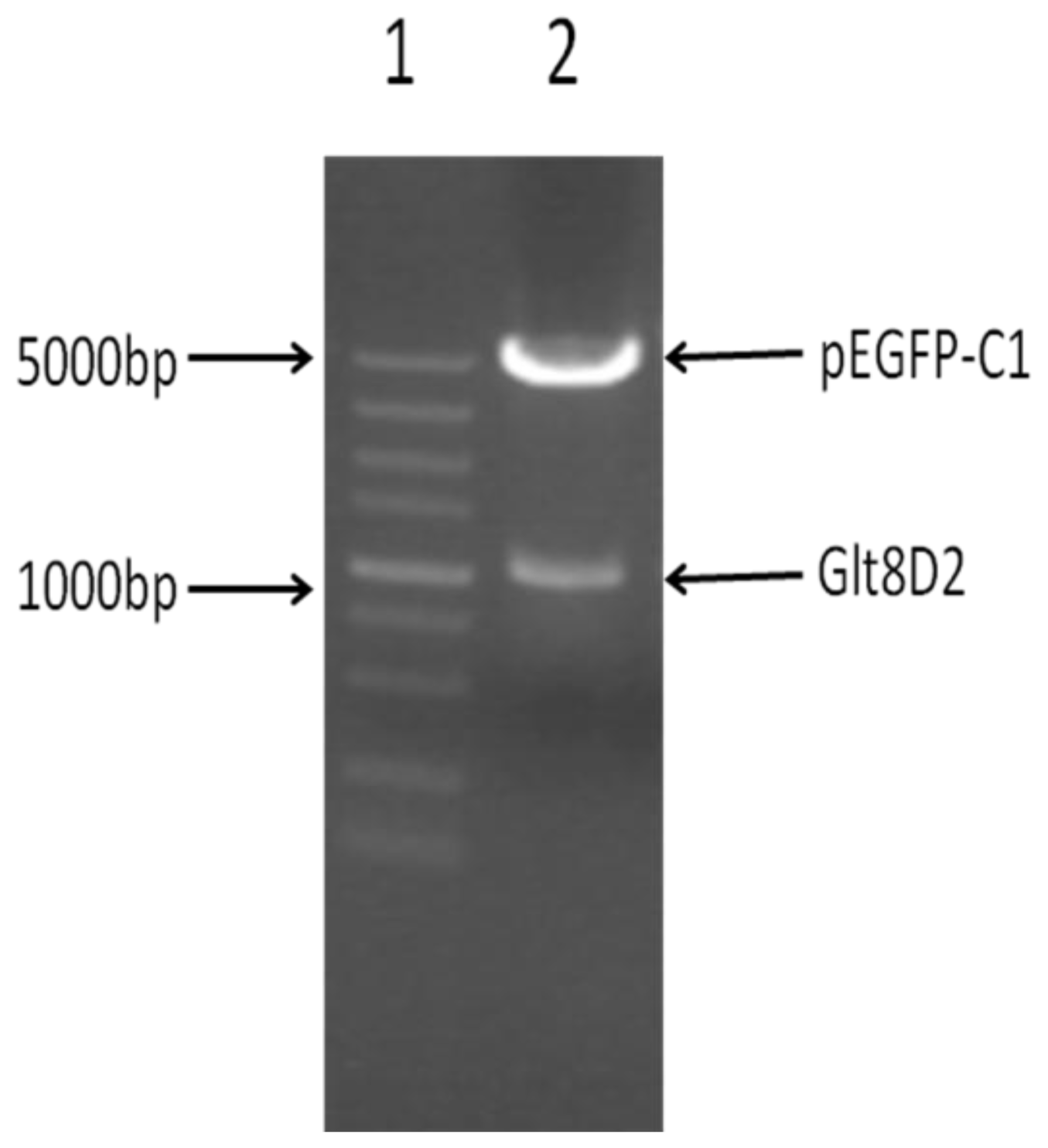
2.2. Successful Construction of Plasmids
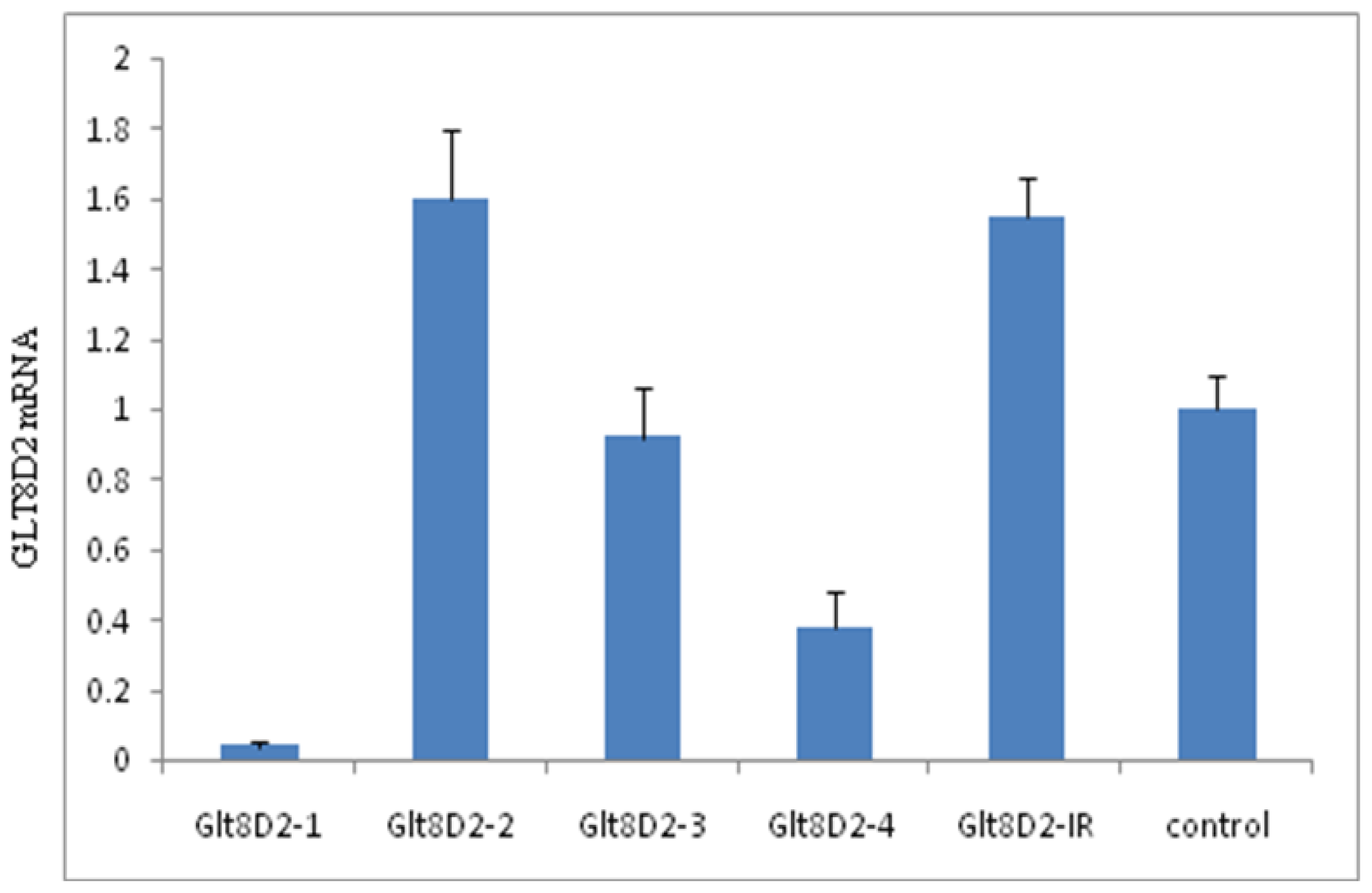
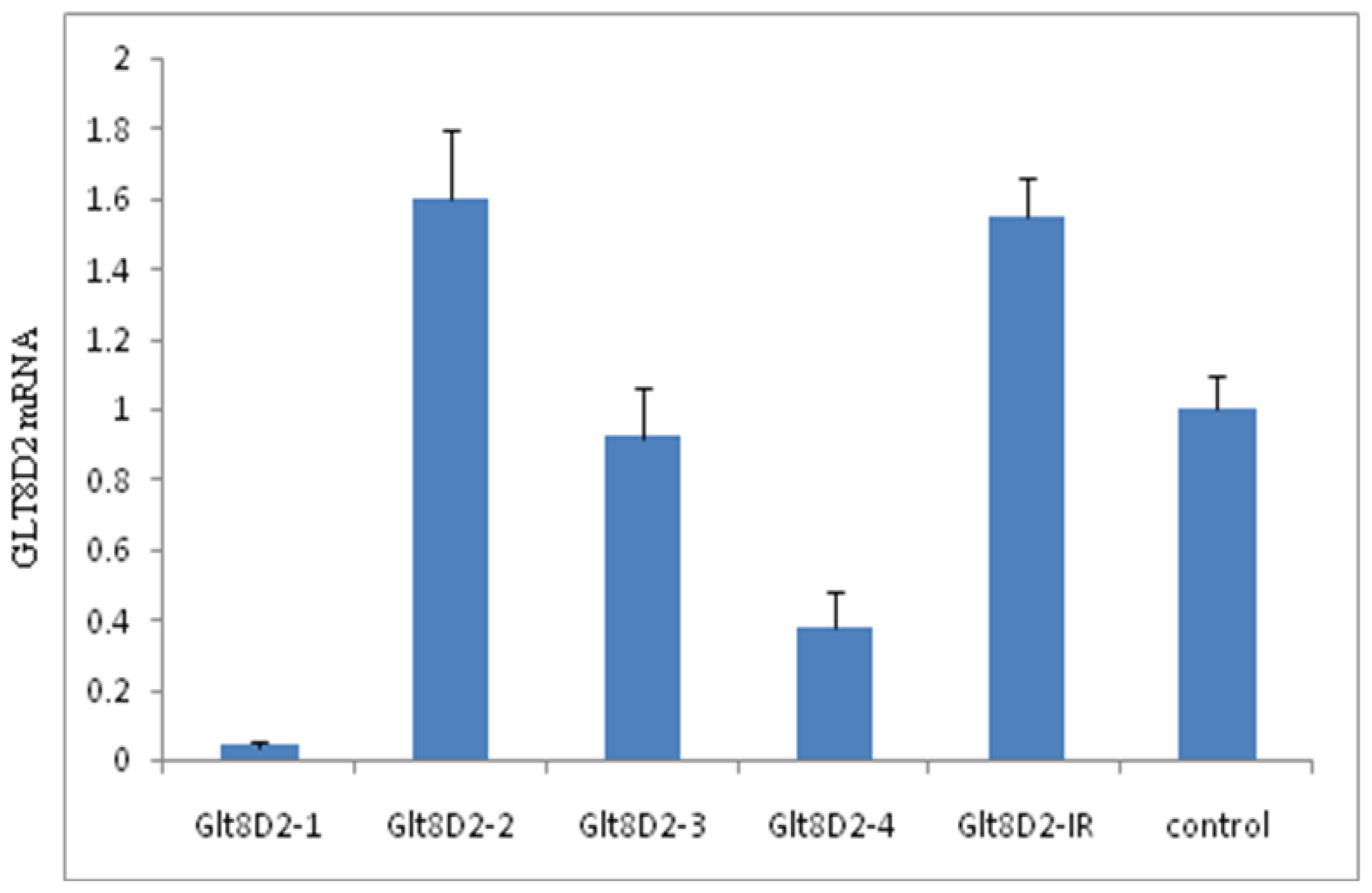
2.3. Knockdown Efficiency of pcDNA6.2-GLT8D2 shRNAs in HepG2 Cells
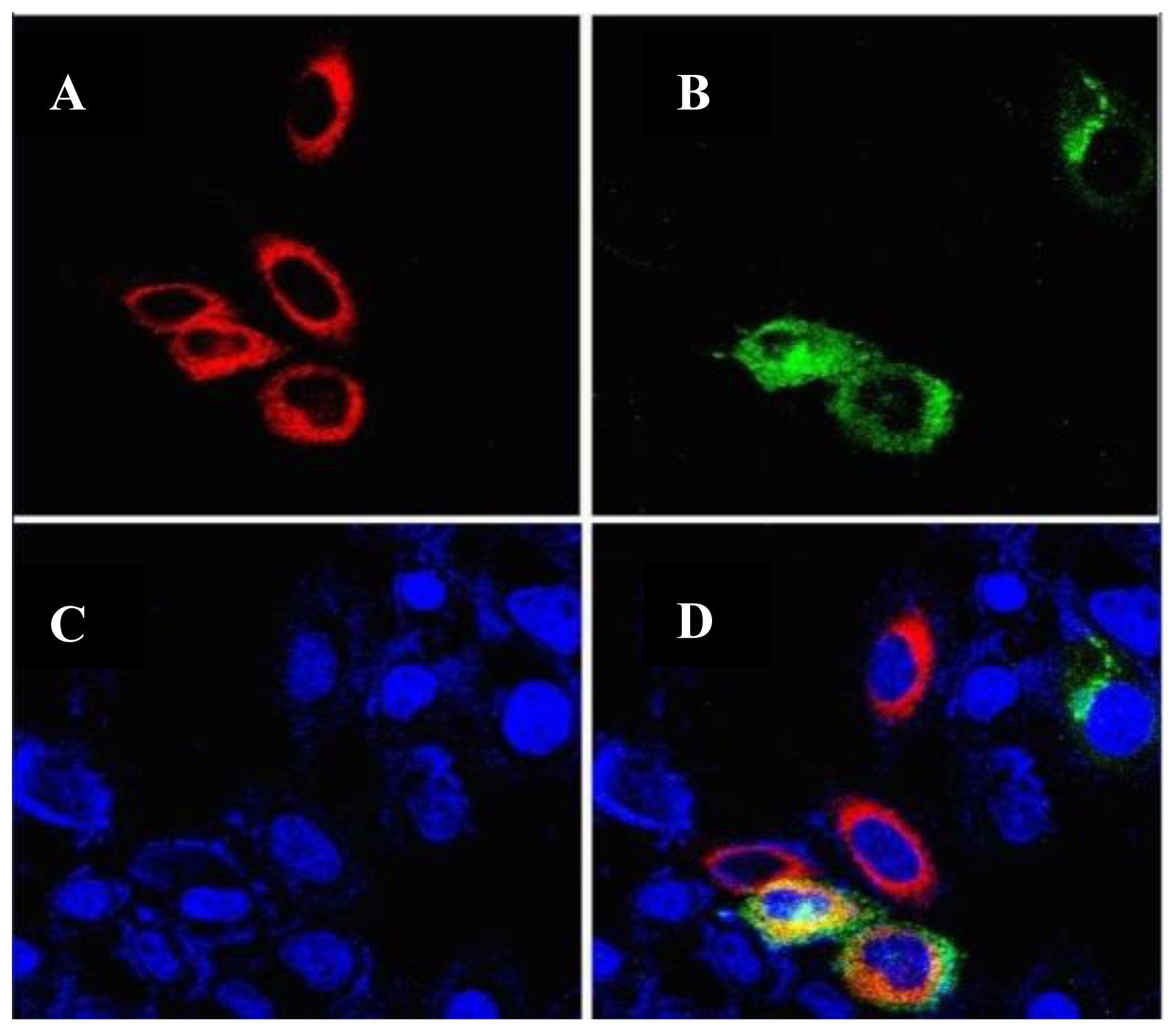
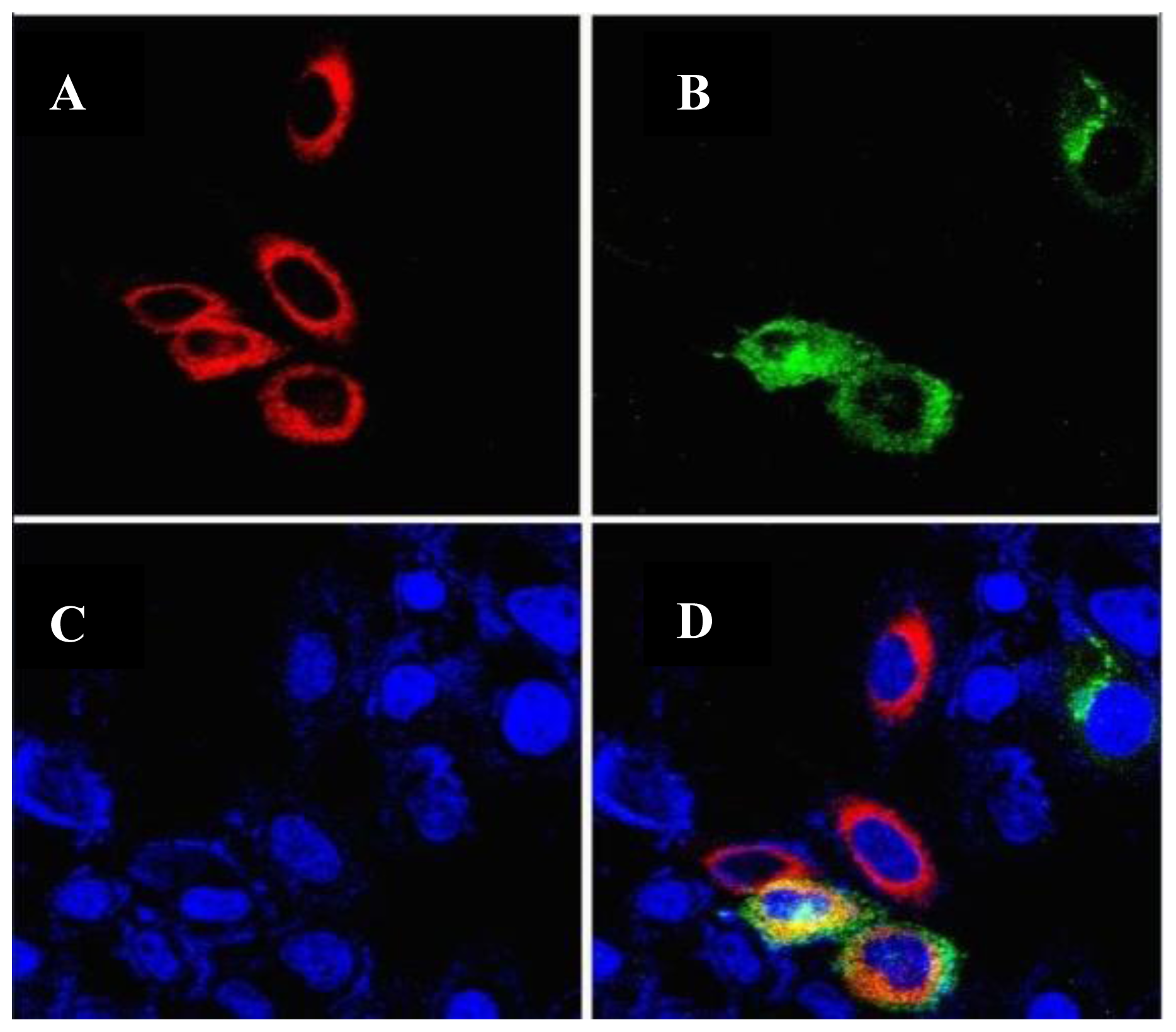
2.4. Intracellular Localization of the GLT8D2 Protein
2.5. Direct Interaction of GLT8D2 and Apo-B100
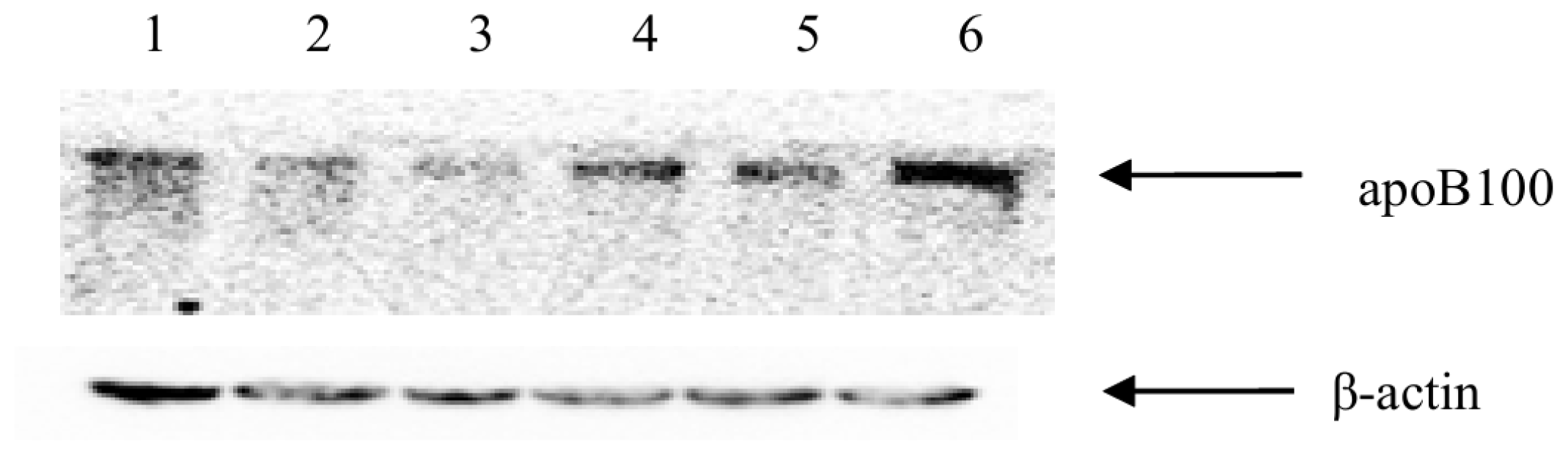
2.6. GLT8D2 Affects ApoB100 Expression in HepG2 Cells
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Cell Culture
4.3. Total RNA Extraction and cDNA Amplification
4.4. Plasmid Construction
4.4.1. Plasmid pEGFP-C1-GLT8D2
4.4.2. Plasmid pcDNA6.2-GLT8D2 shRNA
4.4.3. Plasmid pEGFP-C1-muta-GLT8D2
4.4.4. Other Plasmids
4.5. Intracellular Localization of GLT8D2
4.6. Co-Immunoprecipitation (Co-IP) and Western Blot (WB)
4.7. Detection of ApoB100 Expression in HepG2 Cells
4.8. Statistics
5. Conclusions




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| shRNA ID | Sequences | |
|---|---|---|
| shRNA GLT8D2 1 | Sence: | 5′-TGCTGTCATGTTGGCAACAATCACACGTTTTGGCCACTGACTGACGTGTGATTTGCCAACATGA-3′ |
| Antisence: | 5′-CCTGTCATGTTGGCAAATCACACGTCAGTCAGTGGCCAAAACGTGTGATTGTTGCCAACATGAC-3′ | |
| shRNA GLT8D2 2 | Sence: | 5′-TGCTGTTCCCATGAAACACAATCAGCGTTTTGGCCACTGACTGACGCTGATTGTTTCATGGGAA-3′ |
| Antisence: | 5′-CCTGTTCCCATGAAACAATCAGCGTCAGTCAGTGGCCAAAACGCTGATTGTGTTTCATGGGAAC-3′ | |
| shRNA GLT8D2 3 | Sence: | 5′-TGCTGAAATGCTCCGAATATCTGGCAGTTTTGGCCACTGACTGACTGCCAGATTCGGAGCATTT-3′ |
| Antisence: | 5′-CCTGAAATGCTCCGAATCTGGCAGTCAGTCAGTGGCCAAAACTGCCAGATATTCGGAGCATTTC-3′ | |
| shRNA GLT8D2 4 | Sence: | 5′-TGCTGTACAGAATCACACAGAGGGTCGTTTTGGCCACTGACTGACGACCCTCTGTGATTCTGTA-3′ |
| Antisence: | 5′-CCTGTACAGAATCACAGAGGGTCGTCAGTCAGTGGCCAAAACGACCCTCTGTGTGATTCTGTAC-3′ | |
| shRNA GLT8D2 IR | Sence: | 5′-TGCTGAAATGTACTGCGCGTGGAGACGTTTTGGCCACTGACTGACGTCTCCACGCAGTACATTT-3′ |
| Antisence: | 5′-CCTGAAATGTACTGCGTGGAGACGTCAGTCAGTGGCCAAAACGTCTCCACGCGCAGTACATTTC-3′ | |
Acknowledgments
Conflicts of Interest
References
- Zhan, Y.T.; Weng, J.; Li, L.; Xu, Q.; Song, X.; Guo, X.X. Protective effect of probucol on liver injury induced by carbon tetrachloride in rats. Hepatol. Int 2011, 5, 899–905. [Google Scholar]
- Zhan, Y.T.; An, W. Roles of liver innate immune cells in nonalcoholic fatty liver disease. World J. Gastroenterol 2010, 16, 4652–4660. [Google Scholar]
- Gabbi, C.; Bertolotti, M.; Anzivino, C.; Macchioni, D.; Del Puppo, M.; Ricchi, M.; Carubbi, F.; Tagliafico, E.; Romagnoli, D.; Odoardi, M.R.; et al. Effects of bile duct ligation and cholic acid treatment on fatty liver in two rat models of non-alcoholic fatty liver disease. Dig. Liver Dis 2012, 44, 1018–1026. [Google Scholar]
- Quercioli, A.; Montecucco, F.; Mach, F. Update on the treatments of non-alcoholic fatty liver disease (NAFLD). Cardiovasc. Hematol. Disord. Drug Targets 2009, 9, 261–270. [Google Scholar]
- Fisher, E.A. The degradation of apolipoprotein B100: Multiple opportunities to regulate VLDL triglyceride production by different proteolytic pathways. Biochim. Biophys. Acta 2012, 1821, 778–781. [Google Scholar]
- Fabbrini, E.; Mohammed, B.S.; Magkos, F.; Korenblat, K.M.; Patterson, B.W.; Klein, S. Alterations in adipose tissue and hepatic lipid kinetics in obese men and women with nonalcoholic fatty liver disease. Gastroenterology 2008, 134, 424–431. [Google Scholar]
- Lusis, A.J.; Pajukanta, P. A treasure trove for lipoprotein biology. Nat. Genet 2008, 40, 129–130. [Google Scholar]
- Liang, J.S.; Kim, T.; Fang, S.; Yamaguchi, J.; Weissman, A.M.; Fisher, E.A.; Ginsberg, H.N. Overexpression of the tumor autocrine motility factor receptor Gp78, a ubiquitin protein ligase, results in increased ubiquitinylation and decreased secretion of apolipoprotein B100 in HepG2 cells. J. Biol. Chem 2003, 278, 23984–23988. [Google Scholar]
- Barvkar, V.T.; Pardeshi, V.C.; Kale, S.M.; Kadoo, N.Y.; Gupta, V.S. Phylogenomic analysis of UDP glycosyltransferase 1 multigene family in Linum usitatissimum identified genes with varied expression patterns. BMC Genomics 2012, 13, 175. [Google Scholar]
- Taniguchi, N.; Nakamura, K.; Narimatsu, H.; Von der Lieth, C.W.; Paulson, J. Human disease glycomics/proteome initiative workshop and the 4th HUPO annual congress. Proteomics 2006, 6, 12–13. [Google Scholar]
- UniProt. Available online: http://www.uniprot.org/uniprot/Q9H1C3 (accessed on 17 September 2013).
- Paschen, W.; Doutheil, J. Disturbances of the functioning of endoplasmic reticulum: A key mechanism underlying neuronal cell injury? J. Cereb. Blood Flow Metab 1999, 19, 1–18. [Google Scholar]
- Csala, M.; Kereszturi, É.; Mandl, J.; Bánhegyi, G. The endoplasmic reticulum as the extracellular space inside the cell: Role in protein folding andglycosylation. Antioxid. Redox Signal 2012, 16, 1100–1108. [Google Scholar]
- Harazono, A.; Kawasaki, N.; Kawanishi, T.; Hayakawa, T. Site-specific glycosylation analysis of human apolipoprotein B100 using LC/ESI MS/MS. Glycobiology 2005, 15, 447–462. [Google Scholar]
- Ihara, Y.; Yoshimura, M.; Miyoshi, E.; Nishikawa, A.; Sultan, A.S.; Toyosawa, S.; Ohnishi, A.; Suzuki, M.; Yamamura, K.; Ijuhin, N.; et al. Ectopic expression of N-acetylglucosaminyltransferase III in transgenic hepatocytes disrupts apolipoprotein B secretion and induces aberrant cellular morphology with lipid storage. Proc. Natl. Acad. Sci. USA 1998, 95, 2526–2530. [Google Scholar]
- Cartwright, I.J.; Higgins, J.A. Intracellular events in the assembly of very- low-density-lipoprotein lipids with apolipoprotein B in isolated rabbit hepatocytes. Biochem. J 1995, 310, 897–907. [Google Scholar]
- Rutledge, A.C.; Su, Q.; Adeli, K. Apolipoprotein B100 biogenesis: A complex array of intracellular mechanisms regulating folding, stability, and lipoprotein assembly. Biochem. Cell Biol 2010, 88, 251–267. [Google Scholar]
- Liao, W.; Chan, L. Tunicamycin induces ubiquitination and degradation of apolipoprotein B in HepG2 cells. Biochem. J 2001, 353, 493–501. [Google Scholar]
- Macri, J.; Adeli, K. Conformational changes in apolipoprotein B modulate intracellular assembly and degradation of ApoB-containing lipoprotein particles in HepG2 cells. Arterioscler. Thromb. Vasc. Biol 1997, 17, 2982–2994. [Google Scholar]
- Nagae, M.; Yamaguchi, Y. Function and 3D Structure of the N-Glycans on Glycoproteins. Int. J. Mol. Sci 2012, 13, 8398–8429. [Google Scholar]
- Hirayama, H.; Seino, J.; Kitajima, T.; Jigami, Y.; Suzuki, T. Free oligosaccharides to monitor glycoprotein endoplasmic reticulum-associated degradation in Saccharomyces cerevisiae. J. Biol. Chem 2010, 285, 12390–12404. [Google Scholar]
- Kawai, F.; Grass, S.; Kim, Y.; Choi, K.J.; St Geme, J.W., 3rd; Yeo, H.J. C-terminus glycans with critical functional role in the maturation of secretory glycoproteins. PLoS One 2011, 6, e19979. [Google Scholar]
- Cioaca, D.; Ghenea, S.; Spiridon, L.N.; Marin, M.; Petrescu, A.J.; Petrescu, S.M. Structural insights into the glycosyltransferase activity of the Actinobacillus pleuropneumoniae HMW1C-like protein. J. Biol. Chem 2011, 286, 38546–38557. [Google Scholar]
- Qiu, W.; Avramoglu, R.K.; Rutledge, A.C.; Tsai, J.; Adeli, K. Mechanisms of glucosamine- induced suppression of the hepatic assembly and secretion of apolipoprotein B-100-containing lipoproteins. J. Lipid Res 2006, 47, 1749–1761. [Google Scholar]
- Vukmirica, J.; Nishimaki-Mogami, T.; Tran, K.; Shan, J.; McLeod, R.S.; Yuan, J.; Yao, Z. The N-linked oligosaccharides at the amino terminus of human apoB are important for the assembly and secretion of VLDL. J. Lipid Res 2002, 43, 1496–1507. [Google Scholar]
- Giraudo, C.G.; Maccioni, H.J. Endoplasmic reticulum export of glycosyltransferases depends on interaction of a cytoplasmic dibasic motif with Sar1. Mol. Biol. Cell 2003, 14, 3753–3766. [Google Scholar]
- Gusarova, V.; Brodsky, J.L.; Fisher, E.A. Apolipoprotein B100 exit from the endoplasmic reticulum (ER) is COPII-dependent, and its lipidation to very low density lipoprotein occurs post-ER. J. Biol. Chem 2003, 278, 48051–48058. [Google Scholar]
- Chan, L. Apolipoprotein B, the major protein component of triglyceride-rich and low density lipoproteins. J. Biol. Chem 1992, 267, 25621–25624. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wei, H.-S.; Wei, H.-L.; Zhao, F.; Zhong, L.-P.; Zhan, Y.-T. Glycosyltransferase GLT8D2 Positively Regulates ApoB100 Protein Expression in Hepatocytes. Int. J. Mol. Sci. 2013, 14, 21435-21446. https://doi.org/10.3390/ijms141121435
Wei H-S, Wei H-L, Zhao F, Zhong L-P, Zhan Y-T. Glycosyltransferase GLT8D2 Positively Regulates ApoB100 Protein Expression in Hepatocytes. International Journal of Molecular Sciences. 2013; 14(11):21435-21446. https://doi.org/10.3390/ijms141121435
Chicago/Turabian StyleWei, Hong-Shan, Hong-Lian Wei, Fei Zhao, Le-Ping Zhong, and Yu-Tao Zhan. 2013. "Glycosyltransferase GLT8D2 Positively Regulates ApoB100 Protein Expression in Hepatocytes" International Journal of Molecular Sciences 14, no. 11: 21435-21446. https://doi.org/10.3390/ijms141121435