Cyclic Stretch Induces Inducible Nitric Oxide Synthase and Soluble Guanylate Cyclase in Pulmonary Artery Smooth Muscle Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
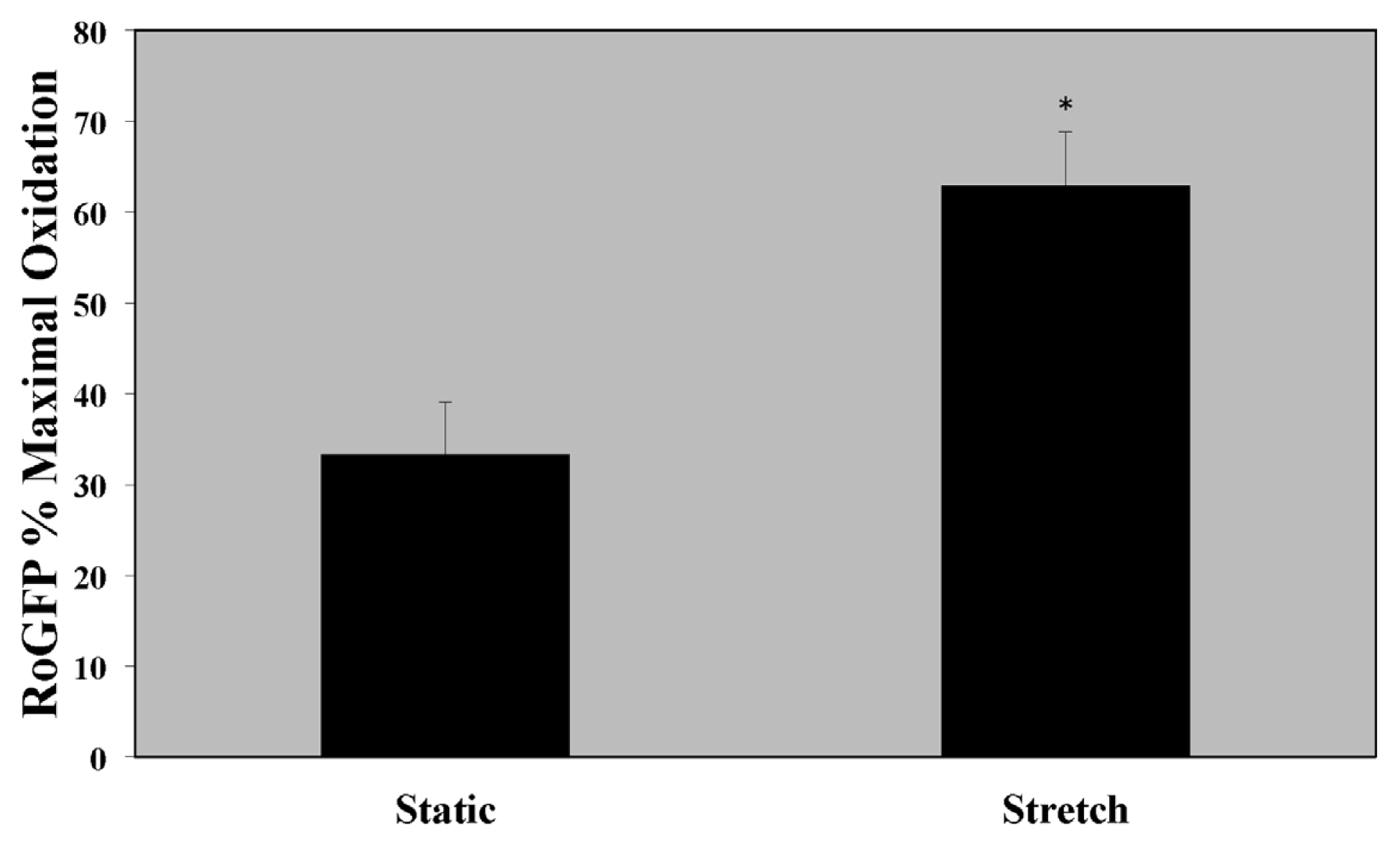
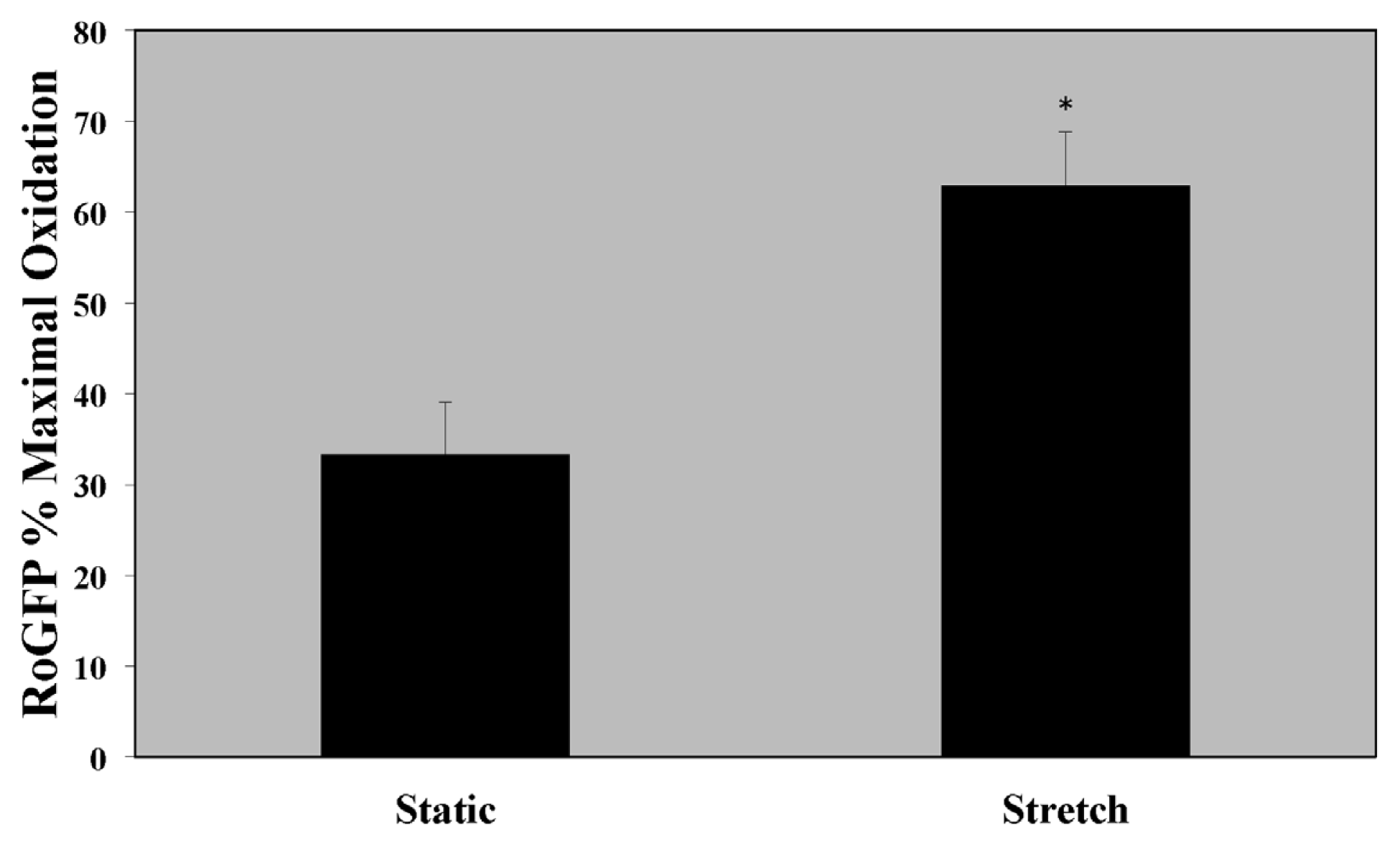
2.1. Cytosolic Oxidant Stress Is Increased in PASMC Exposed to Cyclic Stretch
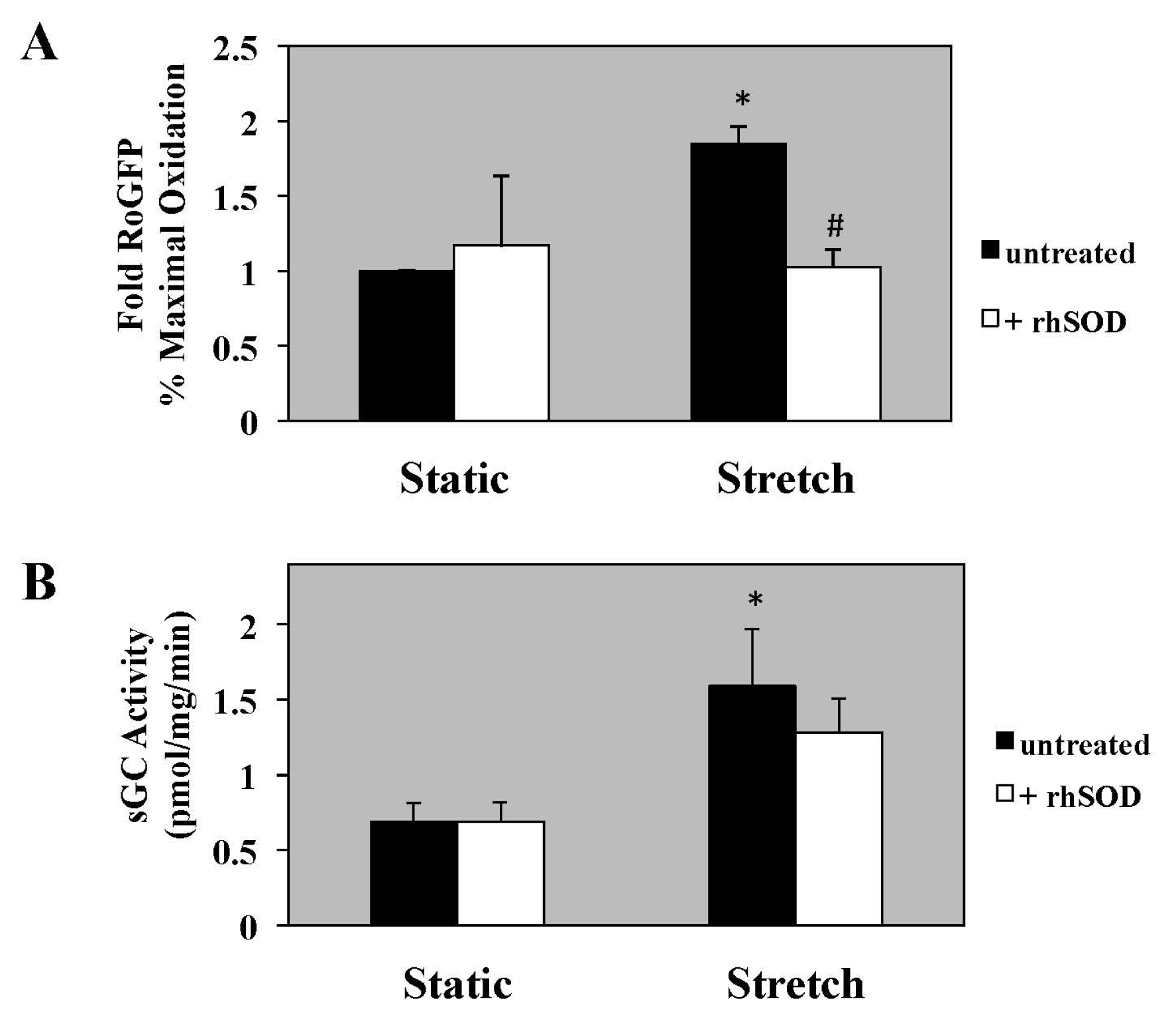
2.2. rhSOD Does not Block Stretch-Induced Increase in sGC Activity
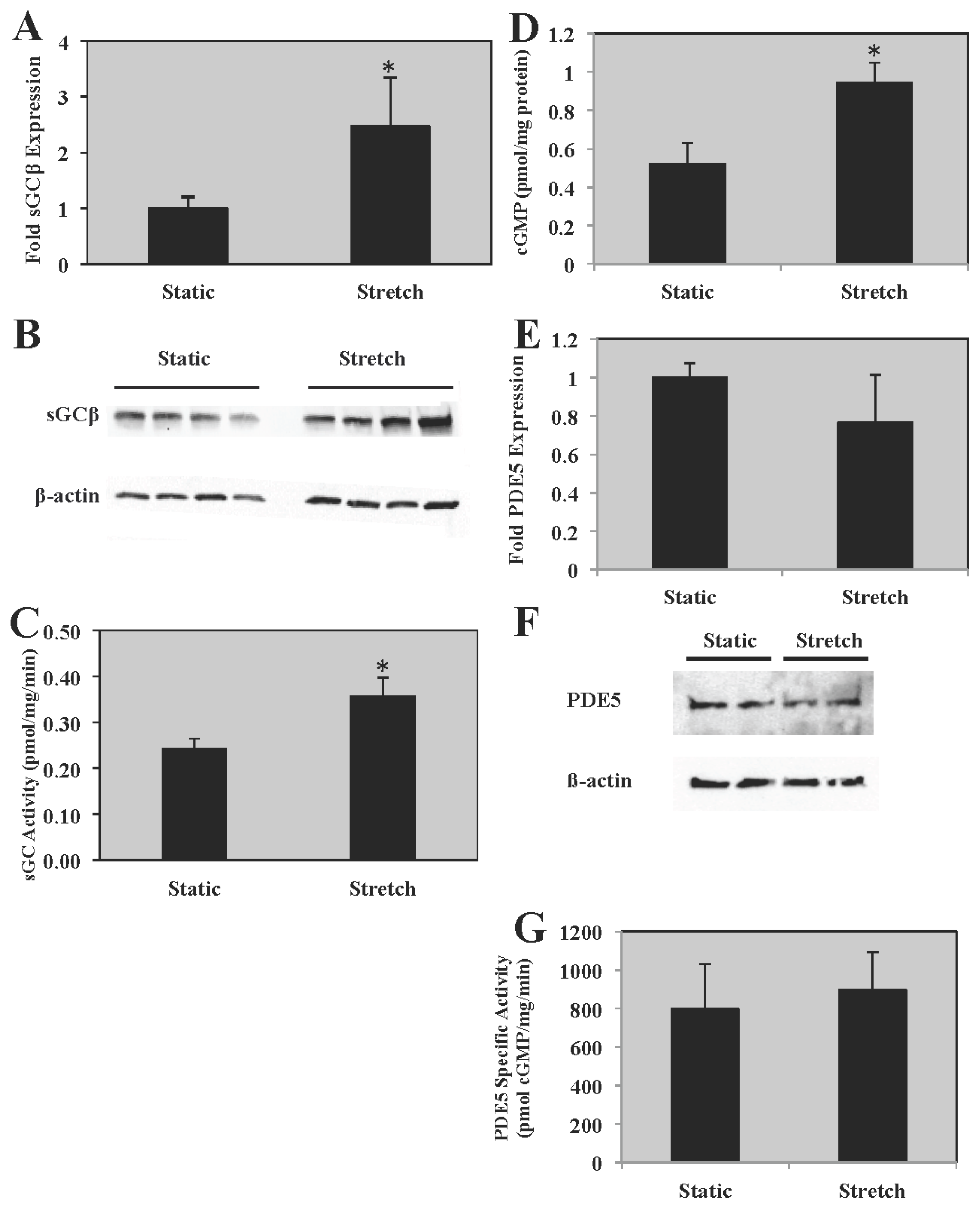
2.3. sGC Expression and Activity Is Increased in PASMC Exposed to Cyclic Stretch
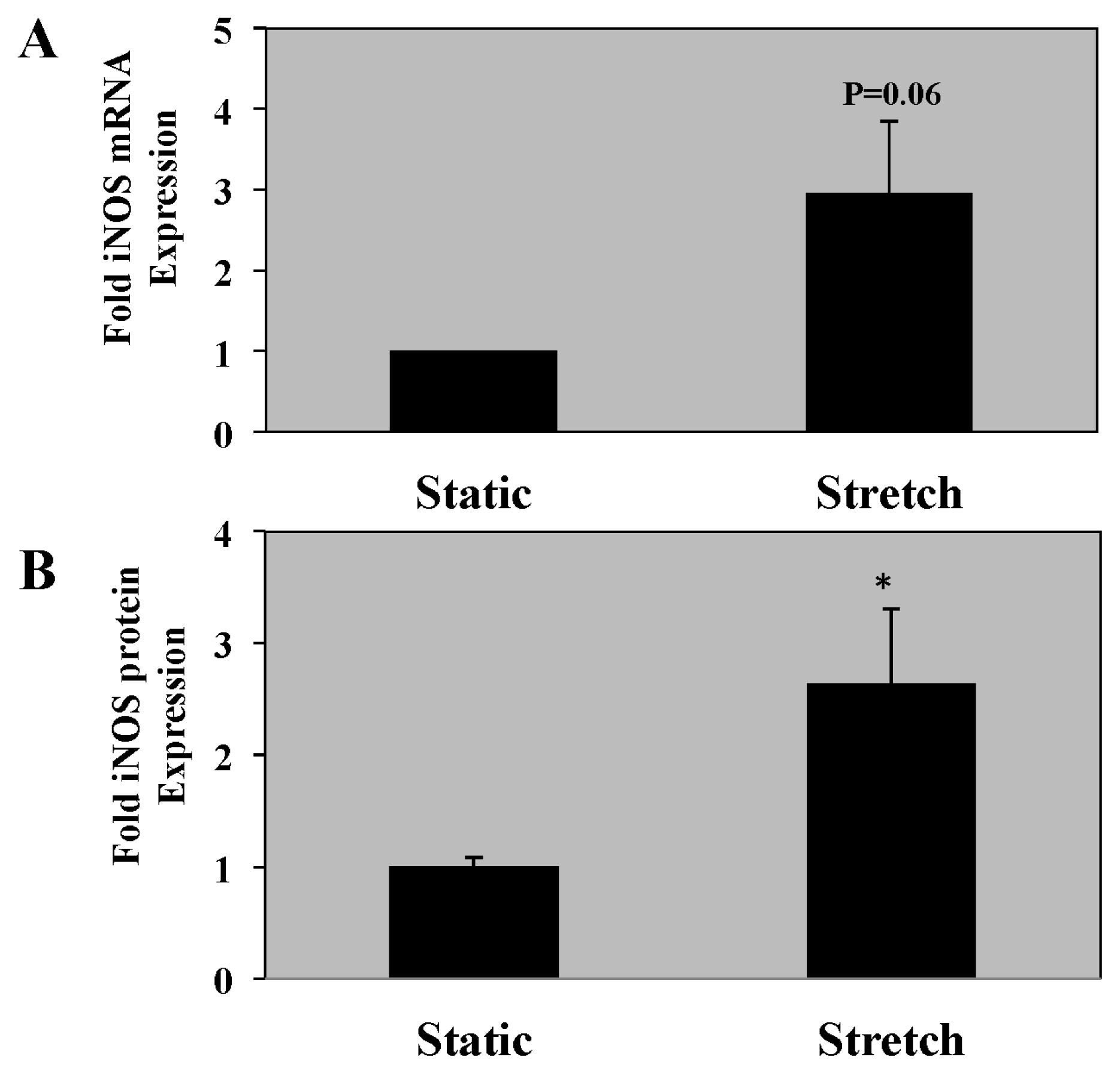
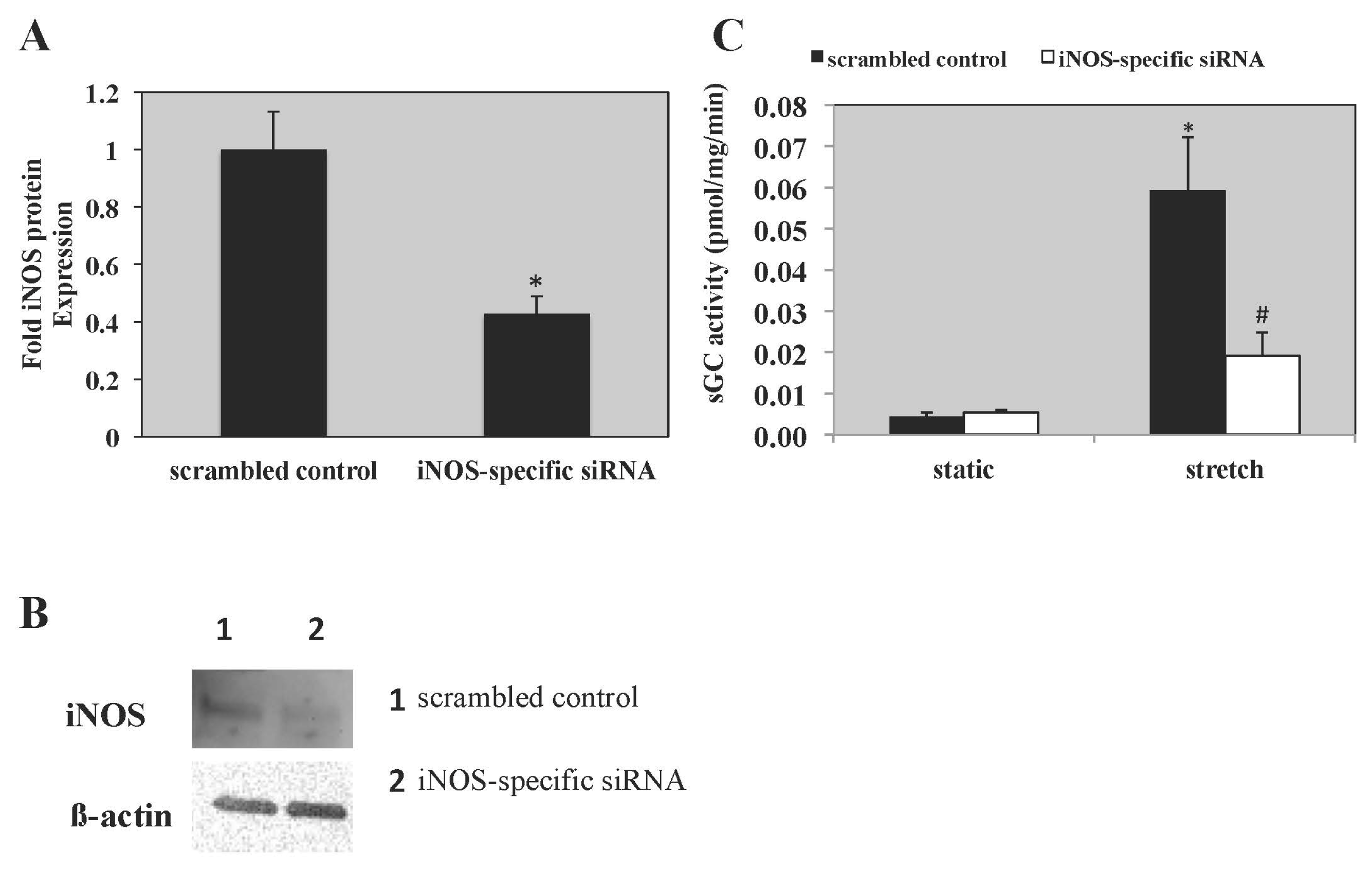
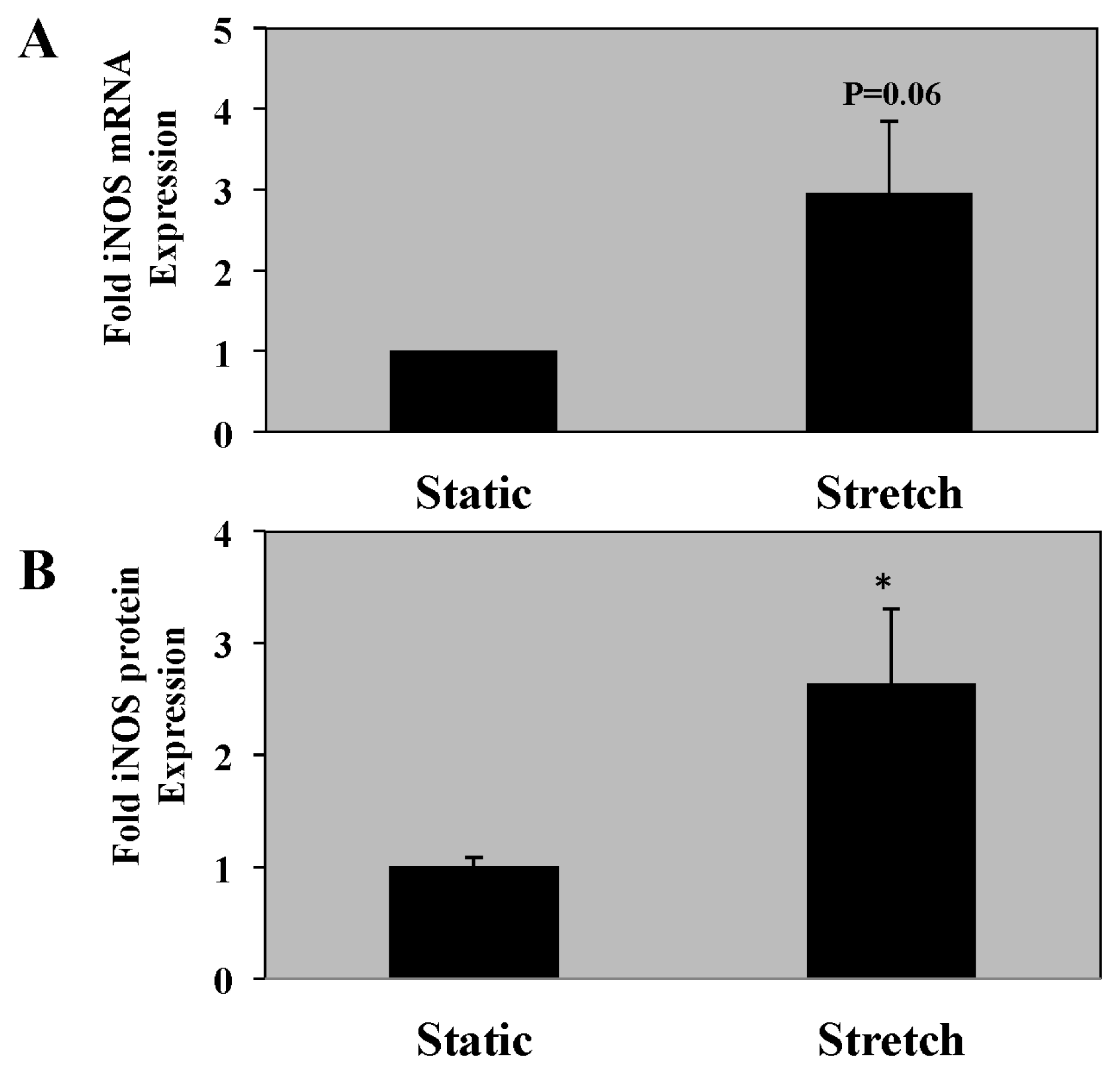
2.4. iNOS mRNA and Protein Expression Is Increased in PASMC Exposed to Cyclic Stretch
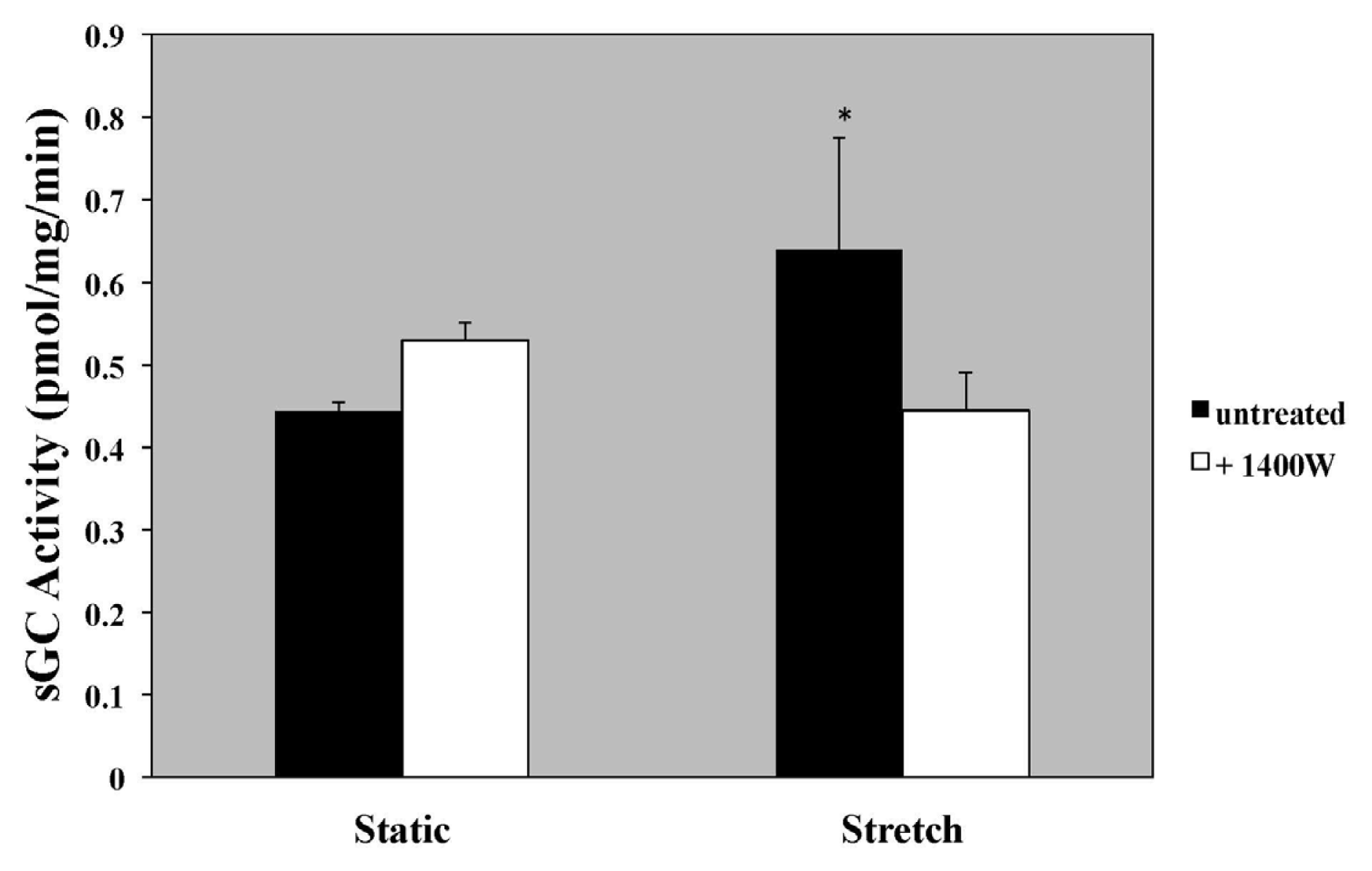
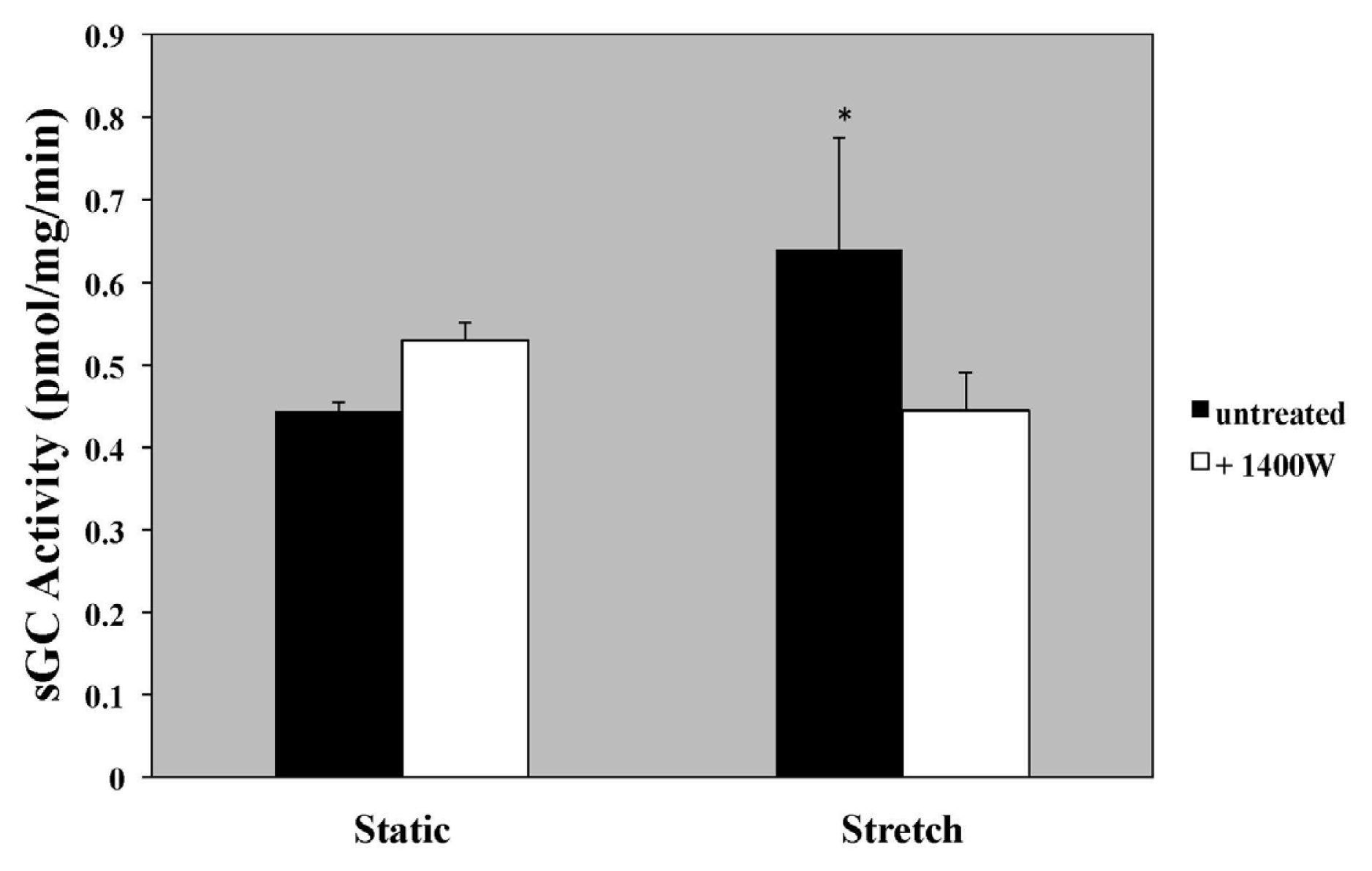
2.5. iNOS Inhibition with 1400 W and iNOS-Specific siRNA Blocks Stretch-Induced sGC Activity
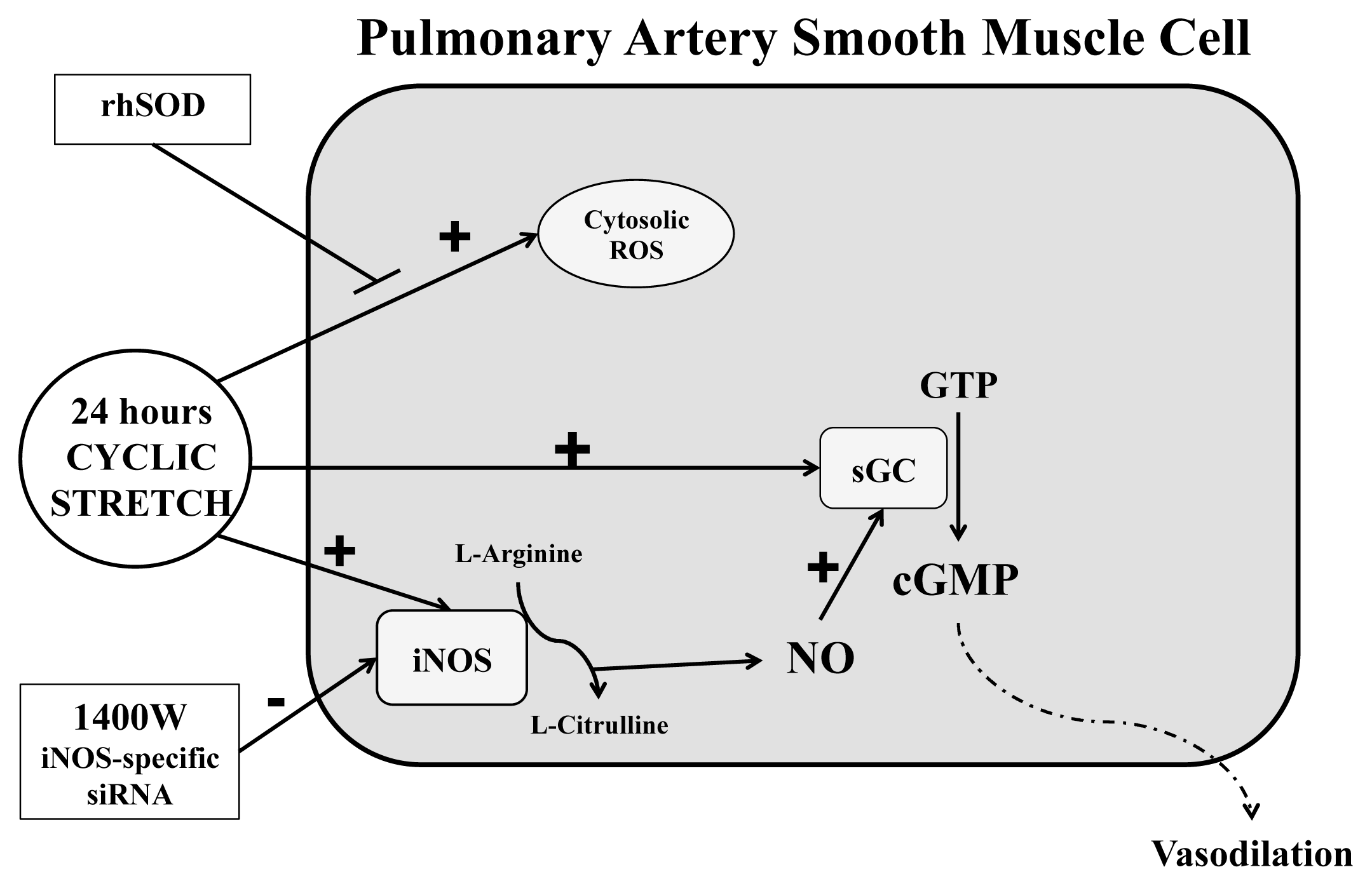
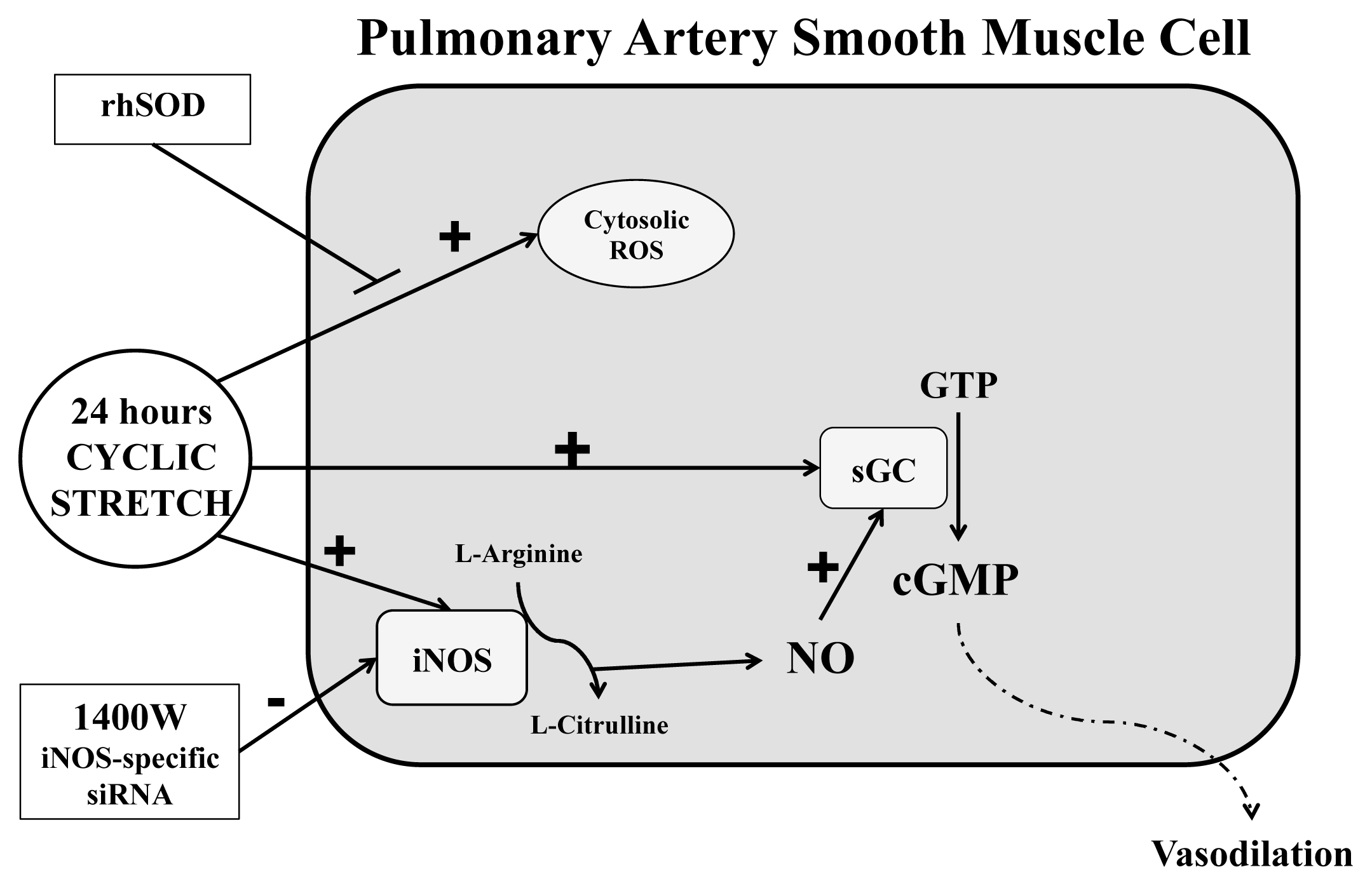
3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. Transfection with iNOS-Specific siRNA
4.3. Detection of Reactive Oxygen Species (ROS)
4.4. Soluble Guanylate Cyclase Activity Assay
4.5. Western Blot Analysis
4.6. cGMP Enzyme Linked Immunoassay
4.7. PDE5 Activity Assay
4.8. iNOS mRNA Expression
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Birukov, K.G. Cyclic stretch, reactive oxygen species and vascular remodeling. Antioxid. Redox Signal 2009, 11, 1651–1667. [Google Scholar]
- Hitomi, H.; Fukui, T.; Moriwaki, K.; Matsubara, K.; Sun, G.P.; Rahman, M.; Nishiyama, A.; Kiyomoto, H.; Kimura, S.; Ohmori, K.; et al. Synergistic effect of mechanical stretch and angiotensin II on superoxide production via NADPH oxidase in vascular smooth muscle cells. J. Hypertens 2006, 24, 1089–1095. [Google Scholar]
- Rairigh, R.L.; Parker, T.A.; Ivy, D.D.; Kinsella, J.P.; Fan, I.D.; Abman, S.H. Role of inducible nitric oxide synthase in the pulmonary vascular response to birth-related stimuli in the ovine fetus. Circ. Res 2001, 88, 721–726. [Google Scholar]
- Rairigh, R.L.; Storme, L.; Parker, T.A.; le Cras, T.D.; Kinsella, J.P.; Jakkula, M.; Abman, S.H. Inducible NO synthase inhibition attenuates shear stress-induced pulmonary vasodilation in the ovine fetus. Am. J. Physiol 1999, 276, L513–L521. [Google Scholar]
- Farrow, K.N.; Lakshminrusimha, S.; Czech, L.; Groh, B.S.; Gugino, S.F.; Davis, J.M.; Russell, J.A.; Steinhorn, R.H. Superoxide dismutase and inhaled nitric oxide normalize phosphodiesterase 5 expression and activity in neonatal lambs with persistent pulmonary hypertension. Am. J. Physiol. Lung Cell. Mol. Physiol 2010, 299, L109–L116. [Google Scholar]
- Steinhorn, R.H.; Russell, J.A.; Morin, F.C., III. Disruption of cyclic GMP production in pulmonary arteries isolated from fetal lambs with pulmonary hypertension. Am. J. Physiol. Heart Circ. Physiol. 1995, 268, H1483–H1489. [Google Scholar]
- Tzao, C.; Nickerson, P.A.; Russell, J.A.; Gugino, S.F.; Steinhorn, R.H. Pulmonary hypertension alters soluble guanylate cyclase activity and expression in pulmonary arteries isolated from fetal lambs. Pediatr Pulmonol 2001, 31, 97–105. [Google Scholar]
- Brennan, L.A.; Steinhorn, R.H.; Wedgwood, S.; Mata-Greenwood, E.; Roark, E.A.; Russell, J.A.; Black, S.M. Increased superoxide generation is associated with pulmonary hypertension in fetal lambs: A role for NADPH oxidase. Circ. Res 2003, 92, 683–691. [Google Scholar]
- Lakshminrusimha, S.; Russell, J.A.; Wedgwood, S.; Gugino, S.F.; Kazzaz, J.A.; Davis, J.M.; Steinhorn, R.H. Superoxide dismutase improves oxygenation and reduces oxidation in neonatal pulmonary hypertension. Am. J. Respir. Crit. Care Med 2006, 174, 1370–1377. [Google Scholar]
- Wedgwood, S.; Lakshminrusimha, S.; Czech, L.; Schumacker, P.T.; Steinhorn, R.H. Increased p22phox/Nox4 Expression is involved in remodeling through hydrogen peroxide signaling in experimental persistent pulmonary hypertension of the newborn. Antioxid. Redox Signal. 2012. [Google Scholar] [CrossRef]
- Wedgwood, S.; Lakshminrusimha, S.; Farrow, K.N.; Czech, L.; Gugino, S.F.; Soares, F.; Russell, J.A.; Steinhorn, R.H. Apocynin improves oxygenation and increases eNOS in persistent pulmonary hypertension of the newborn. Am. J. Physiol. Lung Cell. Mol. Physiol 2012, 302, L616–L626. [Google Scholar]
- Stasch, J.P.; Schmidt, P.M.; Nedvetsky, P.I.; Nedvetskaya, T.Y.; Kumar, A.H.S.; Meurer, S.; Deile, M.; Taye, A.; Knorr, A.; Lapp, H.; et al. Targeting the heme-oxidized nitric oxide receptor for selective vasodilatation of diseased blood vessels. J. Clin. Invest 2006, 116, 2552–2561. [Google Scholar]
- Farrow, K.N.; Lakshminrusimha, S.; Reda, W.J.; Wedgwood, S.; Czech, L.; Gugino, S.F.; Davis, J.M.; Russell, J.A.; Steinhorn, R.H. Superoxide dismutase restores eNOS expression and function in resistance pulmonary arteries from neonatal lambs with persistent pulmonary hypertension. Am. J. Physiol. Lung Cell. Mol. Physiol 2008, 295, L979–L987. [Google Scholar]
- Steinhorn, R.H.; Farrow, K.N. Pulmonary Hypertension in the Neonate. NeoReviews 2007, 8, e14–e21. [Google Scholar]
- Mohammed, K.A.; Nasreen, N.; Tepper, R.S.; Antony, V.B. Cyclic stretch induces PlGF expression in bronchial airway epithelial cells via nitric oxide release. Am. J. Physiol. Lung Cell. Mol. Physiol 2007, 292, L559–L566. [Google Scholar]
- Morisugi, T.; Tanaka, Y.; Kawakami, T.; Kirita, T. Mechanical stretch enhances NF-kappaB-dependent gene expression and poly(ADP-ribose) synthesis in synovial cells. J. Biochem 2010, 147, 633–644. [Google Scholar]
- Mata-Greenwood, E.; Jenkins, C.; Farrow, K.N.; Konduri, G.G.; Russell, J.A.; Lakshminrusimha, S.; Black, S.M.; Steinhorn, R.H. eNOS function is developmentally regulated: Uncoupling of eNOS occurs postnatally. Am. J. Physiol. Lung Cell. Mol. Physiol 2006, 290, L232–L241. [Google Scholar]
- Koesling, D.; Russwurm, M.; Mergia, E.; Mullershausen, F.; Friebe, A. Nitric oxide-sensitive guanylyl cyclase: Structure and regulation. Neurochem. Int 2004, 45, 813–819. [Google Scholar]
- Evgenov, O.V.; Pacher, P.; Schmidt, P.M.; Hasko, G.; Schmidt, H.H.; Stasch, J.P. NO-independent stimulators and activators of soluble guanylate cyclase: Discovery and therapeutic potential. Nat. Rev. Drug Discov 2006, 5, 755–768. [Google Scholar]
- Rairigh, R.L.; Le Cras, T.D.; Ivy, D.D.; Kinsella, J.P.; Richter, G.; Horan, M.P.; Fan, I.D.; Abman, S.H. Role of inducible nitric oxide synthase in regulation of pulmonary vascular tone in the late gestation ovine fetus. J. Clin. Invest 1998, 101, 15–21. [Google Scholar]
- Standley, P.R.; Stanley, M.A.; Senechal, P. Activation of mitogenic and antimitogenic pathways in cyclically stretched arterial smooth muscle. Am. J. Physiol. Endocrinol. Metab 2001, 281, E1165–E1171. [Google Scholar]
- Li, D.; Laubach, V.E.; Johns, R.A. Upregulation of lung soluble guanylate cyclase during chronic hypoxia is prevented by deletion of eNOS. Am. J. Physiol. Lung Cell. Mol. Physiol 2001, 281, L369–L376. [Google Scholar]
- Farrow, K.N.; Groh, B.S.; Schumacker, P.T.; Lakshminrusimha, S.; Czech, L.; Gugino, S.F.; Russell, J.A.; Steinhorn, R.H. Hyperoxia increases phosphodiesterase 5 expression and activity in ovine fetal pulmonary artery smooth muscle cells. Circ. Res 2008, 102, 226–233. [Google Scholar]
- Mata-Greenwood, E.; Grobe, A.; Kumar, S.; Noskina, Y.; Black, S.M. Cyclic stretch increases VEGF expression in pulmonary arterial smooth muscle cells via TGF-beta1 and reactive oxygen species: A requirement for NAD(P)H oxidase. Am. J. Physiol. Lung Cell. Mol. Physiol 2005, 289, L288–L298. [Google Scholar]
- Hanson, G.T.; Aggeler, R.; Oglesbee, D.; Cannon, M.; Capaldi, R.A.; Tsien, R.Y.; Remington, S.J. Investigating mitochondrial redox potential with redox-sensitive green fluorescent protein indicators. J. Biol. Chem 2004, 279, 13044–13053. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Bloch, K.D.; Filippov, G.; Sanchez, L.S.; Nakane, M.; de la Monte, S.M. Pulmonary soluble guanylate cyclase, a nitric oxide receptor, is increased during the perinatal period. Am. J. Physiol 1997, 272, L400–L406. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar]



© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shah, M.R.; Wedgwood, S.; Czech, L.; Kim, G.A.; Lakshminrusimha, S.; Schumacker, P.T.; Steinhorn, R.H.; Farrow, K.N. Cyclic Stretch Induces Inducible Nitric Oxide Synthase and Soluble Guanylate Cyclase in Pulmonary Artery Smooth Muscle Cells. Int. J. Mol. Sci. 2013, 14, 4334-4348. https://doi.org/10.3390/ijms14024334
Shah MR, Wedgwood S, Czech L, Kim GA, Lakshminrusimha S, Schumacker PT, Steinhorn RH, Farrow KN. Cyclic Stretch Induces Inducible Nitric Oxide Synthase and Soluble Guanylate Cyclase in Pulmonary Artery Smooth Muscle Cells. International Journal of Molecular Sciences. 2013; 14(2):4334-4348. https://doi.org/10.3390/ijms14024334
Chicago/Turabian StyleShah, Monica R., Stephen Wedgwood, Lyubov Czech, Gina A. Kim, Satyan Lakshminrusimha, Paul T. Schumacker, Robin H. Steinhorn, and Kathryn N. Farrow. 2013. "Cyclic Stretch Induces Inducible Nitric Oxide Synthase and Soluble Guanylate Cyclase in Pulmonary Artery Smooth Muscle Cells" International Journal of Molecular Sciences 14, no. 2: 4334-4348. https://doi.org/10.3390/ijms14024334