A Novel Insight into the Cardiotoxicity of Antineoplastic Drug Doxorubicin


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
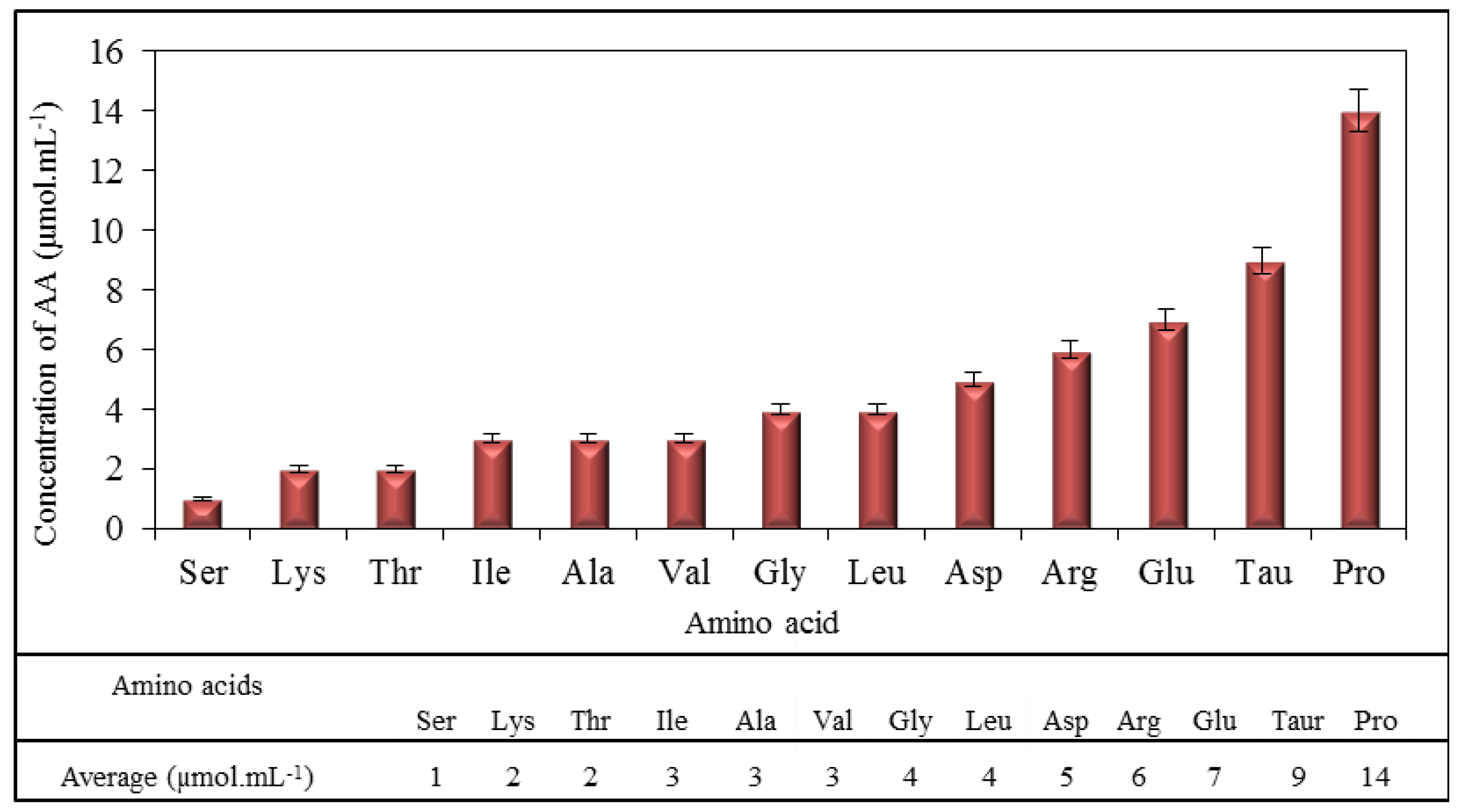
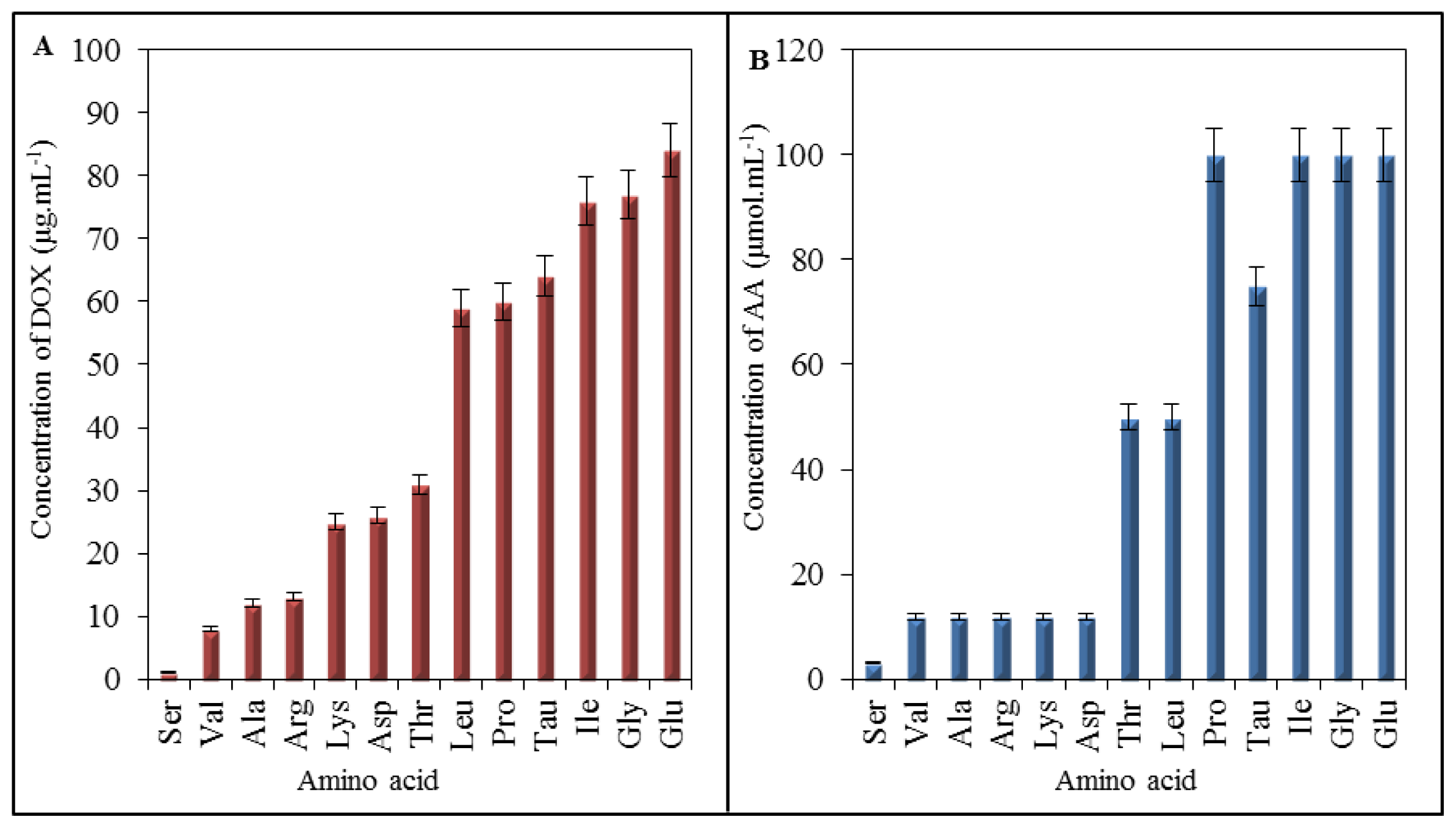
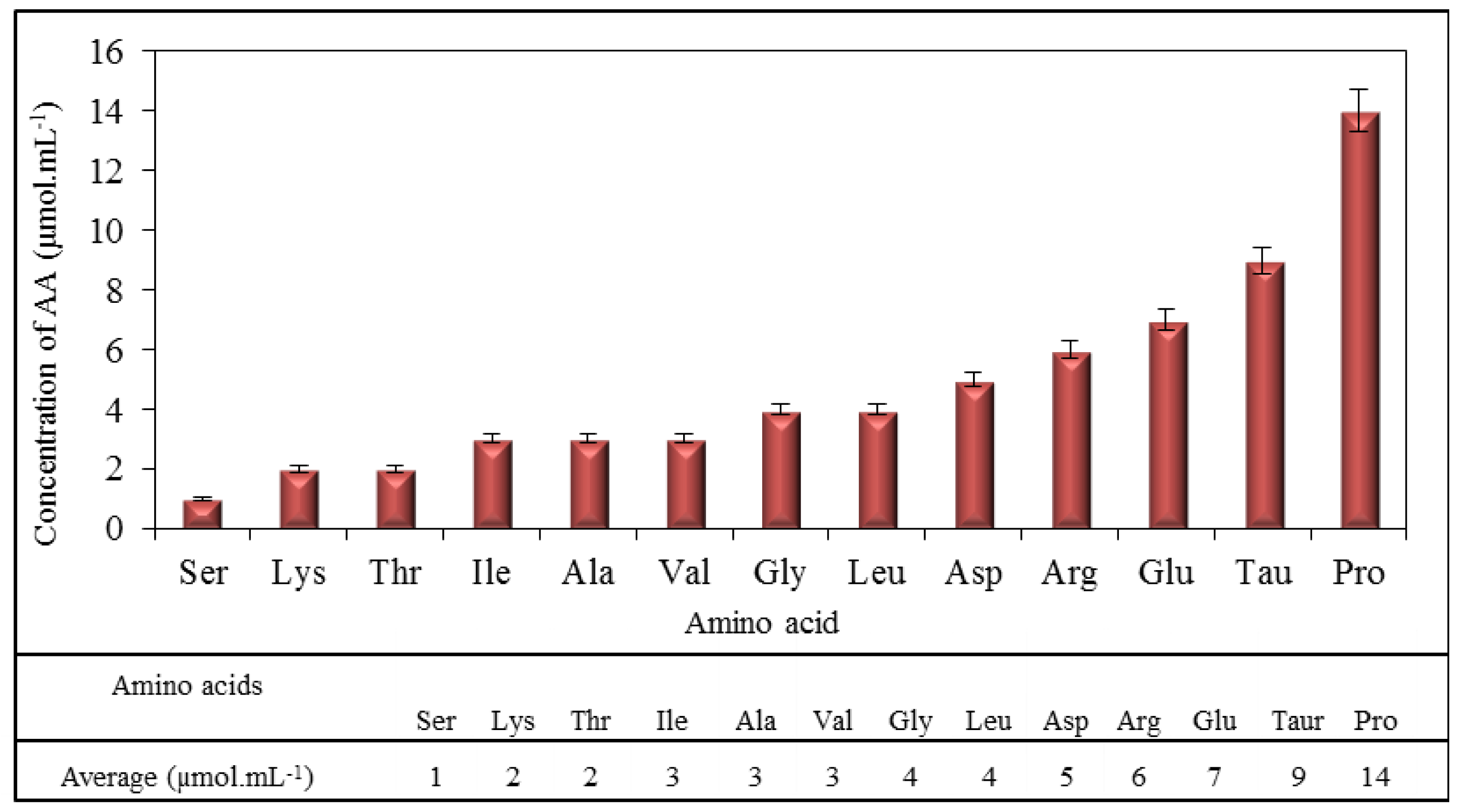
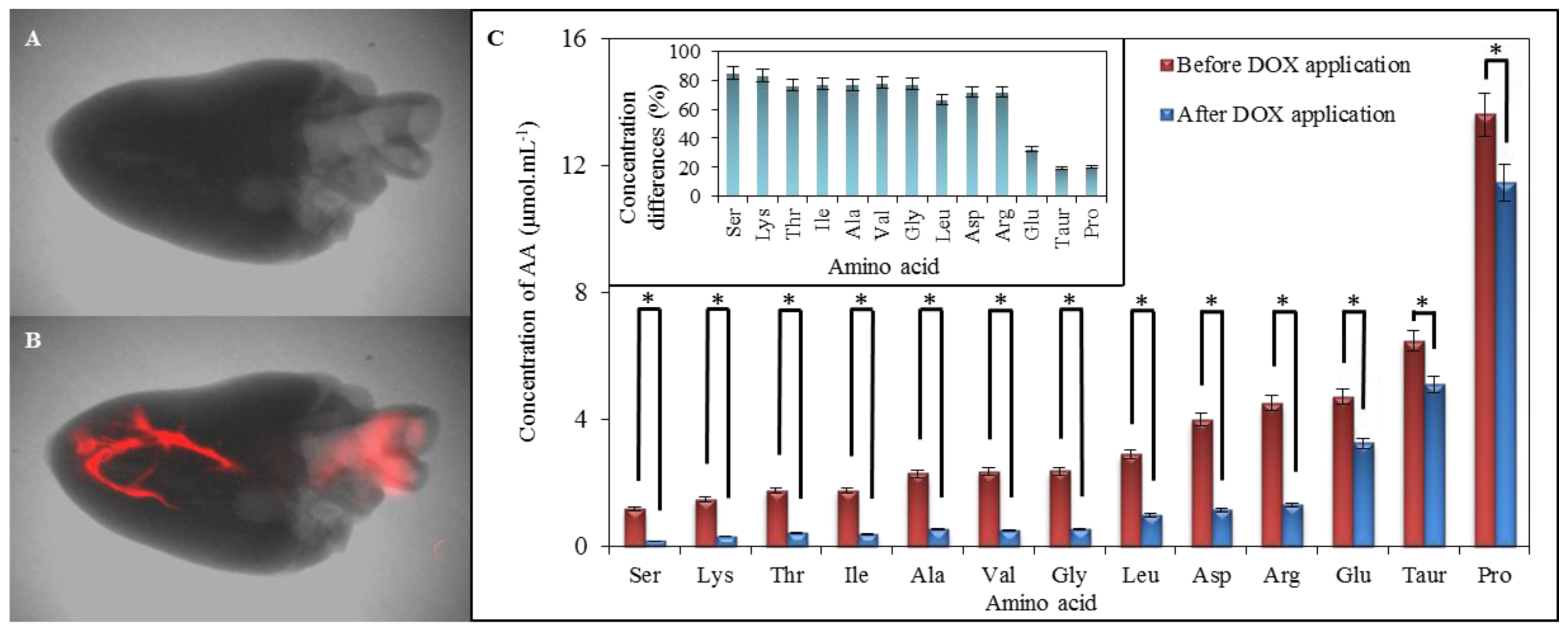
2.1. Amino Acid Profile of Chicken Myocardium
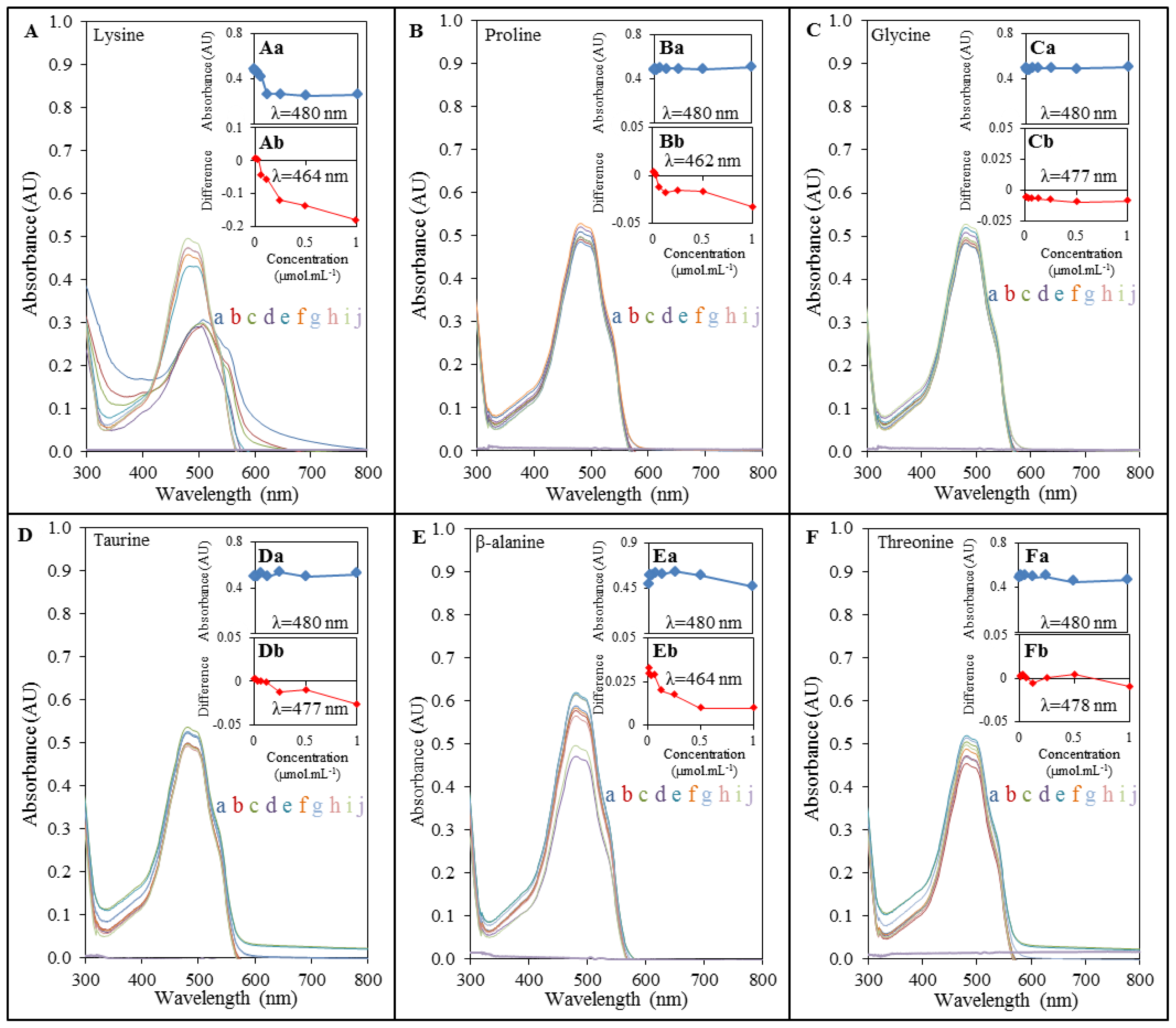
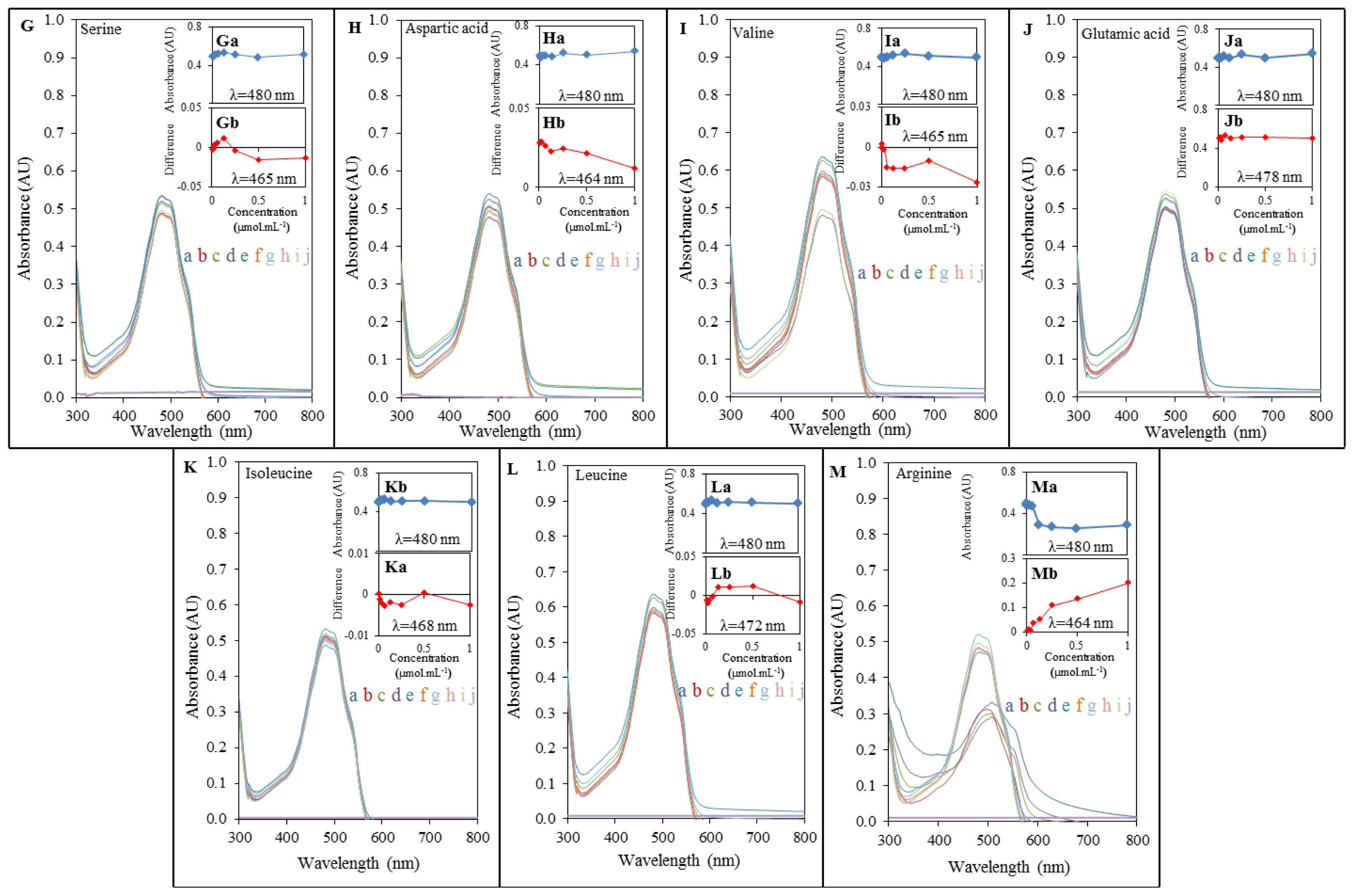
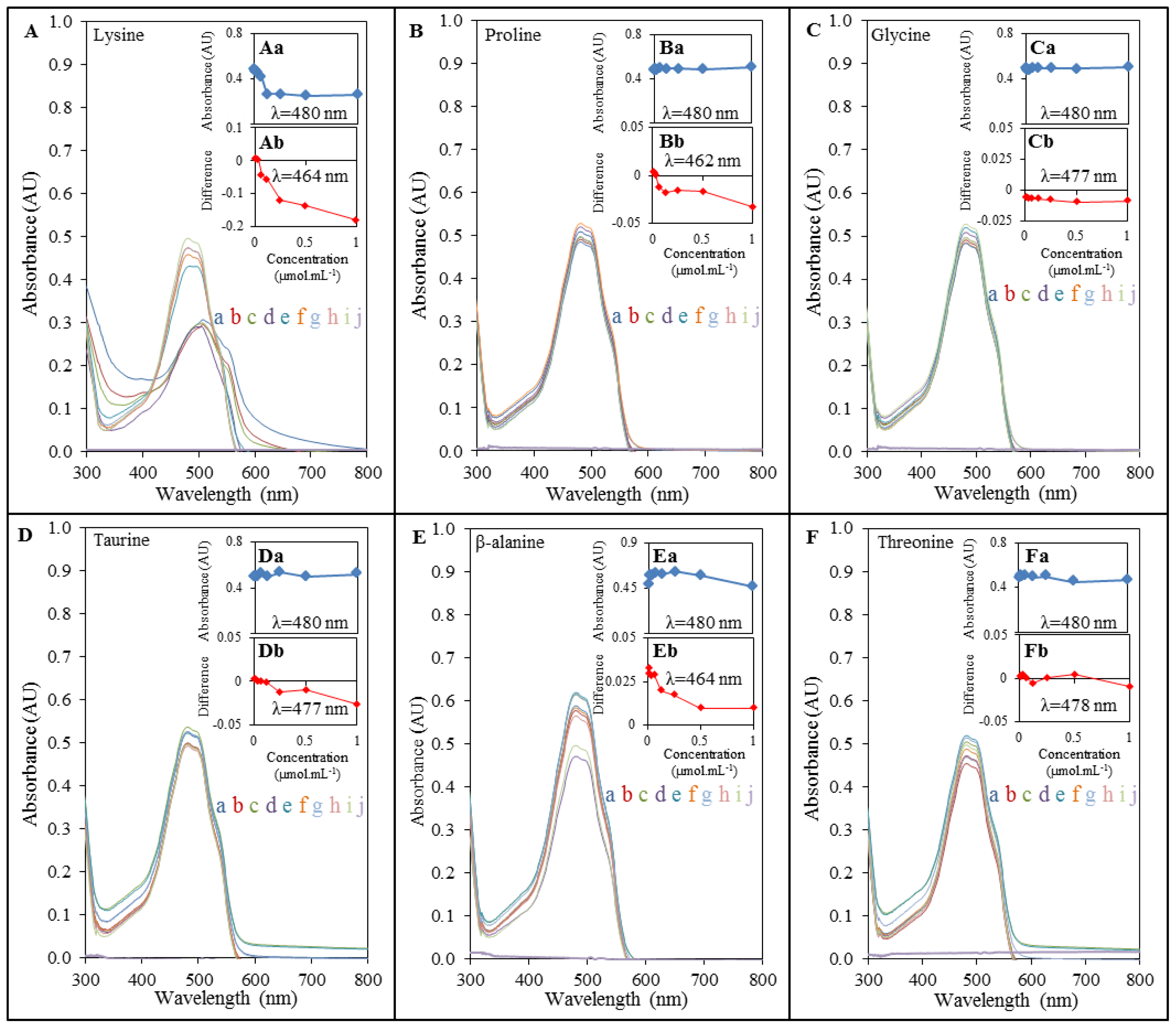
2.2. Spectrophotometric Analysis of Amino Acids-Doxorubicin Interactions
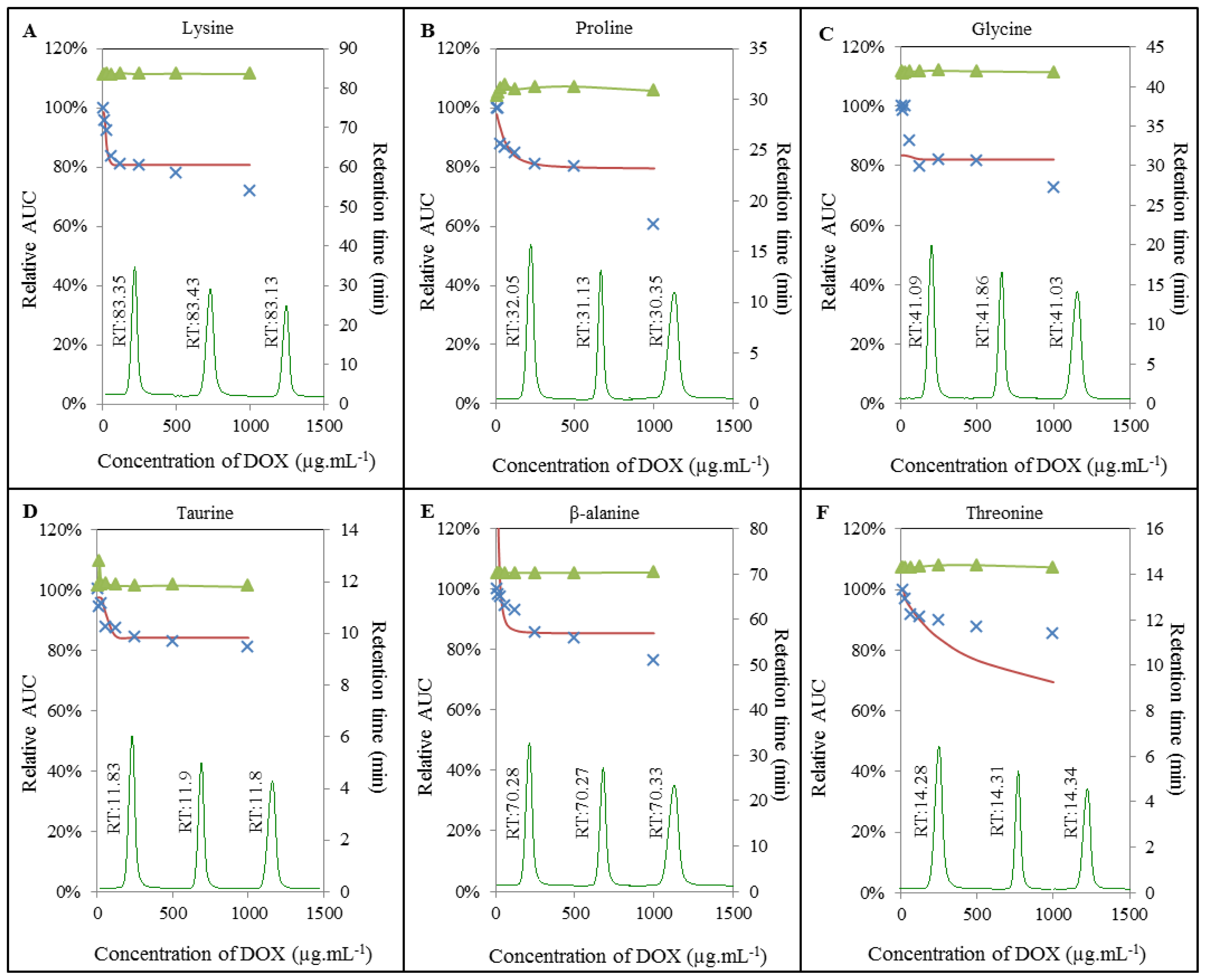
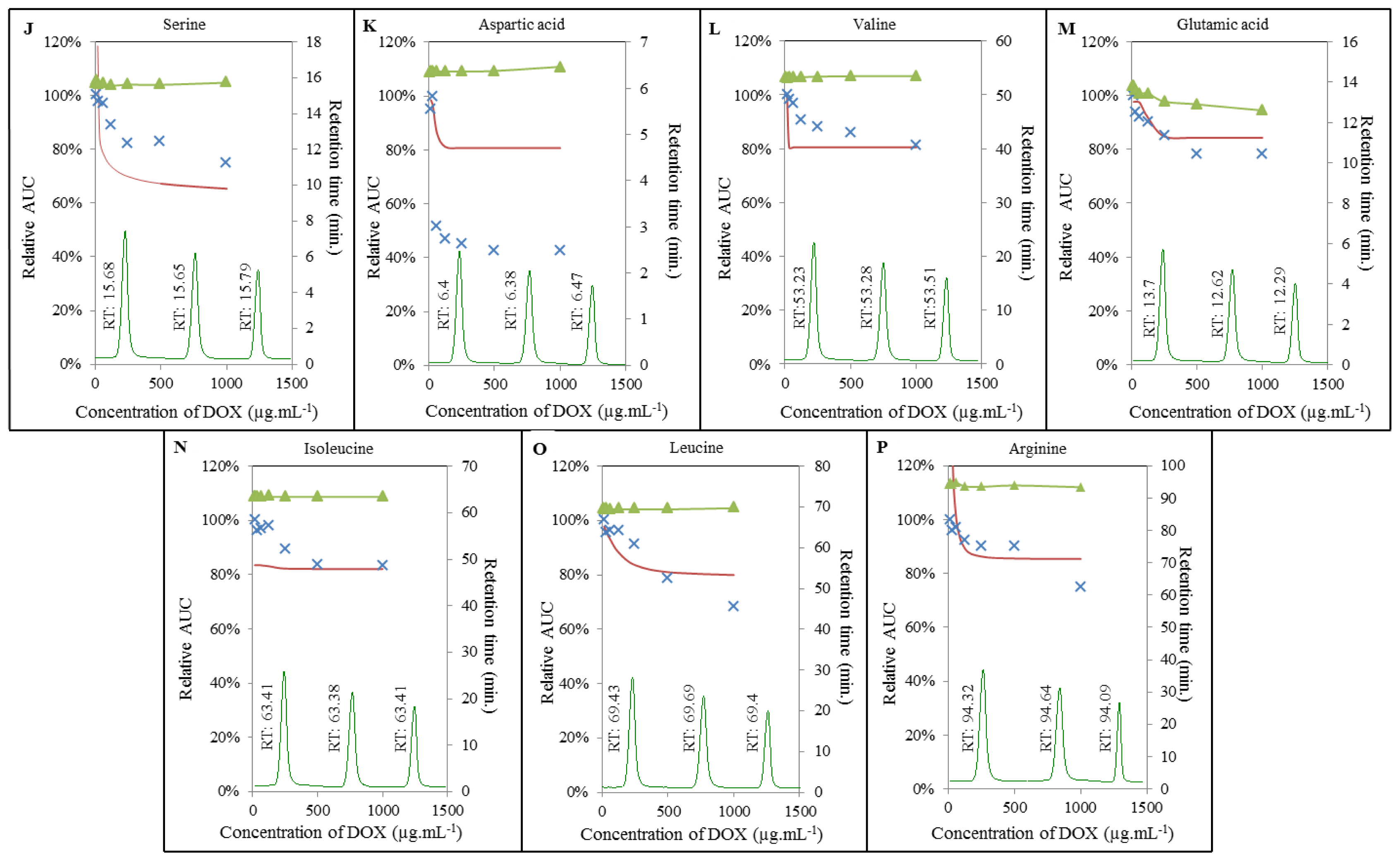
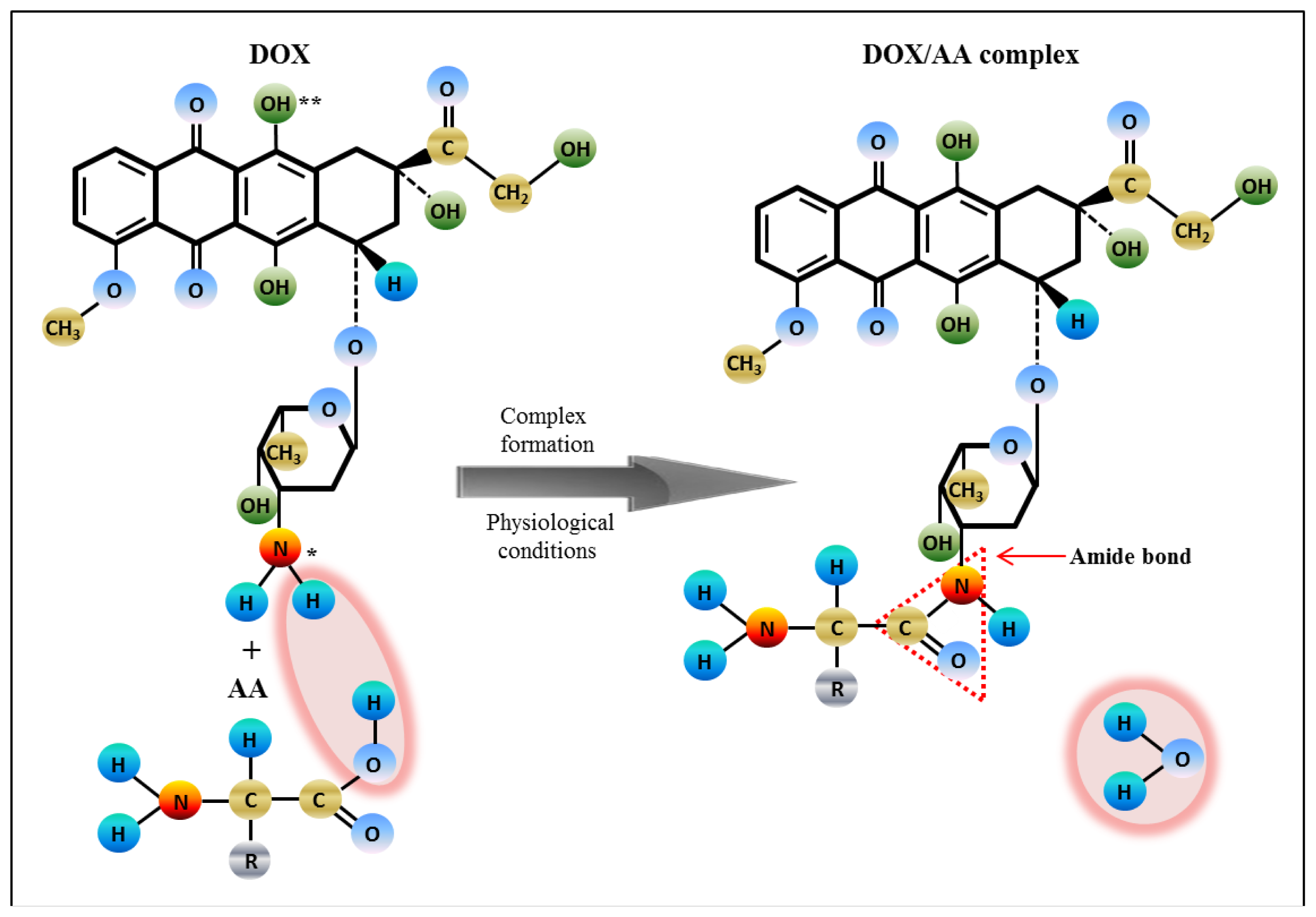
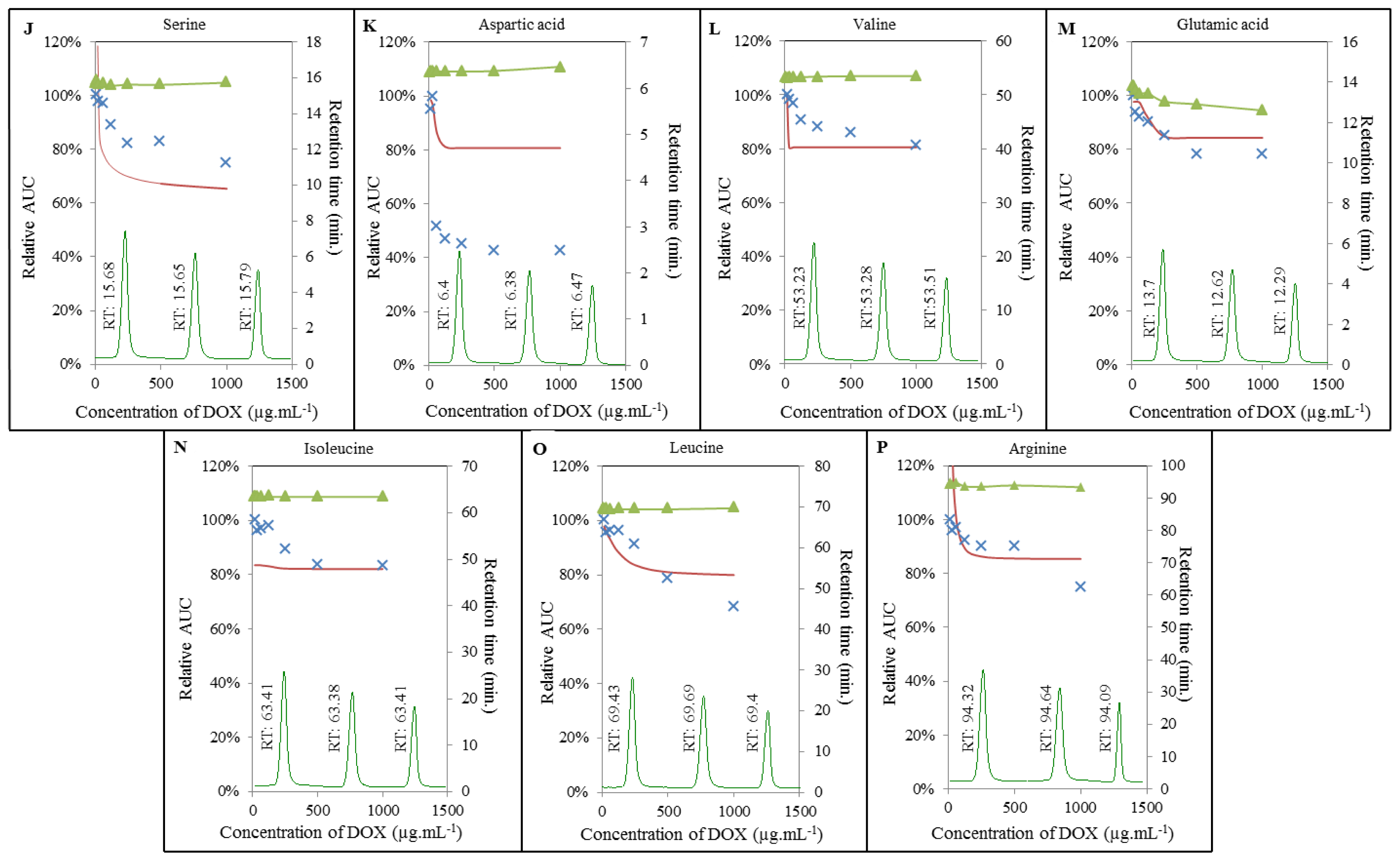
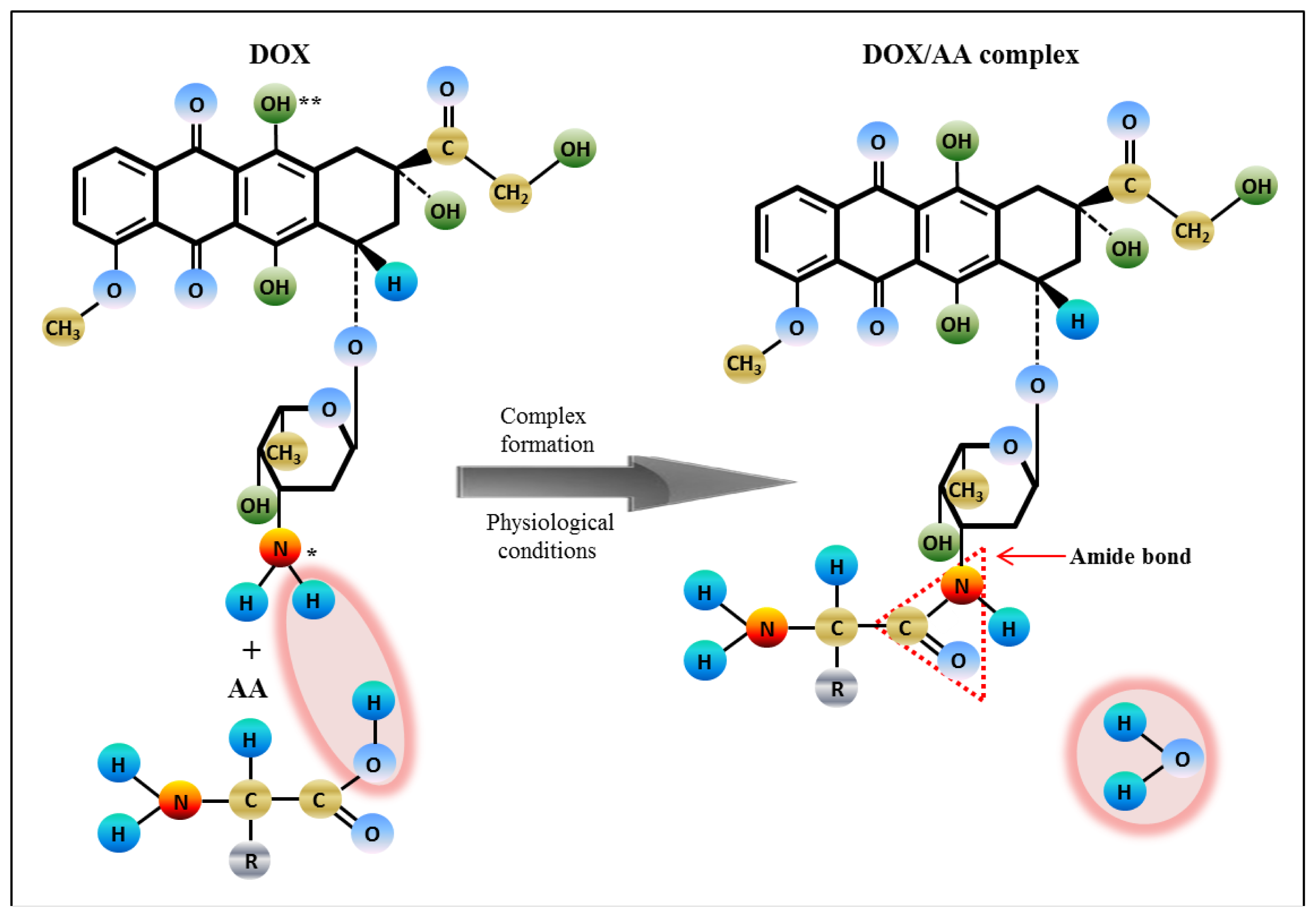
2.3. Analysis of Amino Acids-Doxorubicin Interactions by IELC
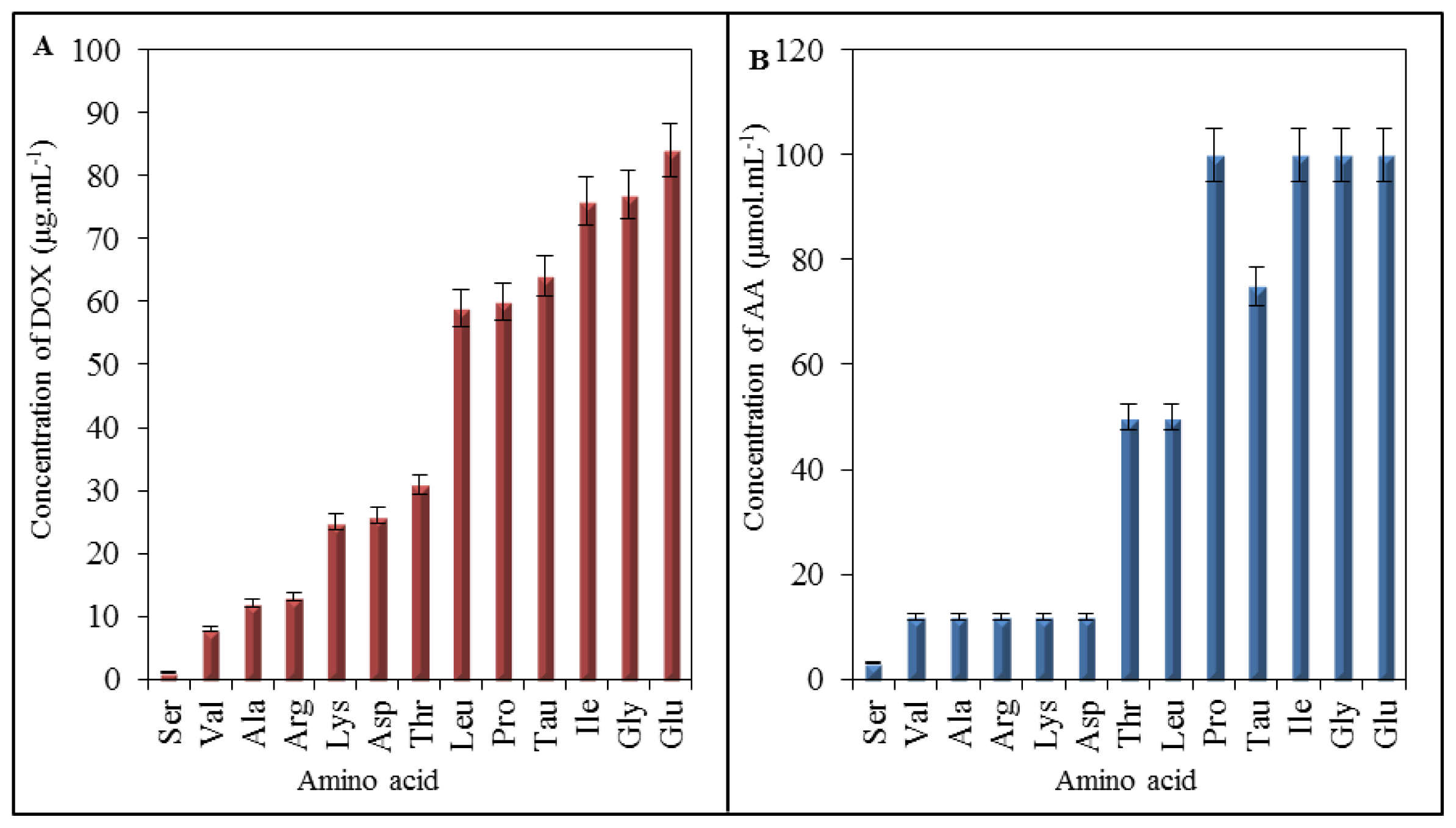
2.4. Effect of Doxorubicin on Breaking Points of Amino Acids
2.5. Impact of Doxorubicin on Amino Acids
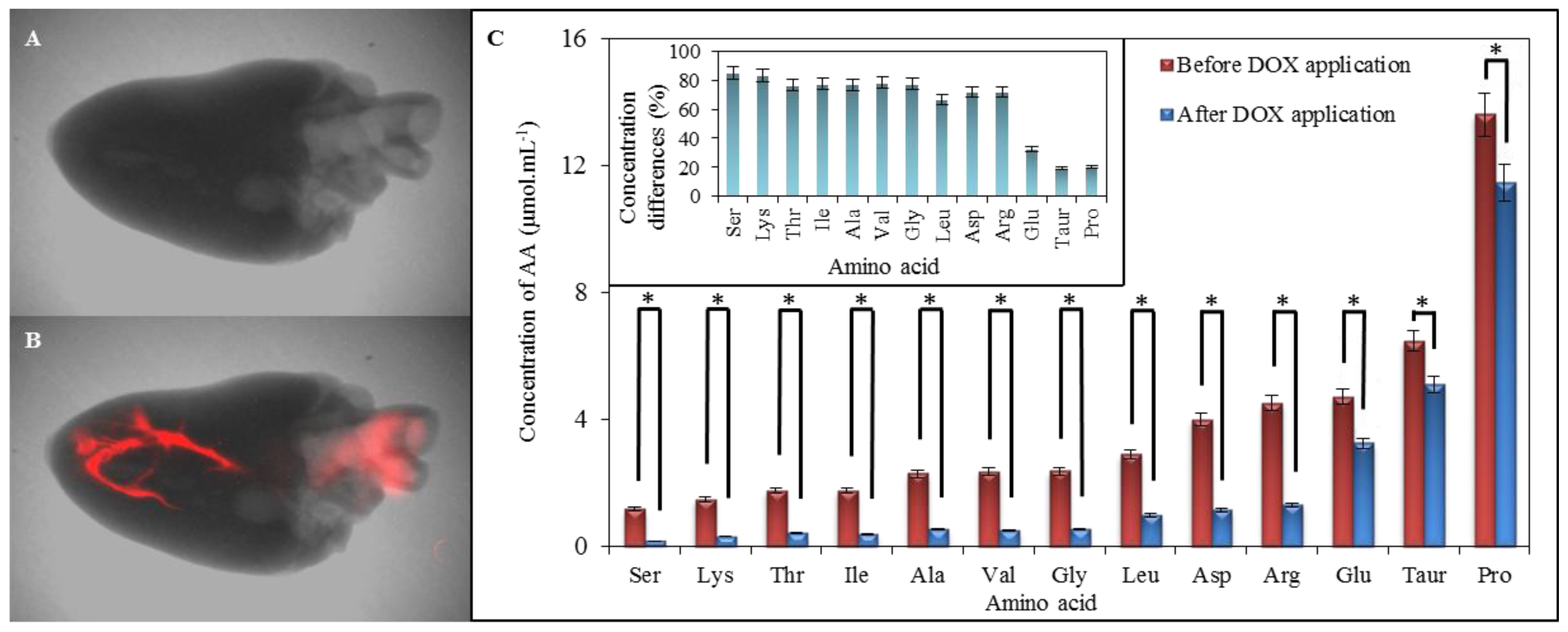
2.6. Comparison of Amino Acid Profile of Myocardium before and after Application of Doxorubicin
3. Experimental Section
3.1. Chemicals and pH Measurement
3.2. Preparation of Myocardium Samples and DOX Fluorescence Detection
3.3. Preparation of Amino Acid-Doxorubicin Sample for Interaction Study
3.4. Determination of Content of Amino Acids in Myocardium and Analysis of Interactions Using IELC
3.5. Spectrophotometric Analysis
3.6. Determination of Breaking Points of Amino Acids
3.7. Descriptive Statistics
4. Conclusions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino acid | Ser | Val | Ala | Arg | Lys | Asp | Thr | Leu | Pro | Tau | Ile | Gly | Glu |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BP (μg mL−1 of DOX) | 1 | 8 | 12 | 13 | 25 | 26 | 31 | 59 | 60 | 64 | 76 | 77 | 84 |
Acknowledgments
Abbreviations
| 4E-BP | 4E-binding protein |
| AA | Amino acids |
| BCAAs | Branched-chain amino acids |
| DNR | Daunorubicin |
| DOX | Doxorubicin |
| IELC | Ion-exchange liquid chromatography |
| mPTPs | Mitochondrial permeability transition pores |
| mTOR | Mammalian target of rapamycin |
| ROS | Reactive oxygen species |
| S6K | S6 kinase |
| SREBP | Sterol response element binding protein. |
Conflicts of Interest
References
- Das, J.; Ghosh, J.; Manna, P.; Sil, P.C. Taurine protects rat testes against doxorubicin-induced oxidative stress as well as p53, Fas and caspase 12-mediated apoptosis. Amino Acids 2012, 42, 1839–1855. [Google Scholar]
- Holley, A.K.; Dhar, S.K.; Xu, Y.; St Clair, D.K. Manganese superoxide dismutase: Beyond life and death. Amino Acids 2012, 42, 139–158. [Google Scholar]
- Hossain, S.; Yamamoto, H.; Chowdhury, E.H.; Wu, X.; Hirose, H.; Haque, A.; Doki, Y.; Mori, M.; Akaike, T. Fabrication and intracellular delivery of doxorubicin/carbonate apatite nanocomposites: Effect on growth retardation of established colon tumor. PLoS One 2013, 8, 1–11. [Google Scholar]
- Farolfi, A.; Melegari, E.; Aquilina, M.; Scarpi, E.; Ibrahim, T.; Maltoni, R.; Sarti, S.; Cecconetto, L.; Pietri, E.; Ferrario, C.; et al. Trastuzumab-induced cardiotoxicity in early breast cancer patients: A retrospective study of possible risk and protective factors. Heart 2013, 99, 634–639. [Google Scholar]
- Li, M.Q.; Song, W.T.; Tang, Z.H.; Lv, S.X.; Lin, L.; Sun, H.; Li, Q.S.; Yang, Y.; Hong, H.; Chen, X.S. Nanoscaled poly(l-glutamic acid)/doxorubicin-amphiphile complex as pH-responsive drug delivery system for effective treatment of nonsmall cell lung cancer. ACS Appl. Mater. Interfaces 2013, 5, 1781–1792. [Google Scholar]
- Nissen, M.J.; Tsai, M.L.; Blaes, A.H.; Swenson, K.K.; Koering, S. Effectiveness of treatment summaries in increasing breast and colorectal cancer survivors’ knowledge about their diagnosis and treatment. J. Cancer Surviv 2013, 7, 211–218. [Google Scholar]
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: Molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev 2004, 56, 185–229. [Google Scholar]
- Yuan, A.; Wu, J.H.; Song, C.C.; Tang, X.L.; Qiao, Q.; Zhao, L.L.; Gong, G.M.; Hu, Y.Q. A novel self-assembly albumin nanocarrier for reducing doxorubicin-mediated cardiotoxicity. J. Pharm. Sci 2013, 102, 1626–1635. [Google Scholar]
- Box, V.G.S. The intercalation of DNA double helices with doxorubicin and nagalomycin. J. Mol. Gr. Model 2007, 26, 14–19. [Google Scholar]
- Swift, L.P.; Rephaeli, A.; Nudelman, A.; Phillips, D.R.; Cutts, S.M. Doxorubicin-DNA adducts induce a non-topoisomerase II-mediated form of cell death. Cancer Res 2006, 66, 4863–4871. [Google Scholar]
- Schwartz, R.G.; Jain, D.; Storozynsky, E. Traditional and novel methods to assess and prevent chemotherapy-related cardiac dysfunction noninvasively. J. Nuclear Cardiol 2013, 20, 443–464. [Google Scholar]
- Kizek, R.; Adam, V.; Hrabeta, J.; Eckschlager, T.; Smutny, S.; Burda, J.V.; Frei, E.; Stiborova, M. Anthracyclines and ellipticines as DNA-damaging anticancer drugs: Recent advances. Pharmacol. Ther 2012, 133, 26–39. [Google Scholar]
- Perego, P.; Corna, E.; de Cesare, M.; Gatti, L.; Polizzi, D.; Pratesi, G.; Supino, R.; Zunino, F. Role of apoptosis and apoptosis-related genes in cellular response and antitumor efficacy of anthracyclines. Curr. Med. Chem 2001, 8, 31–37. [Google Scholar]
- Yang, E.S.; Huh, Y.J.; Park, J.W. RNA interference targeting sensitive-to-apoptosis gene potentiates doxorubicin- and staurosporine-induced apoptosis of PC3 cells. Anticancer Res 2013, 33, 847–855. [Google Scholar]
- Wallace, K.B. Doxorubicin-induced cardiac mitochondrionopathy. Pharm. Toxicol 2003, 93, 105–115. [Google Scholar]
- Sardao, V.A.; Oliveira, P.J.; Holy, J.; Oliveira, C.R.; Wallace, K.B. Doxorubicin-induced mitochondrial dysfunction is secondary to nuclear p53 activation in H9c2 cardiomyoblasts. Cancer Chemother. Pharm 2009, 64, 811–827. [Google Scholar]
- Toldo, S.; Goehe, R.W.; Lotrionte, M.; Mezzaroma, E.; Sumner, E.T.; Biondi-Zoccai, G.G.L.; Seropian, I.M.; van Tassell, B.W.; Loperfido, F.; Palazzoni, G.; et al. Comparative cardiac toxicity of anthracyclines in vitro and in vivo in the mouse. PLoS One 2013, 8, 1–8. [Google Scholar]
- Fong, M.Y.; Jin, S.Y.; Rane, M.; Singh, R.K.; Gupta, R.; Kakar, S.S. Withaferin a synergizes the therapeutic effect of doxorubicin through ROS-mediated autophagy in ovarian cancer. PLoS One 2012, 7, 1–16. [Google Scholar]
- Gharanei, M.; Hussain, A.; Janneh, O.; Maddock, H.L. Doxorubicin induced myocardial injury is exacerbated following ischaemic stress via opening of the mitochondrial permeability transition pore. Toxicol. Appl. Pharm 2013, 268, 149–156. [Google Scholar]
- Yang, J.Q.; Maity, B.; Huang, J.; Gao, Z.; Stewart, A.; Weiss, R.M.; Anderson, M.E.; Fisher, R.A. G-protein inactivator RGS6 mediates myocardial cell apoptosis and cardiomyopathy caused by doxorubicin. Cancer Res 2013, 73, 1662–1667. [Google Scholar]
- Yao, Z.W.; Hu, W.; Yin, S.; Huang, Z.; Zhu, Q.; Chen, J.N.; Zang, Y.H.; Dong, L.; Zhang, J.F. 3,3′-Diindolymethane ameliorates adriamycin-induced cardiac fibrosis via activation of a BRCA1-dependent anti-oxidant pathway. Pharm. Res 2013, 70, 139–146. [Google Scholar]
- Zeng, Q.B.; Wen, H.B.; Wen, Q.; Chen, X.H.; Wang, Y.G.; Xuan, W.L.; Liang, J.S.; Wan, S.H. Cucumber mosaic virus as drug delivery vehicle for doxorubicin. Biomaterials 2013, 34, 4632–4642. [Google Scholar]
- Eckman, D.M.; Stacey, R.B.; Rowe, R.; D’Agostino, R.; Kock, N.D.; Sane, D.C.; Torti, F.M.; Yeboah, J.; Workman, S.; Lane, K.S.; et al. Weekly doxorubicin increases coronary arteriolar wall and adventitial thickness. PLoS One 2013, 8, 1–6. [Google Scholar]
- Pereira Neto, G.B.; Andrade, J.N.B.; Sousa, M.G.; Camacho, A.A. Holter electrocardiography in dogs showing doxorubicin-induced dilated cardiomyopathy. Arq. Bras. Med. Vet. Zootec 2006, 58, 1037–1042. [Google Scholar]
- Lim, C.C.; Zuppinger, C.; Guo, X.X.; Kuster, G.M.; Helmes, M.; Eppenberger, H.M.; Suter, T.M.; Liao, R.L.; Sawyer, D.B. Anthracyclines induce calpain-dependent titin proteolysis and necrosis in cardiomyocytes. J. Biol. Chem 2004, 279, 8290–8299. [Google Scholar]
- Liu, T.C.K.; Ismail, S.; Brennan, O.; Hastings, C.; Duffy, G.P. Encapsulation of cardiac stem cells in superoxide dismutase-loaded alginate prevents doxorubicin-mediated toxicity. J. Tissue Eng. Regen. Med 2013, 7, 302–311. [Google Scholar]
- Masarik, M.; Kynclova, H.; Huska, D.; Hubalek, J.; Adam, V.; Babula, P.; Eckschlager, T.; Stiborova, M.; Kizek, R. DNA-doxorubicin interactions revealed by electrochemistry. Int. J. Mol. Med 2010, 26, 46. [Google Scholar]
- Feridooni, T.; Hotchkiss, A.; Remley-Carr, S.; Saga, Y.; Pasumarthi, K.B.S. Cardiomyocyte specific ablation of p53 is not sufficient to block doxorubicin induced cardiac fibrosis and associated cytoskeletal changes. PLoS One 2011, 6, 1–12. [Google Scholar]
- Drake, K.J.; Sidorov, V.Y.; McGuinness, O.P.; Wasserman, D.H.; Wikswo, J.P. Amino acids as metabolic substrates during cardiac ischemia. Exp. Biol. Med 2012, 237, 1369–1378. [Google Scholar]
- Nejdl, L.; Sochor, J.; Zitka, O.; Cernei, N.; Ruttkay-Nedecky, B.; Kopel, P.; Babula, P.; Adam, V.; Hubalek, J.; Kizek, R. Spectrometric and chromatographic study of reactive oxidants hypochlorous and hypobromous acids and their interactions with taurine. Chromatographia 2013, 76, 363–373. [Google Scholar]
- Huang, Y.; Zhou, M.Y.; Sun, H.P.; Wang, Y.B. Branched-chain amino acid metabolism in heart disease: An epiphenomenon or a real culprit? Cardiovasc. Res 2011, 90, 220–223. [Google Scholar]
- Weitzel, L.B.; Ambardekar, A.V.; Brieke, A.; Cleveland, J.C.; Serkova, N.J.; Wischmeyer, P.E.; Lowes, B.D. Left ventricular assist device effects on metabolic substrates in the failing heart. PLoS One 2013, 8, 1–6. [Google Scholar]
- Tazina, E.V.; Ignatieva, E.V.; Polozkova, A.P.; Oborotova, N.A. Qualitative and quantitative analysis of thermosensitive liposomes loaded with doxorubicin. Pharm. Chem. J 2012, 46, 54–59. [Google Scholar]
- Albright, C.F.; Graciani, N.; Han, W.; Yue, E.; Stein, R.; Lai, Z.H.; Diamond, M.; Dowling, R.; Grimminger, L.; Zhang, S.Y.; et al. Matrix metalloproteinase-activated doxorubicin prodrugs inhibit HT1080 xenograft growth doxorubicin with less toxicity. Mol. Cancer Ther 2005, 4, 751–760. [Google Scholar]
- Manocha, B.; Margaritis, A. Controlled release of doxorubicin from doxorubicin/γ-polyglutamic acid ionic complex. J. Nanomater 2010, 2010, 1–9. [Google Scholar]
- Taetrneyer, H.; Harinstein, M.E.; Gheorghiade, M. More than bricks and mortar: Comments on protein and amino acid metabolism in the heart. Am. J. Cardiol 2008, 101, 3E–7E. [Google Scholar]
- Proud, C.G. Regulation of mammalian translation factors by nutrients. Eur. J. Biochem 2002, 269, 5338–5349. [Google Scholar]
- Zhang, D.H.; Contu, R.; Latronico, M.V.G.; Zhang, J.A.L.; Rizzi, R.; Catalucci, D.; Miyamoto, S.; Huang, K.; Ceci, M.; Gu, Y.S.; et al. MTORC1 regulates cardiac function and myocyte survival through 4E-BP1 inhibition in mice. J. Clin. Investig 2010, 120, 2805–2816. [Google Scholar]
- Katta, A.; Kundla, S.; Kakarla, S.K.; Wu, M.Z.; Fannin, J.; Paturi, S.; Liu, H.; Addagarla, H.S.; Blough, E.R. Impaired overload-induced hypertrophy is associated with diminished mTOR signaling in insulin-resistant skeletal muscle of the obese Zucker rat. Am. J. Physiol. Regul. Integr. Comp. Physiol 2010, 299, R1666–R1675. [Google Scholar]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar]
- Porstmann, T.; Santos, C.R.; Griffiths, B.; Cully, M.; Wu, M.; Leevers, S.; Griffiths, J.R.; Chung, Y.L.; Schulze, A. SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metab 2008, 8, 224–236. [Google Scholar]
- Zima, T.; Tesar, V.; Mantle, D.; Koll, M.; Patel, V.; Richardson, P.J.; Preedy, V.R. Acute doxorubicin (adriamycin) dosage does not reduce cardiac protein synthesis in vivo, but decreases diaminopeptidase I and proline endopeptidase activities. Exp. Mol. Pathol 2001, 70, 154–161. [Google Scholar]
- Takemura, G.; Fujiwara, H. Doxorubicin-induccd cardiomyopathy from the cardiotoxic mechanisms to management. Prog. Cardiovasc. Dis 2007, 49, 330–352. [Google Scholar]
- Gianni, L.; Herman, E.H.; Lipshultz, S.E.; Minotti, G.; Sarvazyan, N.; Sawyer, D.B. Anthracycline cardiotoxicity: From bench to bedside. J. Clin. Oncol 2008, 26, 3777–3784. [Google Scholar]
- Yellon, D.M.; Hausenloy, D.J. Mechanisms of disease: Myocardial reperfusion injury. N. Engl. J. Med 2007, 357, 1121–1135. [Google Scholar]
- Halestrap, A.P. What is the mitochondrial permeability transition pore? J. Mol. Cell. Cardiol 2009, 46, 821–831. [Google Scholar]
- Halestrap, A.P.; Pasdois, P. The role of the mitochondrial permeability transition pore in heart disease. Biochim. Biophys. Acta 2009, 1787, 1402–1415. [Google Scholar]
- Gava, F.N.; Zacche, E.; Ortiz, E.M.G.; Champion, T.; Bandarra, M.B.; Vasconcelos, R.O.; Barbosa, J.C.; Camacho, A.A. Doxorubicin induced dilated cardiomyopathy in a rabbit model: An update. Res. Vet. Sci 2013, 94, 115–121. [Google Scholar]
- She, P.X.; Olson, K.C.; Kadota, Y.; Inukai, A.; Shimomura, Y.; Hoppel, C.L.; Adams, S.H.; Kawamata, Y.; Matsumoto, H.; Sakai, R.; et al. Leucine and protein metabolism in obese zucker rats. PLoS One 2013, 8, 1–19. [Google Scholar]
- Wilkinson, D.J.; Hossain, T.; Hill, D.S.; Phillips, B.E.; Crossland, H.; Williams, J.; Loughna, P.; Churchward-Venne, T.A.; Breen, L.; Phillips, S.M.; et al. Effects of leucine and its metabolite β-hydroxy-β-methylbutyrate on human skeletal muscle protein metabolism. J. Physiol 2013, 591, 2911–2923. [Google Scholar]
- Yoo, H.S.; Park, T.G. Folate-receptor-targeted delivery of doxorubicin nano-aggregates stabilized by doxorubicin-PEG-folate conjugate. J. Control. Release 2004, 100, 247–256. [Google Scholar]
- Ryppa, C.; Mann-Steinberg, H.; Fichtner, I.; Weber, H.; Satchi-Fainaro, R.; Biniossek, M.L.; Kratz, F. In vitro and in vivo evaluation of doxorubicin conjugates with the divalent peptide E-[c(RGDfK)2] that targets integrin αvβ3. Bioconjug. Chem 2008, 19, 1414–1422. [Google Scholar]
- King, H.D.; Dubowchik, G.M.; Mastalerz, H.; Willner, D.; Hofstead, S.J.; Firestone, R.A.; Lasch, S.J.; Trail, P.A. Monoclonal antibody conjugates of doxorubicin prepared with branched peptide linkers: Inhibition of aggregation by methoxytriethyleneglycol chains. J. Med. Chem 2002, 45, 4336–4343. [Google Scholar]
- Agudelo, D.; Bourassa, P.; Bruneau, J.; Berube, G.; Asselin, E.; Tajmir-Riahi, H.A. Probing the binding sites of antibiotic drugs doxorubicin and N-(trifluoroacetyl) doxorubicin with human and bovine serum albumins. PLoS One 2012, 7, 1–13. [Google Scholar]
- Long, G.L.; Winefordner, J.D. Limit of detection. Anal. Chem 1983, 55, A712–A724. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Heger, Z.; Cernei, N.; Kudr, J.; Gumulec, J.; Blazkova, I.; Zitka, O.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R. A Novel Insight into the Cardiotoxicity of Antineoplastic Drug Doxorubicin. Int. J. Mol. Sci. 2013, 14, 21629-21646. https://doi.org/10.3390/ijms141121629
Heger Z, Cernei N, Kudr J, Gumulec J, Blazkova I, Zitka O, Eckschlager T, Stiborova M, Adam V, Kizek R. A Novel Insight into the Cardiotoxicity of Antineoplastic Drug Doxorubicin. International Journal of Molecular Sciences. 2013; 14(11):21629-21646. https://doi.org/10.3390/ijms141121629
Chicago/Turabian StyleHeger, Zbynek, Natalia Cernei, Jiri Kudr, Jaromir Gumulec, Iva Blazkova, Ondrej Zitka, Tomas Eckschlager, Marie Stiborova, Vojtech Adam, and Rene Kizek. 2013. "A Novel Insight into the Cardiotoxicity of Antineoplastic Drug Doxorubicin" International Journal of Molecular Sciences 14, no. 11: 21629-21646. https://doi.org/10.3390/ijms141121629