1. Introduction
Ovarian cancer is a malignant tumor that represents a serious threat to women’s health and is one of the leading causes of cancer-related deaths in females. More than 90% of ovarian cancers are classified as epithelial and are believed to arise from the ovarian epithelium [
1]; however, evidence suggests that the fallopian tube inner surface epithelium may also be the origin of some ovarian cancers [
2]. The risk factors for ovarian cancer include family history of ovarian carcinoma and genetic mutations. Ovarian cancer has a disproportionately poor survival rate due to the late onset of symptoms. Increased understanding of the relevant molecular mechanisms that regulate ovarian carcinogenesis may result in improved methods of diagnosis, treatment, and prevention.
Ras homolog gene family member A (
RhoA) is a small (~22 kDa) G protein/guanosine triphosphatase that is part of the Ras-related C3 botulinum toxin substrate (Rac) subfamily of the Rho family [
3].
RhoA can reorganize the cell cytoskeleton and regulate cell migration by activating effector proteins such as Rho-associated coiled-coil kinase (ROCK) [
4]; such changes are associated with tumor invasion and migration in several types of carcinoma cells [
5,
6]. Phosphoinositide 3-kinase/protein kinase B (PI3K/Akt)-dependent phosphorylation of glycogen synthase kinase-3β (GSK3β) and RhoA activation regulate Wnt-5a–induced gastric cancer cell migration [
7], in line with results for breast cancer cells [
8].
Wnt-5a is a representative ligand for the seven-transmembrane receptor frizzled-5 and the tyrosine kinase orphan receptor 2, activating β-catenin–independent pathways and regulating a variety of cellular functions, such as proliferation, differentiation, migration, adhesion, and polarity [
9]. Marked expression of Wnt-5a has previously been reported in human metastatic melanoma and a variety of human primary tumor samples. However, the exact nature of its role in ovarian carcinoma remains undetermined.
To clarify the role of RhoA and Wnt-5a in ovarian carcinogenesis and subsequent progression, we examined their mRNA and protein expression in normal fallopian tube epithelium, benign tumors, primary ovarian carcinomas, and metastatic ovarian carcinomas simultaneously, the first time such an investigation has been carried out, and compared them with the clinicopathological features of patients with ovarian carcinoma. To study the molecular mechanisms, we observed the effects of RhoA or Wnt-5a knockdown on the phenotypes and expression of molecules regulated in ovarian carcinoma cells in vitro.
3. Discussion
As reviewed by Hall [
4],
RhoA exerts multiple functions in tumor metastasis by orchestrating the action of multiple downstream effectors and promoting the degradation and reconstruction of the extracellular matrix. Ellenbroek
et al. [
10] reported that RhoA protein was overexpressed in head and neck cancers and gastric cancer, indicating that RhoA upregulation is closely associated with carcinogenesis. Our study showed that
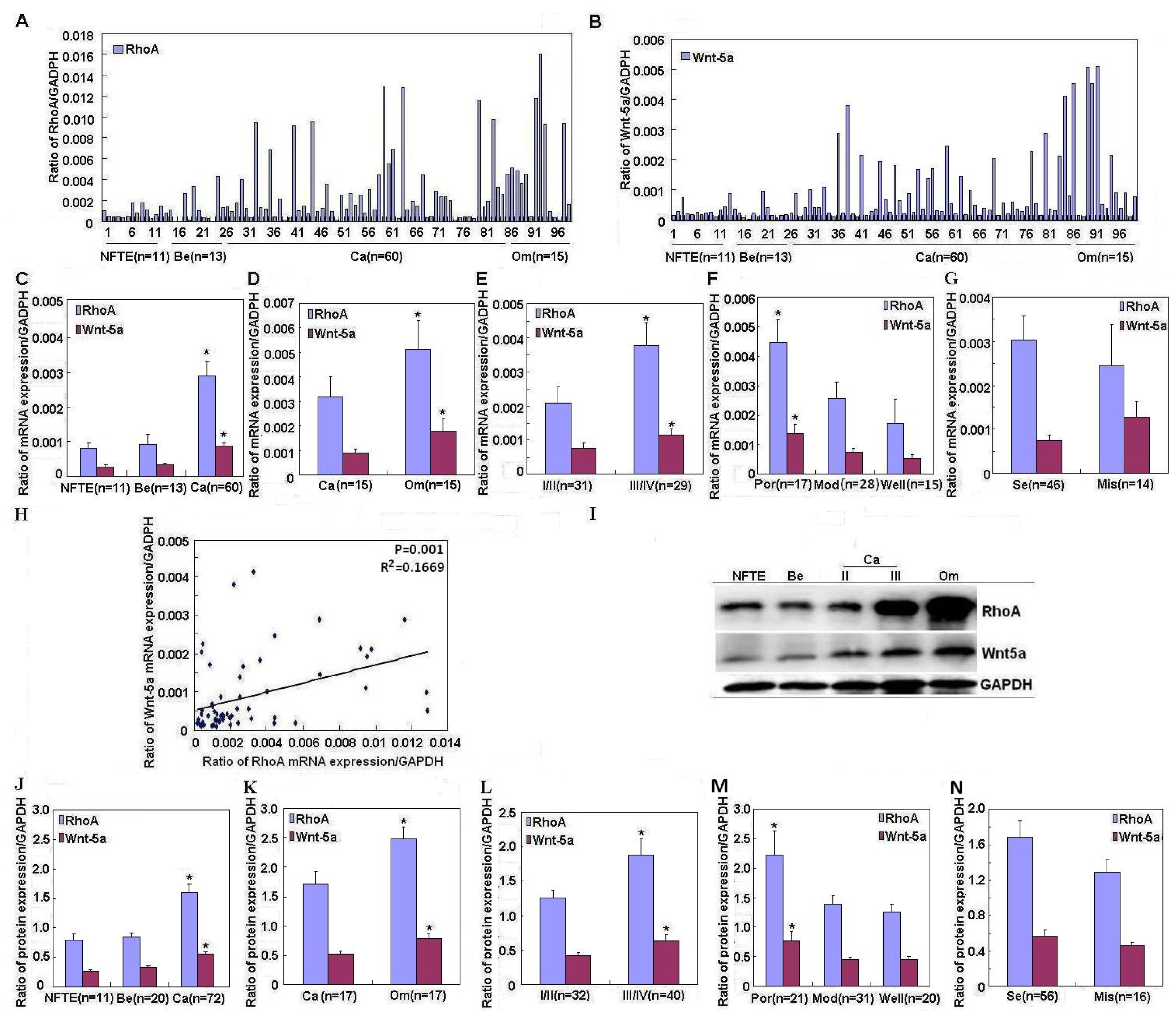
RhoA expression was significantly upregulated in ovarian carcinomas compared to normal fallopian tube epithelium and benign tumors, in accordance with Horiuchi
et al. [
11]. However,
RhoA expression was upregulated in metastatic omentum compared to ovarian carcinomas; additionally,
RhoA expression was positively associated with FIGO stage and degree of differentiation in ovarian carcinoma. These findings indicate that
RhoA overexpression might affect ovarian carcinogenesis and subsequent progression.
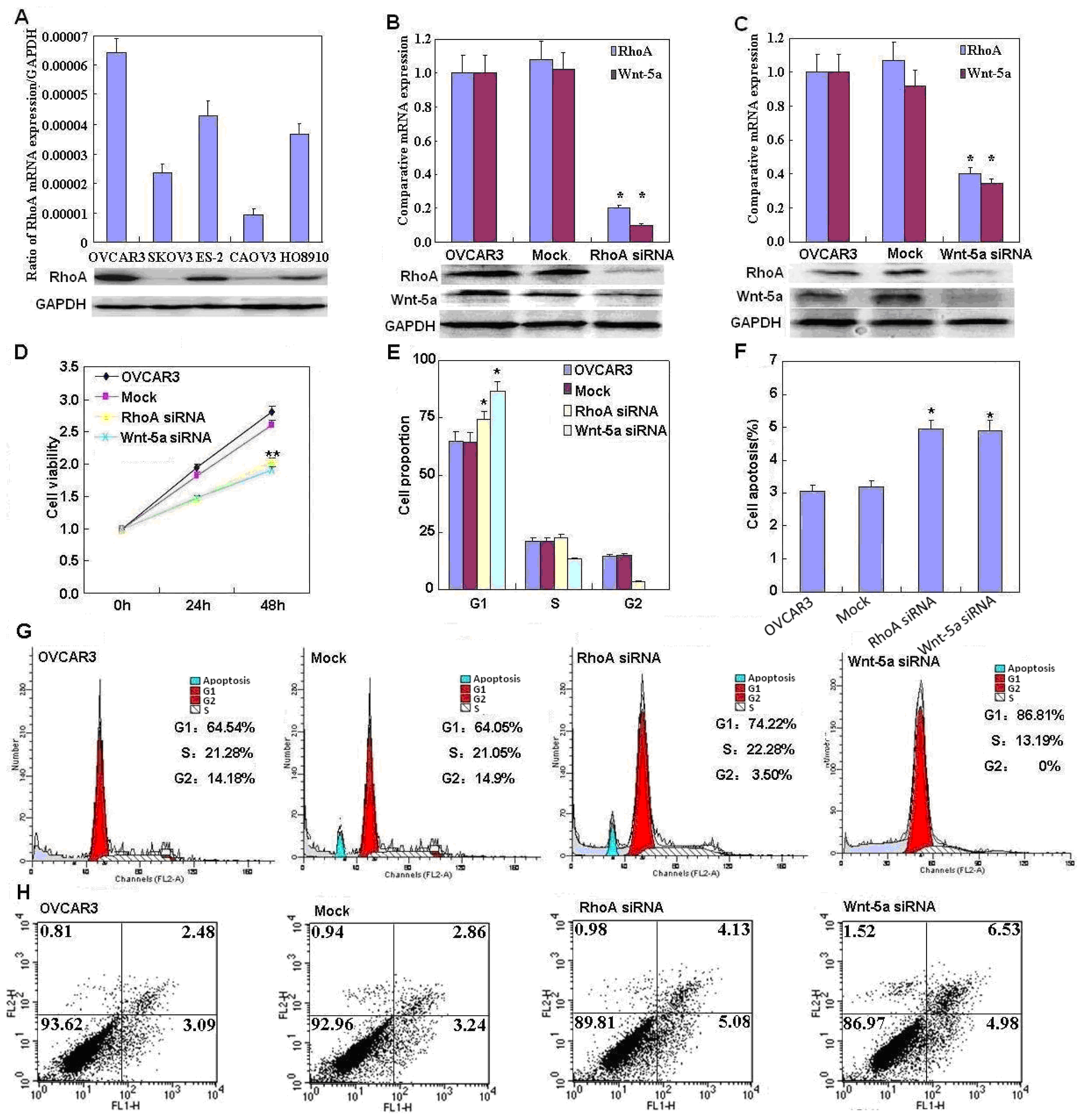
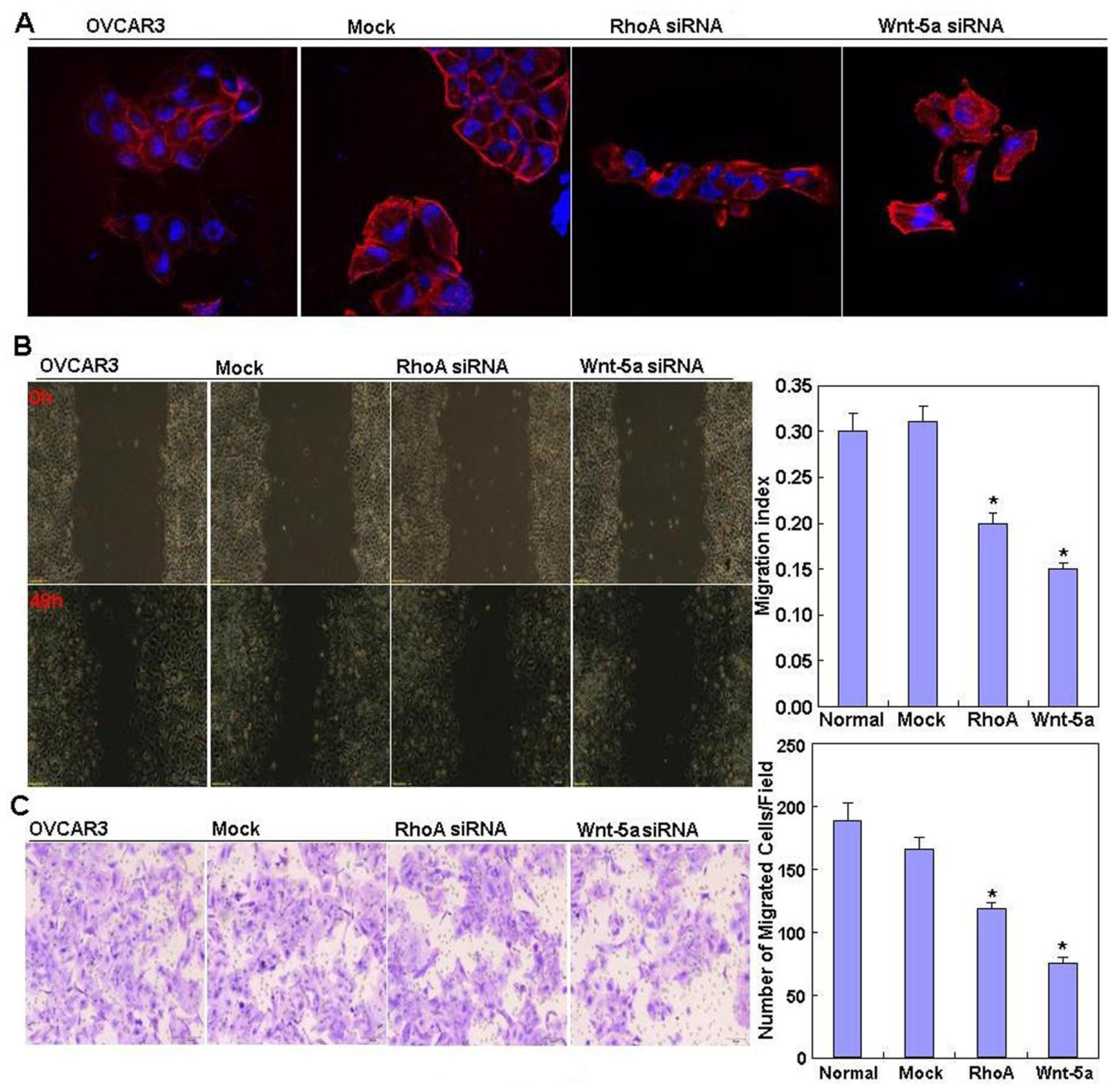
In addition, we found that RhoA knockdown induced G
1 arrest and apoptosis, decreased lamellipodia formation, and reduced cell migration and invasion, which indicates that RhoA downregulation may suppress the aggressive phenotypes of ovarian carcinoma cells. Our data are in agreement with that of Peacock
et al. [
12], who reported that
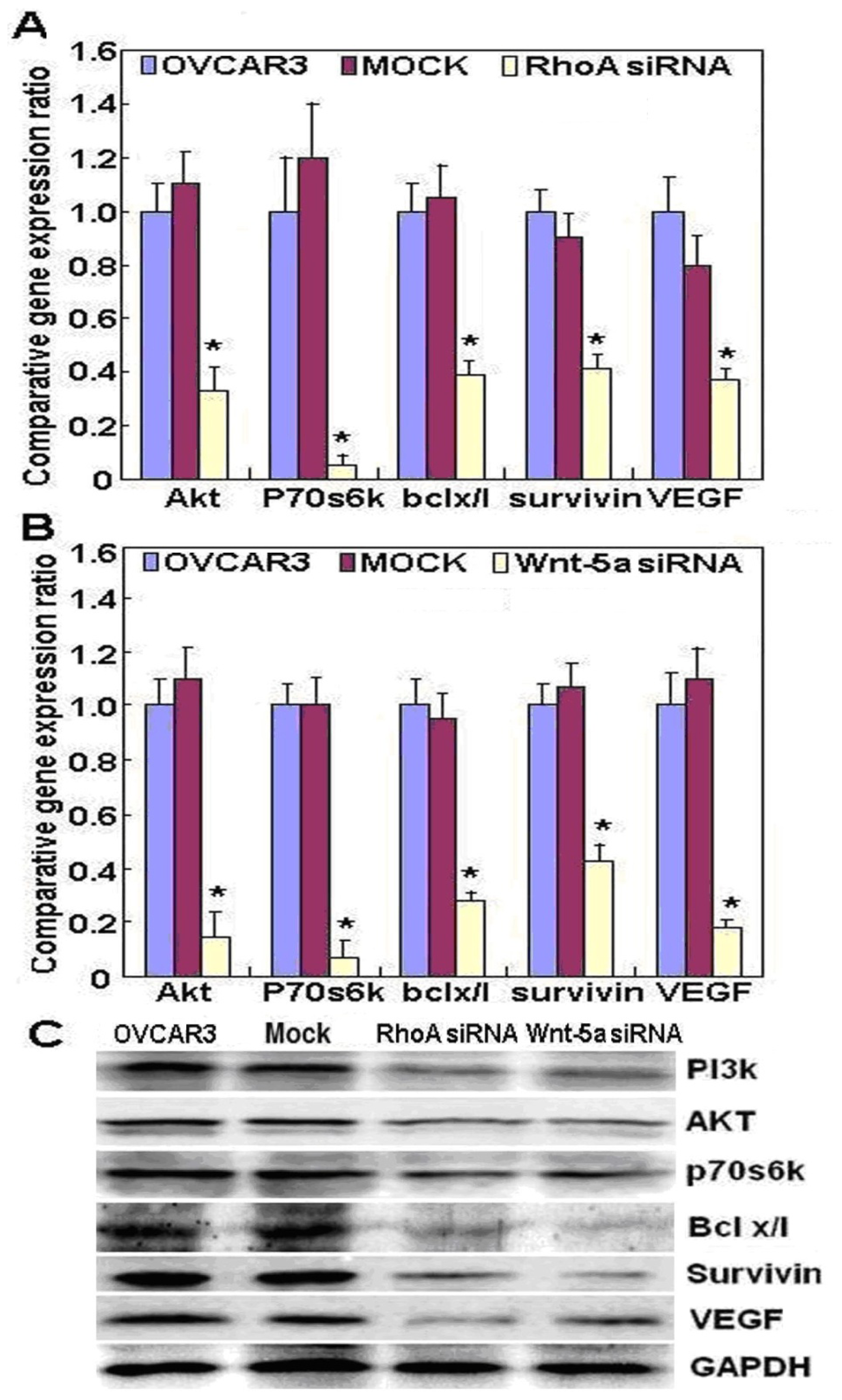
RhoA promotes reorganization of the actin cytoskeleton, regulates cell shape and attachment, and coordinates cell motility and actomyosin contractility. We also found that knockdown of RhoA significantly reduced both mRNA and protein expression of Akt, p70S6k, Bcl-xL, survivin, and VEGF in OVCAR3 cells. According to the literature, these molecules are involved in the regulation of proliferation, apoptosis, invasion, and metastasis of cancer cells [
13–
15]. Taken together, these results suggest that
RhoA may promote proliferation, anti-apoptosis, and invasion by modulating the expression of these genes.
Wnt-5a is a prototypic ligand that activates a β-catenin–independent pathway in Wnt signaling.
Wnt-5a has been demonstrated to exert differential effects on cancer development. It promotes cancer progression and metastasis in malignant melanoma, breast cancer, and gastric cancer [
8,
16–
18]. Kurayoshi
et al. [
18] reported that
Wnt-5a stimulated cell motility, invasiveness, and aggressiveness in some cancer cells, suggesting that it exerts oncogenic effects. Li
et al. [
19] also reported that
Wnt-5a exerted stem cell functions in lung cancer. In contrast, some reports indicate that in view of its ability to inhibit cell proliferation, motility, or invasion in thyroid tumor and colorectal cancer cell lines,
Wnt-5a acts as a tumor suppressor [
20–
24]. Our study showed that
Wnt-5a was also significantly upregulated in ovarian carcinomas compared to normal fallopian tube epithelium and benign tumors and in metastatic omentum compared to primary carcinomas, in line with Peng
et al. [
25] and Badiglian
et al. [
26], indicating that
Wnt-5a may be involved in the tumorigenesis and progression of ovarian cancer. The functional experiments also suggested that Wnt-5a knockdown attenuated the aggressiveness of ovarian carcinoma, including proliferation, anti-apoptosis, migration, and invasion, by modulating phenotype-related molecules such as
Akt,
PI3K,
Bcl-xL,
VEGF,
p70S6k, and
survivin.
Although Strutt
et al. [
27] reported that Wnt-mediated cell migration appeared to require RhoA activation, and Qian
et al. [
28] found that
Wnt-5a activated planar cell polarity (PCP) through a RhoA-dependent process, leading to control of cellular movement, we found that Wnt-5a knockdown significantly reduced both RhoA mRNA and protein expression.
RhoA mRNA expression was positively correlated with that of Wnt-5a mRNA in ovarian carcinoma tissue. These results suggest that there may be a positive correlation between
RhoA and
Wnt-5a expression in ovarian carcinoma. However, further study of validation in
in vitro models may better explain the obtained results.
In conclusion, we demonstrated for the first time that RhoA and Wnt-5a may be associated with ovarian carcinogenesis and apoptosis inhibition; there might be positive correlation between RhoA and Wnt-5a expression. RhoA has potential as a biomarker of tumorigenesis, differentiation, and progression in ovarian carcinoma.
4. Patients and Methods
4.1. Cell Culture and Transfection
The ovarian carcinoma cell lines CAOV3 (serous adenocarcinoma), OVCAR3 (serous cystic adenocarcinoma), SKOV3 (serous papillary cystic adenocarcinoma), HO8910 (serous cystic adenocarcinoma), and ES-2 (clear cell carcinoma) were purchased from ATCC (Manassas, VA, USA). They were maintained in RPMI 1640 (ES-2, HO8910, OVCAR3), DMEM (CAOV3), or McCoy’s 5A (SKOV3) media supplemented with 10% fetal bovine serum (FBS), 100 units/mL penicillin, and 100 μg/mL streptomycin in a humidified atmosphere of 5% CO2 at 37 °C. OVCAR3 cells were transfected with RhoA siRNA or Wnt-5a siRNA (Sigma-Aldrich, St. Louis, MO, USA). The RhoA target sequences were 5′-GUUUAUUAAUCUUAGUGUAdTdT-3′ (sense) and 5′-UACACUAAGAUUAAUAAACdTdT-3′ (antisense). The Wnt-5a target sequences were 5′-CUGACUACUGCGUGCGCAAdTdT-3′ (sense) and 5′-UUGCGCACGCAGUAGUCAGdTdT-3′ (antisense). The NC siRNA sequences were 5′-UUCUCCGAACGUGUCACGUTT-3′ (sense) and 5′-ACGUGACACGUUCGGAGAATT-3′ (antisense). Transfected cells were collected by centrifugation, rinsed with phosphate-buffered saline (PBS), and total proteins were extracted by sonication in radioimmunoprecipitation assay (RIPA) buffer.
4.2. Cell Viability Assay
Cell Counting Kit-8 (CCK-8; Dojindo, Tokyo, Japan) was used to determine the numbers of viable cells. Briefly, 2.0 × 103 cells/well were seeded into 96-well plates and allowed to adhere. The next day, the media was changed to starvation medium (total volume 100 μL/well) for an additional 24 h, and cells were challenged with 200 ng/mL RhoA/Wnt-5a/Mock siRNAs (R & D Systems, Minneapolis, MN, USA). At the appropriate time points, 10 μL CCK-8 solution was added to each well, the plates incubated for 4 h, and the absorbance values measured at 450 nm using a microplate reader.
4.3. Cell Cycle Analysis
After incubation for 48 h at 37 °C in an atmosphere of 5% CO2, cells were trypsinized, harvested, washed twice in PBS, and fixed in 10 mL ice-cold ethanol for at least 2 h. Cells were then washed twice in PBS and incubated with 500 μL RNase (0.25 mg/mL) at 37 °C for 30 min. Cells were pelleted by centrifugation, resuspended in 50 μg/mL Propidium Iodide (PI, KeyGen, Nanjing, China), and incubated at 4 °C in the dark for 30 min. The PI signal was examined using flow cytometry.
4.4. Flow Cytometric Apoptosis Assay
Flow cytometry was performed using cells stained with PI and FITC-labeled annexin V (KeyGen, Nanjing, China) to detect phosphatidylserine externalization as an endpoint indicator of early apoptosis, as described in the manufacturer’s instructions. Briefly, after incubation for 48 h at 37 °C in an atmosphere of 5% CO2, cells were washed twice with cold PBS, resuspended in 1× binding buffer at 1 × 106 cells/mL, and incubated with 200 μL Binding Buffer and 10 μL Annexin V–FITC. Samples were gently vortexed, incubated for 15 min at 25 °C in the dark, 300 μL Binding Buffer and 5 μL PI were added to each tube, and flow cytometry was performed within 1 h.
4.5. Wound Healing Assay
Cells were seeded at 1.0 × 106 cells/well in 6-well culture plates. After cells had grown to confluence, the confluent monolayer in each well was scratched with a pipette tip, washed three times with PBS, and cultured in FBS-free medium. Cells were photographed at 0, 12, 24, 48 h (n = 9); the scratch area was measured using Image J software (National Institutes of Health, Bethesda, MD, USA). The wound healing rate = (Area of original wound − Area of actual wound at different times)/Area of original wound × 100%.
4.6. Cell Invasion Assay
A thin layer of Matrigel (40 μL from 8 mg/mL stock solution; Becton-Dickinson Labware, Bedford, MA, USA) was overlaid on the upper surface of 6.5-mm Transwell chambers (8-μm pore size; BD Bioscience, San Jose, CA, USA). The Matrigel was allowed to solidify by incubating the plates for 4 h at 37 °C. Culture medium was added to the bottom Transwell chamber. Cells were resuspended in serum-free RPMI 1640 (HyClone, Logan, UT, USA) at 2.5 × 105 cells/mL, and 5 × 104 cells were added to the top Transwell chambers. Following 48-h incubation, cells that had not invaded through the Matrigel were removed from the upper surface using cotton swabs. Cells that had invaded through the Matrigel and reached the bottom surface of the filters were fixed in methanol and stained with 0.1% crystal violet. Invasion was quantified by counting the number of cells under a Olympus fluorescence microscope (Tokyo, Japan) equipped with a 16-square reticle. The surface area of this grid was 1 mm2. Five separate fields were counted for each filter (n = 3); the total numbers of cells were compared among experimental groups using the t-test with the assumption of two-tail distribution and two samples with equal variance. A difference of p < 0.05 was considered statistically significant.
4.7. Immunofluorescent Staining
Cells were grown on glass coverslips, fixed with PBS containing 4% formaldehyde for 10 min, permeabilized with 0.2% Triton X-100 in PBS for 10 min at room temperature, washed with PBS, and incubated with Alexa Fluor® 568 phalloidin (Invitrogen, Carlsbad, CA, USA) overnight at 4 °C to visualize the lamellipodia. Nuclei were stained with 1 μg/mL diamidino phenylindole (Sigma-Aldrich, St. Louis, MO, USA) for 15 min at 37 °C. Coverslips were mounted using SlowFade® Gold antifade reagent (Invitrogen, Carlsbad, CA, USA) and cells were observed using a laser confocal microscope (Olympus, Tokyo, Japan).
4.8. Ovarian Epithelium and Carcinoma Samples
Between August 2003 and December 2011, normal fallopian tube epithelium, ovarian epithelial benign tumors (serous and mucinous cystadenoma), primary carcinoma specimens, and metastatic omentum were obtained from patients undergoing surgical resection at the Department of Gynecology, The First Affiliated Hospital of China Medical University. The tumor specimens were microscopically confirmed by pathologists. The average age at surgery was 51.6 years (range 20–81 years). Each ovarian carcinoma specimen was evaluated according to the International Federation of Gynecology and Obstetrics (FIGO) staging system (2009). The histological architecture of ovarian carcinoma was defined in terms of World Health Organization classification. Samples were frozen immediately in liquid nitrogen and stored at −80 °C until analysis. None of the patients underwent chemotherapy, radiotherapy, or adjuvant treatment before surgery. Informed consent was obtained from all subjects; the China Medical University Ethics Committee approved the study.
4.9. Real-Time Reverse Transcriptase PCR
Total RNA was extracted from ovarian carcinoma cell lines and ovarian tissues using TRIzol (Takara, Shiga, Japan) according to the manufacturer’s protocol. Total RNA (2 μg) was reverse transcribed to complementary DNA (cDNA) using avian myeloblastosis virus transcriptase and random primers (Takara, Shiga, Japan). The oligonucleotide primers for PCR were based on GenBank sequences (
Table S3). Real-time PCR amplification of the cDNA was performed in 20-μL reactions according to the SYBR Premix Ex Taq™ II kit (Takara, Shiga, Japan); glyceraldehyde-3-phosphate dehydrogenase (
GAPDH) was used as the internal control.
4.10. Western Blotting
Protein was extracted in ice-cold RIPA lysis buffer and its concentration was determined by protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA). Denatured proteins (100 μg) were separated on sodium dodecyl sulfate–polyacrylamide gels, transferred to Hybond membranes (Amersham, Munich, Germany), and blocked overnight in 5% skimmed milk in Tris-buffered saline with Tween 20 (TBST). For immunoblotting, membranes were incubated for 15 min with antibodies against RhoA (Abcam, Cambridge, UK), Wnt-5a, PI3K, Akt, Bcl-xL, survivin, VEGF (Santa Cruz Biotechnology, Santa Cruz, CA, USA), or p70s6k (T421/s424; Cell Signaling Technology, Danvers, MA, USA). Then, the membranes were rinsed with TBST and incubated with anti-mouse, anti-rabbit, or anti-goat IgG antibodies conjugated to horseradish peroxidase (1:1000; Dako, Carpinteria, CA, USA) for 15 min. All incubations were performed in a microwave oven (Oriental Rotor, Tokyo, Japan) with intermittent irradiation. Bands were visualized on X-ray film (Fujifilm, Tokyo, Japan) using ImageQuant LAS 4000 (Fujifilm, Tokyo, Japan) and ECL Plus detection reagents (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Subsequently, the membranes were washed with WB Stripping Solution (pH 2–3; Nacalai Tesque, Tokyo, Japan) for 1 h and reprobed using mouse anti-GAPDH antibody (Sigma-Aldrich, St. Louis, MO, USA) as the internal control.
4.11. Statistical Analysis
Statistical analysis was performed using the Spearman correlation test for ranked data and the Mann–Whitney U test and paired samples t-test to compare the means of different groups. p < 0.05 were considered statistically significant. SPSS 10.0 software (SPSS, Chicago, IL, USA) was used to analyze all data.

{kind=link}
{kind=link}
{kind=link}
{kind=link}



