Cold Signaling and Cold Response in Plants

1
Faculty of Life and Environmental Sciences, University of Tsukuba, Tsukuba 305-8572, Japan
2
Department of Agriculture, Ryukoku University, Kyoto 610-8577, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2013, 14(3), 5312-5337; https://doi.org/10.3390/ijms14035312
Submission received: 31 January 2013
/
Revised: 26 February 2013
/
Accepted: 26 February 2013
/
Published: 6 March 2013
(This article belongs to the Special Issue Abiotic and Biotic Stress Tolerance Mechanisms in Plants)
Abstract
:Plants are constantly exposed to a variety of environmental stresses. Freezing or extremely low temperature constitutes a key factor influencing plant growth, development and crop productivity. Plants have evolved a mechanism to enhance tolerance to freezing during exposure to periods of low, but non-freezing temperatures. This phenomenon is called cold acclimation. During cold acclimation, plants develop several mechanisms to minimize potential damages caused by low temperature. Cold response is highly complex process that involves an array of physiological and biochemical modifications. Furthermore, alterations of the expression patterns of many genes, proteins and metabolites in response to cold stress have been reported. Recent studies demonstrate that post-transcriptional and post-translational regulations play a role in the regulation of cold signaling. In this review article, recent advances in cold stress signaling and tolerance are highlighted.
1. Introduction
Only one-third of the total land area on Earth is free of ice and 42% of land experiences temperatures below −20 °C [1]. In such areas, plants require specialized mechanisms to survive exposure to low temperature. Cold stress can be classified as chilling (0–15 °C) and freezing (<0 °C) stresses. Generally, plants originating from temperate regions, such as spinach and Arabidopsis, exhibit a variable degree of chilling tolerance and can increase their freezing tolerance during exposure to chilling and non-freezing temperatures. This process is known as cold acclimation [2]. On the other hand, plants of tropical and subtropical origins are sensitive to chilling stress and lack the cold acclimation mechanism.
The molecular basis of cold acclimation and acquired freezing tolerance in plants, mainly Arabidopsis and winter cereals, has been extensively studied. To adapt to cold stress during cold acclimation, gene expression is reprogrammed and the metabolism is also modified [3]. Cold response is a very complex trait involving many different metabolic pathways, gene regulations and cell compartments [4]. This review highlights cold stress signaling, the transcriptional and post-translational regulation of gene expression during cold acclimation, the interrelationship between cold responses and plant hormones and the transgenic approach to confer cold tolerance onto plants.
2. Cold Stress Sensing and Second Messengers
Thus far, no plant sensors for low temperature have been identified. Multiple primary sensors are thought to be involved in stress sensing. Each sensor may perceive a specific aspect of the stress and may be involved in a distinct branch of the cold signaling pathway. Plants may sense low temperature through changes in the physical properties of membranes, because membrane fluidity is reduced during cold stress [5]. In fact, plasma membrane rigidification raised by a membrane rigidifier, dimethyl sulfoxide (DMSO), can induce the expression of COR (cold-responsive) genes, even at normal growth temperatures, whereas the application of a membrane fluidizer, benzyl alcohol, prevents the induction of COR gene expression at low temperatures [5,6].
Within seconds of a cold shock, a transient increase in cytosolic Ca2+ levels is observed [7]. Membrane rigidification induces cytosolic Ca2+ signatures, and the transient increase in Ca2+ regulates COR gene expression. Because COR gene expression was impaired by gadolinium, a mechanosensitive Ca2+ channel blocker, it is suggested that mechanosensitive Ca2+ channels may be involved in the perception of cold-induced membrane rigidification [7]. The Ca2+ signal can be transduced into the nucleus. Nuclear [Ca2+], which is monitored by a chimera protein, formed by the fusion of aequorin to nucleaoplasmin, is also transiently increased after cold shock, and the peak of nuclear [Ca2+] is delayed at 5 to 10 s, compared to the peak of cytosolic [Ca2+] [8]. In animal cells, the increase in nuclear [Ca2+] is caused by nuclear envelope, which is continuous with the endoplasmic reticulum, one of major Ca2+ stores [9]. The increase in nuclear [Ca2+] can be propagated by cytosolic Ca2+ transients via the nuclear pore complexes [9]. Because the architecture of nuclear envelope in plants is similar to that described in animal cells with the presence of numerous nuclear pore complexes [10], the nuclear Ca2+ signal may be initiated from nuclear envelope and propagated by cytosolic Ca2+ transients in plants. Because nuclear Ca2+ signaling is also important to control gene transcription in plants [11], as well as animal cells [9], identification of transporters, which are localized to plasma membrane or membrane of nuclear envelope and are involved in regulation of cold-inducible Ca2+ transients, may elucidate the detail mechanisms how Ca2+ signal regulates cold signaling.
The cold stress-induced Ca2+ signature can be decoded by different pathways. Plants possess groups of Ca2+ sensors, including CaM (calmodulin) and CMLs (CaM-like), CDPKs (Ca2+-dependent protein kinases), CCaMK (Ca2+-and Ca2+/CaM-dependent protein kinase), CAMTA (CaM-binding transcription activator), CBLs (calcineurin B-like proteins) and CIPKs (CBL-interacting protein kinases). Genetic analysis demonstrated that CDPKs work as positive regulators [12], but calmodulin3 is a negative regulator of gene expression and cold tolerance in plants [13]. CBLs relay the Ca2+ signal by interacting with and regulating the family of CIPKs. As the cbl1 mutant exhibits a chilling sensitive phenotype, CBL1 regulates cold response by interacting with CIPK7 [14]. CAMTA3 has been identified as a positive regulator of CBF2/DREB1C expression through binding to a regulatory element (CG-1 element, vCGCGb) in its promoter [15]. The camta2 camta3 double mutant plants are sensitive to freezing temperatures. The expression of CBF3/DREB1A is not regulated by CAMTA, because there is no CG-1 element in its promoter [15].
In addition to the plasma membrane, chloroplast may also play a role in sensing ambient temperature. Under low temperature, an imbalance between the capacity to harvest light energy and the capacity to dissipate this energy through metabolic activity causes excess photosystem II (PSII) excitation pressure, leading to generation of reactive oxygen species (ROS). Detail is described in the section “Chloroplast and cold response”.
The phosphorylation of proteins in response to cold and the suppression of protein phosphatase activity may also provide a means for the plant to sense low temperature. The MPK (mitogen-activated protein kinase) cascade is implicated in the regulation of cold signaling and cold tolerance. Arabidopsis MPK4 and MPK6 are phosphorylated by MKK2 (MAP kinase kinase2) when exposed to cold stress, and constitutively activated MKK2 overexpressing plants exhibit cold tolerance and the up-regulation of CBF/DREB1s [CRT (C-repeat)/DRE (dehydration responsive-element) binding proteins] [16]. The cold activation of SAMK, an alfalfa MPK, requires membrane rigidification, and the activation of SAMK by low temperatures is inhibited by blocking the influx of extracellular Ca2+ and is prevented by an antagonist of CDPKs, suggesting that membrane rigidification, Ca2+ fluxes and CDPKs are required for the activation of MPK cascades in alfalfa [17]. Together with these results, several signaling pathways are triggered to promote the production of COR (cold-responsive) proteins.
3. ICE-CBF/DREB1 Pathway and Cold-Responsive Gene Regulation
3.1. ICEs (Inducer of CBF Expressions) Are Transcription Factors Controlling Cold Signaling through the Regulation of CBF/DREB1s
Among several cold signaling pathways, the CBF/DREB1-dependent cold signaling pathway is the best characterized and the key regulatory pathway [18]. In Arabidopsis, three CBF/DREB1s are involved in the regulation of COR gene expression and cold tolerance [19,20]. The CBF/DREB1 (mainly CBF3/DREB1A) pathway is controlled by a MYC-type transcription factor ICE1 (inducer of CBF expression1) [21].
The Arabidopsis ice1 mutant exhibits both chilling and freezing sensitivity, and the overexpression of ICE1 confers increased freezing tolerance [21]. ICE1 can bind to the MYC recognition cis-elements (CANNTG) in the promoter of CBF3/DREB1A and induce the expression of CBF3/DREB1A and its regulons during cold acclimation (Figure 1) [21]. Approximately 40% of COR genes and 46% of cold-regulated transcription factor genes are regulated by ICE1, suggesting that ICE1 functions as a master regulator controlling CBF3/DREB1A and many other COR genes [22]. The ice1 mutation affects the early cold induction of CBF1/DREB1B and CBF2/DREB1C, but the induction is not reduced at later times [21], suggesting that other ICE1-like proteins mediate the cold induction of CBF1/DREB1B and CBF2/DREB1C. Supporting this idea, the overexpression of ICE2, a homolog of ICE1, enhances the expression of CBF1/DREB1B and freezing tolerance in Arabidopsis[23]. In the promoter of maize, ZmDREB1, the hyperacetylation of histones H3 and H4 and DNA demethylation, occur in the ICE1-binding region after cold treatment, accompanied by chromatin decondensation [24], suggesting that chromatin remodeling is required for the regulation of CBF/DREB1 by ICE1. Because trichostatin A, an inhibitor of HDACs (histone deacetylases) treatment decreases in induction of ZmDREB1 and ZmCOR413, HDACs may selectively activate transcription [24]. ICE1 is regulated by ubiquitylation [25] and sumoylation (Figure 1) [26] (detail is described in the section “Post-translational regulation”). However, the precise mechanisms for the activation of ICE1 and for transducing signals from perception and the second messengers to ICE1 still remain unknown.
ICE1 is functionally conserved in higher plants (Table 1). The overexpression of SlICE1 enhances the chilling tolerance and activates the expression of SlCBF1, SlDRCi7 (dehydrin Ci7 homolog) and SlP5CS (Δ1-pyrroline-5-carboxylase synthase) in tomatoes (Solanum lycopersicum) [27]. Furthermore, the accumulation of antioxidants, several amino acids, amines and sugars is increased in red tomato fruits of SlICE1-overexpressing plants, leading to an increase in antioxidant activity [28]. Wheat (Triticum aestivum) contains two ICE1 homologs, TaICE141 and TaICE187, which regulate the expression of the wheat CBF group IV genes. The overexpression of these genes in Arabidopsis enhances the CBF/DREB1-dependent COR gene expression and cold tolerance [29]. OsICE1 and OsICE2 in rice (Oryza sativa) are induced by cold stress and sequentially upregulate OsDREB1B, OsHsfA3 (rice heat shock factor A3) and OsTPP1 (rice trehalose 6-phosphate phosphatase), suggesting that these transcription factors are involved in the response to cold stress [30]. The jasmonate-induced chilling tolerance of the banana plant (Musa acuminata) is associated with the induced expression of MaMYC2, which interacts with MaICE1 to activate CBF-dependent cold signaling [31]. Because the function of ICE1 in the cold response is conserved, the overexpression of Arabidopsis ICE1 also improves chilling tolerance and enhances the accumulation of soluble sugars and proline in cucumber [32].
3.2. The CBF/DREB1 Responsive Pathway
CBF/DREB1s can bind to CRT/DRE cis-elements, A/GCCGAC, in the promoter of COR genes to regulate expression of COR genes [56], and belong to the ERF/AP2 (ethylene-responsive element binding factor/APETALA2)-type transcription factor family [57]. Genomic analyses have revealed that CBF/DREB1 genes are organized in tandem (CBF1/DREB1B-CBF3/DREB1A-CBF2/DREB1C) on Arabidopsis chromosome IV [58]. CBF1/DREB1B and CBF3/DREB1A are induced at the same time and earlier than CBF2/DREB1C after cold treatment [59]. Transcriptome analyses in CBF/DREB1-overexpressing transgenic plants reveal that approximately 12% of COR genes in Arabidopsis are controlled by the CBF/DREB1s, but no significant target specificity among the three CBF factors is observed [60,61]. Some transcription factors, such as the ERF/AP2 factors, RAP2.1 and RAP2.6 and the C2H2-type zinc finger, STZ/ZAT10, belong to the CBF-regulon [62,63]. Although no functional difference among three CBF/DREB1 genes was found in an overexpression analysis, a genetic analysis with the cbf2 null mutant demonstrates that the freezing tolerance and expression of CBF1/DREB1B and CBF3/DREB1A are increased in the cbf2 mutant, suggesting that CBF2/DREB1C is a negative regulator of both CBF1/DREB1B and CBF3/DREB1A[64]. On the other hand, CBF1/DREB1B and CBF3/DREB1A are not involved in the regulation of other CBF/DREB1s, but positively regulate cold acclimation by activating the same subset of CBF/DREB1-target genes [65]. These data suggest that CBF1/DREB1B and CBF3/DREB1A have different functions than CBF2/DREB1C. And even though CBF1/DREB1B and CBF3/DREB1A control the same subset of genes, they are concertedly required to induce the whole CBF/DREB1-regulon and complete the development of cold acclimation [18,65].
CBF/DREB1 homologs have been identified in different species. The functional characterization of these homologs reveals that some have similar functions as have Arabidopsis CBF/DREB1s. The overexpression of the homologs from several species in transgenic rice, tobacco or Arabidopsis plants enhances the expression of cold-regulated CBF-regulon genes and cold tolerance. Furthermore, growth retardation is observed in these overexpressing plants, as exhibited by Arabidopsis CBF/DREB1-overexpressing plants (detail is described in the section “Cold response and plant hormone”) (Table 1) [45–50]. On the other hand, the ectopic expression of barley or Arabidopsis CBF/DREB1s in different plants, such as the popular potato, wheat and rice, promotes tolerance to low temperatures (Table 1) [36,40–42]. As for ICE1, the function of CBF/DREB1s seems widely conserved in higher plants. However, as it is noted that no significant increase in cold tolerance is observed in some plants when these factors are overexpressed, differences in CBF/DREB1s function could be considered in different species. The overexpression of either tomato SlCBF1 or Arabidopsis AtCBF3 in transgenic tomatoes does not improve cold tolerance, even though these CBFs enhance the freezing tolerance of Arabidopsis[44]. Additionally, the cold tolerance of transgenic rice expressing Arabidopsis AtCBF1 is not significantly different from that of wild-type plants [66]; however, the overexpression of a monocot-derived CBF, barley HvCBF4, in rice results in an increase in the tolerance to low temperatures [36], suggesting that the function of barley CBF/DREB1 may be different from that of Arabidopsis CBF/DREB1.
A QTL (quantitative trait loci) mapping approach has also demonstrated that CBFs play important roles in cold tolerance. Eleven of 20 CBF/DREB genes are found in two tight tandem clusters on the long arm of chromosome 5H in the same region as the Fr-H2 frost tolerance locus in barley [67,68]. In wheat, Triticum monococcum, similar CBF gene clusters are located at the Fr-Am2 frost tolerance QTL [69,70]. FpCBF6 in Festuca pratensis co-localizes with the major frost tolerance/winter survival QTLs QFt5F-2/QWs5F-1[71]. Together, these results indicate that CBF/DREB acts a key regulator of cold tolerance in various plants.
3.3. Cold-Regulated Genes
The expression of COR has been shown to be critical in plants for both chilling tolerance and cold acclimation. COR78/RD29A, COR47, COR15A and COR6.6 in Arabidopsis and other plants encode dehydrins, which is known as group 2 LEA (LEA II) proteins [72] and are induced by cold stress [73]. LEA proteins are thought to be important for membrane stabilization and prevent protein aggregation [74]. The cold-inducible dehydrins ERD10 (early response to dehydration10) and ERD14 function as chaperones and interact with phospholipid vesicles through electrostatic forces [75,76]. Several dehydrins, such as wheat WCOR410, barley DHN5, peach PCA60 and citrus CuCOR19, increase tolerance to cold stresses [77–81]. In addition, HSP (heat shock protein) expression is also induced by cold in plants [82]. These HSPs function in membrane protection, in the refolding of denatured proteins and in preventing protein aggregation [82,83]. Some PR (pathogen-related) proteins, such as PR1, PR2 (β-1,3-glucanase) and PR5 (thaumatin-like proteins), are induced by cold treatment in Arabidopsis[84]. PR10 (Bet v-1 homologues), PR11 (chitinases) and PR14 (lipid transfer proteins) are also cold-inducible in several species [85–92]. The antifreeze activity of β-1,3-glucanase, chitinases and thaumatin-like proteins inhibits the recrystallization of intercellular ice in the apoplastic space and prevents intracellular ice formation, as cell dehydration is promoted by extracellular freezing [93]. In addition to these proteins, many enzymes are involved in the cold response machinery, such as detoxification and antioxidant cascades, photosynthesis, lignin metabolism, secondary metabolism, cell wall polysaccharide remodeling, starch metabolism, sterol biosynthesis and oligosaccharide synthesis (reviewed in [93]).
Many studies demonstrate the success of transgenic approaches in increasing tolerance to low temperatures. The chloroplast GPAT (glycerol-3-phosphate acyltransferase) of squash, Cucurbita maxima, and Arabidopsis is involved in phosphatidyl glycerol fatty acid desaturation and increases the ratio of unsaturated fatty acids in plant cell membranes, leading to enhancement of cold tolerance [94]. The citrus LEA gene, CuCOR19, enhances the cold tolerance of transgenic tobacco [95]. Similarly, the expression of wheat dehydrin WCS19[96], Arabidopsis COR15A[97] and the co-expression of RAB18 and CO47 or LTI29/XERO2 and LTI30/ERD10[98] increase the freezing tolerance of Arabidopsis transgenic plants. The freezing tolerance of transgenic strawberry leaves expressing wheat dehydrin WCO410 is increased [78]. Rice TPP1 (trehalose-6-phosphate phosphatase) overexpression enhances the cold tolerance of rice [99]. Because trehalose is a nonreducing disaccharide that functions as a stress protection metabolite, the accumulation of trehalose enhances cold tolerance, as well as other abiotic stresses [100]. These results indicate that ICE, CBF/DREB1 and COR genes play important roles in the plant response to low temperatures.
4. Post-Transcriptional Regulation
Post-transcriptional mechanisms based on alternative splicing, pre-mRNA processing, RNA stability, RNA silencing and export from the nucleus play critical roles in cold acclimation and cold tolerance. Pre-mRNA processing and export are important processes for the regulation of gene expression in eukaryotes [101]. Plants regulate the stress-dependent export of mRNA from the nucleus and the selective translation of stress-associated genes and increase the stability of related transcripts [102]. Because, at low temperatures, misfolded RNA molecules become over-stabilized, RNA binding proteins function as RNA chaperones that help RNA achieve their native conformation. Glycine-rich protein GRP7 plays a role in the export of mRNA from the nucleus to the cytoplasm under cold stress conditions [103]. The RNA helicase LOS4 (low expression of osmotically responsive gene4) is important for nuclear mRNA export, particularly in response to temperature stress [104]. mRNA export is inhibited by the los4-1 mutation, leading to the reduced expression of CBF and sensitivity to chilling stress. A null mutation in the AtNUP160 gene, which encodes a nucleoporin protein involved in mRNA export, causes the decreased induction of CBFs and some CBF targets in response to cold [105]. These results suggest that mRNA export plays an important role in the regulation of CBF expression.
In Arabidopsis, the RNA-seq approach revealed that approximately 42% of genes undergo alternative splicing to produce different proteins [106]. EST/cDNA evidence show about 21% of the expressed genes are alternatively spliced in rice [107]. Although most alternative splicing events have not been characterized in plants, several genes encoding protein kinases, transcription factors and splicing regulators have demonstrated the centrality of alternative splicing in the fine-tuning for abiotic stress responses [108]. The pre-mRNA of serine/arginine-rich proteins functions in the regulation of mRNA splicing and alternative splicing of the protein is observed under cold and heat stresses in Arabidopsis[109]. CCA1 (circadian clock associated1) and LHY (late elongated hypocotyl) are MYB-type transcription factors that are core components of the circadian clock in Arabidopsis and positively affect the expression of CBF pathway genes [110]. The CCA1 transcript is alternatively spliced into two variants: CCA1α, the fully spliced variant, and CCA1β, which retains the fourth intron. CCA1α-CCA1α, LHY-LHY or CCA1α-LHY function in the regulation of the circadian clock; however, CCA1β, which lacks a DNA binding domain, interferes with the dimerization. Low temperatures suppressed CCA1 alternative splicing; thus, CCA1β is reduced. Because the overexpression of CCA1α or CCA1β enhances the freezing tolerance or sensitivity, respectively, the alternative splicing of CCA1 regulated by cold stress contributes to freezing tolerance [111]. LHY is also alternatively spliced and the amount of alternatively spliced transcript harboring the long intron of LHY is more abundant in cold-adapted plants. Temperature-dependent alternative splicing is important for decreasing the LHY transcript abundance upon cooling [112]. The alternative splicing of the Arabidopsis transcription factor IDD14 (indeterminate domain14) produces the IDD14β form, which lacks a DNA-binding domain and is produced predominantly under cold conditions [113]. IDD14β interacts with the functional IDD14α, leading to the reduction of the binding activity of the promoter of the QQS (qua-quine starch) gene, which regulates starch accumulation [113]. SR (serine/arginine-rich) proteins are involved in the regulation of mRNA splicing and are also alternatively spliced under cold and heat stresses in Arabidopsis[109].
Small non-coding RNAs, namely, micro-RNAs (miRNAs) and small interfering RNAs (siRNAs), play roles as repressors of gene expression in animals and plants [114]. Plant miRNAs are approximately 21 nt-long small regulatory RNAs that are derived from the processing of longer primary miRNA transcripts [115,116]. miRNAs recognize their mRNA targets based on imperfect sequence complementation. The RISC (RNA-induced silencing complex)-containing miRNA/siRNA induces post-transcriptional gene silencing by the cleavage of mRNA and translational repression [117]. Many cold-regulated miRNAs have been identified by cloning, bioinformatics and high-throughput sequencing approaches in Arabidopsis[118], Populus[119], Brachypodium[120] and rice [121]. Although several miRNAs show similarity among species (miR397 and miR169 are upregulated), several differences in miRNA regulation have been observed, suggesting specificity among species. So far, little is known about the miRNA target genes, whose expression levels are altered in response to cold. The functional analysis of miR169 for cold response has not been characterized, but it is reported that miR169 is involved in the regulation of nitrogen-starvation responses. The miR169a overexpressing plants accumulate less nitrogen and are more sensitive to nitrogen starvation stresses than wild-type [122]. No phenotypic analysis of miR397 has been performed. Cu/Zn SODs (superoxide dismutases) are targets of miR398, whose expression level is downregulated by cold and oxidative stress in Arabidopsis[123]. The stress-induced reduction in miR398 expression results in the accumulation of Cu/Zn SOD transcripts, suggesting that miR398 plays a role in the regulation of ROS detoxification under abiotic stresses.
5. Post-Translational Regulation
The ubiquitylation of a protein leads to its degradation by the 26S proteasome. The ubiquitin-proteasome pathway plays important roles in many biological functions, including abiotic stress responses [124]. Arabidopsis HOS1 (high expression of osmotically responsive gene1) is an ubiquitin E3 ligase that exerts a negative control on cold response and degrades ICE1 (Figure 1) [25]. The hos1 mutant exhibits upregulation of CBF/DREB1s and several cold-regulated genes with cold treatment [125]. HOS1 is shuttled from the cytoplasm to the nucleus during cold acclimation for the poly-ubiquitylation of ICE1 [126]. The substitution of serine 403 of ICE1 to alanine promotes the stabilization of ICE1. The ubiquitylation of ICE1(S403A) is inhibited, and the overexpression of ICE1(S403A) enhances cold tolerance more than the overexpression of wild-type ICE1 [127]. A recent study has demonstrated that the hos1 mutant also exhibits early flowering only under short day conditions, and this photoperiodic control of flowering is caused by HOS1 negatively regulating the levels of the CO (constans) protein, particularly during the daytime [128]. HOS1 is also subjected to alternative splicing, and the overexpression of HOS1-L delays flowering, compared to that of HOS1-S[129]. On the other hand, HOS1 controls flowering time in response to low ambient temperatures (16 °C) and intermittent cold, most likely through the interaction between HOS1 and FVE or FLK (flowering locus K) [130]. Another study demonstrates that intermittent cold treatment triggers the degradation of CO by HOS1, suggesting that the HOS1-CO module contributes to the fine tuning of photoperiodic flowering under short-term temperature fluctuations [131].
The overexpression of another E3 ligase, AtCHIP (carboxyl terminus of HSC70-interacting protein), causes increased sensitivity to low temperatures compared with the wild-type, suggesting its role as a negative regulator of cold response [132]. In wheat, Triticum durum, the RING-finger E3 ligase TdRF1 (RING-finger protein1) interacts with another RING-finger E3 ligase, WVIP2 (wheat viviparus1 interacting protein2), and both genes are upregulated by cold treatment [133]. The transcription factor, WBLH1 (wheat Bel1-type homeodomain1), is degraded in a TdRF1-dependent manner through the 26S proteasome [133]. The overexpression of TdRF1 increases the tolerance of barley cells to dehydration, suggesting that TdRF1 may function in the protection of plants from drought and freezing conditions.
SUMO (small ubiquitin-related modifier) conjugation, or sumoylation, is one of the post-translational modifications [134,135]. Sumoylation plays an important role in a wide variety of cellular processes, including changes in enzyme activity, the regulation of transcription- and chromatin-related processes, sub-cellular relocalization and protection from the ubiquitin-mediated degradation of regulatory proteins [136]. Similar to the ubiquitylation system, sumoylation is processed through the SUMO-specific E1, E2 and E3 enzymes, and SUMO is covalently conjugated to the target consensus motif ΨKxE/D (Ψ, hydrophobic amino acid; K, SUMO target lysine; x, any amino acid; E/D, glutamic acid/aspartic acid). Both loss- and gain-of-function analyses reveal that sumoylation functions in the regulation of responses to abiotic and biotic stresses [134,135,137], such as the response to nutrient availability [138–140], drought tolerance [141,142], basal thermotolerance [143], salt stress tolerance [144,145], copper tolerance [146] and innate immunity [147,148], as well as the development [149–153] and regulation of ABA signaling [154,155]. As the siz1 mutant, which is impaired in the SUMO E3 ligase [138], exhibits hypersensitivity to chilling and freezing stresses [26], sumoylation contributes to the regulation of cold signaling through the stabilization of ICE1. SUMO conjugation to ICE1 blocks its ubiquitylation, allowing for the activation of CBF3/DREB1A transcription (Figure 1). Recent studies have identified hundreds of potential targets for sumoylation [156–159]. Because several transcription factors and chromatin-modifying factors are potential sumoylated targets, these proteins may be involved in the regulation of cold signaling and tolerance. Some examples are GCN5, a histone acetyltransferase and STA1 (stabilized1). GCN5 is an interacting protein of CBF1 and ADA2 [160]. A transcriptional adaptor and the transcription of COR genes was reduced in the gcn5 mutant [161]. STA1 is a pre-mRNA splicing factor that catalyzes the splicing of COR15A for cold tolerance [162].
6. Cold Response and Plant Hormones
Low temperature affects several aspects of plant adaptation, e.g., freezing tolerance, plant growth, abiotic resistance and senescence [163]. Among phytohormones, ABA, auxin, gibberellic acid (GA), salicylic acid (SA) and ethylene are related to the cold responses positively or negatively. Here, the effects of these phytohormones on the cold response are summarized.
The ABA level increases in response to low temperatures [164,165]. The CBF genes are also induced by exogenous ABA [166]. Among cold responses, ABA is not related to vernalization, as an exogenous ABA treatment is not able to induce the expression of a vernalization marker gene, VIN3, but induces the expression of another cold-regulated gene, ADH1[167]. These observations are consistent with early reports showing that the cold signaling associated with vernalization is unnecessary for ABA [168,169] and indicates that the limited effect of ABA to cold-responsive transcriptional regulation.
Temperature has strong effects on the architecture of some plants. Among the above hormones, auxin and GA are directly related to cell-elongation under warm conditions [170]. On the other hand, in the case of Arabidopsis, cold treatment upregulates the GA 2-oxidase gene, encoding the GA-catabolizing enzyme, and represses the GA 20-oxidase gene, encoding a GA-biosynthesizing enzyme. A wheat variety, reduced height 3 (Rht3), exhibits a dwarf phenotype under 20 °C, but normal growth under 10 °C. Rht3 is defective in GA signaling, indicating the importance of GA signaling for warm temperature-dependent cell growth [171]. Hence, the regulation of the GA level is a key output of temperature signaling for determining plant architecture. Under cold or cool conditions, endogenous GA and auxin levels are thus reduced, leading to dwarf architecture in Rht3 wheat variety. The growth retardation of CBF1/DREB1B-overexpressing plants is caused, in part, by accumulation of DELLA proteins [172]. DELLA is degraded by proteasome and degradation of DELLA is enhanced by GA [173]. Thus, growth inhibition of CBF1/DREB1B-overexpressing plants is suppressed by the application of GA and the introduction of the double DELLA mutation gai-t6 rga-24[172]. This data indicates that CBF/DREB1-dependent signaling pathway regulates plant growth through modulation of DELLA protein accumulation.
Endogenous free SA and glucosyl SA are accumulated during chilling in Arabidopsis shoots, wheat and grape berry [174–176]. And SA treatment enhances the cold tolerance of various spices, such as rice, maize, wheat, potato, etc. [177–180]. However, high concentrations and continual application of SA cause growth retardation. Several Arabidopsis mutants, such as cpr1 (constitutive expresser of pathogenesis-related gene1) and acd6 (accelerated cell death6), in which SA is over-accumulated, exhibit a dwarf phenotype and freezing sensitivity [175,181]. ICE1 is a transcription factor classified as a member of the basic Helix-Loop-Helix type transcription factors that binds the promoter of CBF3/DREB1A and, therefore, is one of the central regulators of cold signaling, as described above [163]. In addition, in the ice1 mutant, SA-inducible genes are upregulated [21]. These results indicate the essential function of ICE1 as a cold- and SA-signaling integrator. CAMTA3/AtSR1, a member of the calmodulin binding transcription activator family, recognizes the promoter of CBF2/DREB1C to positively regulate cold tolerance [15] and the promoter of EDS1 to repress disease resistance [182]. These results suggest that cold signaling and SA signaling are interrelated.
Leaf senescence is a complex and tightly regulated process related to cold stress [183]. CBF2/DREB1C overexpression in Arabidopsis shows a delay of senescence in both natural and artificially induced senescence conditions. In the latter cases, an extended lifespan was observed under the darkness, ethylene or ABA treatments, although the detailed molecular mechanism still remains to be solved [184].
7. Chloroplast and Cold Response
As described above, low temperature affects plant architecture. How cold temperatures are sensed is the great enigma for plant scientists. One hypothesis is that the energy balance in chloroplasts may function in sensing ambient temperature [185,186]. Although enzymatic activities are usually limited and dependent on low temperature, the light harvesting system is not affected by the ambient temperature. At low temperature, an imbalance between the capacity for harvesting light energy and the capacity to consume this energy on metabolic activity occurs in leaves. It causes an excess PSII excitation pressure, leading to either reversible downregulation of PSII through the dissipation of excess absorbed energy or the irreversible inactivation of PSII and damage to the D1 reaction center proteins and subsequent inhibition of the photosynthetic capacity [187]. The process is called photoinhibition. Because of the over-reduction of PSII, this imbalance might generate reactive oxygen species (ROS), which cause destruction of the photosynthetic apparatus and damage of whole cells and which presumably function as the second messenger [188,189]. Because the photosynthetic fixation of CO2 is very limited under low temperatures, photoinhibition occurs even under relatively low irradiance [190]. Thus, tolerance to the cold-induced photoinhibition seems to be a mechanism for cold acclimation and to be closely related to freezing tolerance. Over-reduction of PSII may also act as one of the signals triggering gene expression of cold regulated genes in rye, because several cold-inducible genes are also induced by high irradiation [191]. In addition, cold-tolerant crop species, such as spinach, winter wheat, rye and faba bean, are able to maintain the high CO2 assimilation rate, due to changing RubisCO content and nitrogen efficiency, depending on the ambient temperature, whereas cold-sensitive species, such as cucumber, tobacco and rice, are not [192]. Light strength and ambient temperature may be integrated into the alteration of the electron sink in chloroplasts and initiate a signal transduction pathway [186], explaining the coordination of gene expression between plastids and nuclei, which is necessary for proper adaptation to ambient temperature. Various strategies and mechanisms are suggested for acclimation of the photosynthetic apparatus to low temperatures. One of strategies is an increase in the RuBP-regeneration, enhancing the electron flux through the Calvin cycle and photosynthetic capacity in cereals [193–195]. Another strategy is non-photochemical quenching and thermal deactivation of excess light energy occurring within the PSII reaction center (reaction center quenching) [196,197]. Arabidopsis can modify the state transition, PSII and PSI balance, through the phosphorylation and migration of LHCII as cold responses [198]. Despite much physiological evidence supporting this hypothesis, there is little information at the genetic and/or molecular levels, and the molecular mechanisms remain to be elucidated.
8. Conclusions and Perspective
The complex and interactive relationships among different pathways are involved in the regulation of cold acclimation. Among them, the ICE1-CBF/DREB1-dependent pathway is likely to play a central role in regulation of cold signaling, as described above (Figure 1). ICE1 and CBF/DREB1 are also conserved and function in an important role in the regulation of cold signaling in many plants (Table 1). Furthermore, several post-transcriptional and post-translational modifications have been identified. Ubiquitylation and sumoylation are involved in the control of cold signaling through regulation of ICE1. As alternative splicing of clock genes is a regulator for cold signaling, the alternative splicing mechanism may be important to integrate clock and cold responses. Cold signaling and cold responses seem to be regulated by several factors, such as Ca2+ signaling and chloroplast status, as well as clock genes. Identification of CAMTA is direct evidence to connect between Ca2+ signaling and regulation of CBF/DREB1. Because the Ca2+ concentration is transiently increased by cold shock in the cytosol and nucleus, decoding of this Ca2+ signature is required for transducing cold signaling downstream. CAMTA may be one of the candidates for a decoder. The next big challenge should be identification of cold sensors. As described above, multiple primary sensors are thought to be involved in sensing low temperatures, and each sensor may perceive a specific aspect of the stress. Perception of membrane rigidification and an imbalance between capacity of light-energy harvest and of energy dissipation may be included. The elucidation of the sensory mechanism and signaling mechanism from sensor(s) to cold signaling is an important aim in achieving a complete understanding of the cold signaling mechanism.
Acknowledgements
The laboratories are supported by a grant for Scientific Research on Innovative Areas from the Ministry of Education, Culture, Sports, Science and Technology of the Japanese Government (MEXT) on ‘Environmental Sensing of Plants: Signal Perception, Processing Cellular Responses’, a Grant-in-Aid for Challenging Exploratory Research and a Research Grant from the Inamori Foundation.
References
- Ramankutty, N.; Evan, A.T.; Monfreda, C.; Foley, J.A. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol 1999, 50, 571–599. [Google Scholar]
- Chinnusamy, V.; Zhu, J.K.; Sunkar, R. Gene regulation during cold stress acclimation in plants. Methods Mol. Biol 2010, 639, 39–55. [Google Scholar]
- Hannah, M.A.; Heyer, A.G.; Hincha, D.K. A global survey of gene regulation during cold acclimation in Arabidopsis thaliana. PLoS Genet 2005, 1, e26. [Google Scholar]
- Orvar, B.L.; Sangwan, V.; Omann, F.; Dhindsa, R.S. Early steps in cold sensing by plant cells: The role of actin cytoskeleton and membrane fluidity. Plant J 2000, 23, 785–794. [Google Scholar]
- Sangwan, V.; Foulds, I.; Singh, J.; Dhindsa, R.S. Cold-Activation of Brassica napus BN115 promoter is mediated by structural changes in membranes and cytoskeleton, and requires Ca2+ influx. Plant J 2001, 27, 1–12. [Google Scholar]
- Knight, M.R.; Campbell, A.K.; Smith, S.M.; Trewavas, A.J. Transgenic plant aequorin reports the effects of touch and cold-shock and elicitors on cytoplasmic calcium. Nature 1991, 352, 524–526. [Google Scholar]
- Van der Luit, A.H.; Olivari, C.; Haley, A.; Knight, M.R.; Trewavas, A.J. Distinct calcium signaling pathways regulate calmodulin gene expression in tobacco. Plant Physiol 1999, 121, 705–714. [Google Scholar]
- Mauger, J.P. Role of the nuclear envelope in calcium signalling. Biol. Cell 2012, 104, 70–83. [Google Scholar]
- Xu, X.M.; Meier, I. The nuclear pore comes to the fore. Trends Plant Sci 2008, 13, 20–27. [Google Scholar]
- Mazars, C.; Brière, C.; Bourque, S.; Thuleau, P. Nuclear calcium signaling: An emerging topic in plants. Biochimie 2011, 93, 2068–2074. [Google Scholar]
- Saijo, Y.; Hata, S.; Kyozuka, J.; Shimamoto, K.; Izui, K. Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J 2000, 23, 319–327. [Google Scholar]
- Townley, H.E.; Knight, M.R. Calmodulin as a potential negative regulator of Arabidopsis COR gene expression. Plant Physiol 2002, 128, 1169–1172. [Google Scholar]
- Huang, C.; Ding, S.; Zhang, H.; Du, H.; An, L. CIPK7 is involved in cold response by interacting with CBL1 in Arabidopsis thaliana. Plant Sci 2011, 181, 57–64. [Google Scholar]
- Doherty, C.J.; van Buskirk, H.A.; Myers, S.J.; Thomashow, M.F. Roles for Arabidopsis CAMTA transcription factors in cold-regulated gene expression and freezing tolerance. Plant Cell 2009, 21, 972–984. [Google Scholar]
- Teige, M.; Scheikl, E.; Eulgem, T.; Doczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol. Cell 2004, 15, 141–152. [Google Scholar]
- Sangwan, V.; Orvar, B.L.; Beyerly, J.; Hirt, H.; Dhindsa, R.S. Opposite changes in membrane fluidity mimic cold and heat stress activation of distinct plant MAP kinase pathways. Plant J 2002, 31, 629–638. [Google Scholar]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant Sci 2007, 12, 444–451. [Google Scholar]
- Gilmour, S.J.; Fowler, S.G.; Thomashow, M.F. Arabidopsis transcriptional activators CBF1, CBF2, and CBF3 have matching functional activities. Plant Mol. Biol 2004, 54, 767–781. [Google Scholar]
- Gilmour, S.J.; Sebolt, A.M.; Salazar, M.P.; Everard, J.D.; Thomashow, M.F. Overexpression of the Arabidopsis CBF3 transcriptional activator mimics multiple biochemical changes associated with cold acclimation. Plant Physiol 2000, 124, 1854–1865. [Google Scholar]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.H.; Hong, X.; Agarwal, M.; Zhu, J.K. ICE1: A regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev 2003, 17, 1043–1054. [Google Scholar]
- Lee, B.H.; Henderson, D.A.; Zhu, J.K. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar]
- Fursova, O.V.; Pogorelko, G.V.; Tarasov, V.A. Identification of ICE2, a gene involved in cold acclimation which determines freezing tolerance in Arabidopsis thaliana. Gene 2009, 429, 98–103. [Google Scholar]
- Hu, Y.; Zhang, L.; Zhao, L.; Li, J.; He, S.; Zhou, K.; Yang, F.; Huang, M.; Jiang, L.; Li, L. Trichostatin A selectively suppresses the cold-induced transcription of the ZmDREB1 gene in maize. PLoS One 2011, 6, e22132. [Google Scholar]
- Dong, C.H.; Agarwal, M.; Zhang, Y.; Xie, Q.; Zhu, J.K. The negative regulator of plant cold responses, HOS1, is a RING E3 ligase that mediates the ubiquitination and degradation of ICE1. Proc. Natl. Acad. Sci. USA 2006, 103, 8281–8286. [Google Scholar]
- Miura, K.; Jin, J.B.; Lee, J.; Yoo, C.Y.; Stirm, V.; Miura, T.; Ashworth, E.N.; Bressan, R.A.; Yun, D.J.; Hasegawa, P.M. SIZ1-Mediated sumoylation of ICE1 controls CBF3/DREB1A expression and freezing tolerance in Arabidopsis. Plant Cell 2007, 19, 1403–1414. [Google Scholar]
- Miura, K.; Shiba, H.; Ohta, M.; Kang, S.W.; Sato, A.; Yuasa, T.; Iwaya-Inoue, M.; Kamada, H.; Ezura, H. SlICE1 encoding a MYC-type transcription factor controls cold tolerance in tomato, Solanum lycopersicum. Plant Biotechnol 2012, 29, 253–260. [Google Scholar]
- Miura, K.; Sato, A.; Shiba, H.; Kang, S.W.; Kamada, H.; Ezura, H. Accumulation of antioxidants and antioxidant activity in tomato, Solanum lycopersicum, are enhanced by the transcription factor SlICE1. Plant Biotechnol 2012, 29, 261–269. [Google Scholar]
- Badawi, M.; Reddy, Y.V.; Agharbaoui, Z.; Tominaga, Y.; Danyluk, J.; Sarhan, F.; Houde, M. Structure and functional analysis of wheat ICE (inducer of CBF expression) genes. Plant Cell Physiol 2008, 49, 1237–1249. [Google Scholar]
- Nakamura, J.; Yuasa, T.; Huong, T.T.; Harano, K.; Tanaka, S.; Iwata, T.; Phan, T.; Iwaya-Inoue, M. Rice homologs of inducer of CBF expression (OsICE) are involved in cold acclimation. Plant Biotechnol 2011, 28, 303–309. [Google Scholar]
- Zhao, M.L.; Wang, J.N.; Shan, W.; Fan, J.G.; Kuang, J.F.; Wu, K.Q.; Li, X.P.; Chen, W.X.; He, F.Y.; Chen, J.Y.; et al. Induction of jasmonate signalling regulators MaMYC2s and their physical interactions with MaICE1 in methyl jasmonate-induced chilling tolerance in banana fruit. Plant Cell Environ 2012, 36, 30–51. [Google Scholar]
- Liu, L.; Duan, L.; Zhang, J.; Zhang, Z.; Mi, G.; Ren, H. Cucumber (Cucumis sativus L.) over-expressing cold-induced transcriptome regulator ICE1 exhibits changed morphological characters and enhances chilling tolerance. Sci. Hortic 2010, 124, 29–33. [Google Scholar]
- Jaglo-Ottosen, K.R.; Gilmour, S.J.; Zarka, D.G.; Schabenberger, O.; Thomashow, M.F. Arabidopsis CBF1 overexpression induces COR genes and enhances freezing tolerance. Science 1998, 280, 104–106. [Google Scholar]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol 2006, 47, 141–153. [Google Scholar]
- Oh, S.J.; Kwon, C.W.; Choi, D.W.; Song, S.I.; Kim, J.K. Expression of barley HvCBF4 enhances tolerance to abiotic stress in transgenic rice. Plant Biotechnol. J 2007, 5, 646–656. [Google Scholar]
- Morran, S.; Eini, O.; Pyvovarenko, T.; Parent, B.; Singh, R.; Ismagul, A.; Eliby, S.; Shirley, N.; Langridge, P.; Lopato, S. Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotechnol. J 2011, 9, 230–249. [Google Scholar]
- Jaglo, K.R.; Kleff, S.; Amundsen, K.L.; Zhang, X.; Haake, V.; Zhang, J.Z.; Deits, T.; Thomashow, M.F. Components of the Arabidopsis C-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiol 2001, 127, 910–917. [Google Scholar]
- Owens, C.L.; Thomashow, M.F.; Hancock, J.F.; Iezzoni, A.F. CBF1 orthologs in sour cherry and strawberry and the heterologous expression of CBF1 in strawberry. J. Am. Soc. Hortic. Sci 2002, 127, 489–494. [Google Scholar]
- Behnam, B.; Kikuchi, A.; Celebi-Toprak, F.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Watanabe, K. Arabidopsis rd29A::DREB1A enhances freezing tolerance in transgenic potato. Plant Cell. Rep 2007, 26, 1275–1282. [Google Scholar]
- Benedict, C.; Skinner, J.S.; Meng, R.; Chang, Y.; Bhalerao, R.; Huner, N.P.; Finn, C.E.; Chen, T.H.; Hurry, V. The CBF1-dependent low temperature signalling pathway, regulon and increase in freeze tolerance are conserved in Populus spp. Plant Cell Environ 2006, 29, 1259–1272. [Google Scholar]
- Pellegrineschi, A.; Reynolds, M.; Pacheco, M.; Brito, R.M.; Almeraya, R.; Yamaguchi-Shinozaki, K.; Hoisington, D. Stress-Induced expression in wheat of the Arabidopsis thaliana DREB1A gene delays water stress symptoms under greenhouse conditions. Genome 2004, 47, 493–500. [Google Scholar]
- Kasuga, M.; Miura, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. A combination of the Arabidopsis DREB1A gene and stress-inducible rd29A promoter improved drought- and low-temperature stress tolerance in tobacco by gene transfer. Plant Cell Physiol 2004, 45, 346–350. [Google Scholar]
- Zhang, X.; Fowler, S.G.; Cheng, H.; Lou, Y.; Rhee, S.Y.; Stockinger, E.J.; Thomashow, M.F. Freezing-sensitive tomato has a functional CBF cold response pathway, but a CBF regulon that differs from that of freezing-tolerant Arabidopsis. Plant J 2004, 39, 905–919. [Google Scholar]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J 2003, 33, 751–763. [Google Scholar]
- Qin, F.; Sakuma, Y.; Li, J.; Liu, Q.; Li, Y.Q.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Cloning and functional analysis of a novel DREB1/CBF transcription factor involved in cold-responsive gene expression in Zea mays L. Plant Cell Physiol 2004, 45, 1042–1052. [Google Scholar]
- Siddiqua, M.; Nassuth, A. Vitis CBF1 and Vitis CBF4 differ in their effect on Arabidopsis abiotic stress tolerance, development and gene expression. Plant Cell Environ 2011, 34, 1345–1359. [Google Scholar]
- Skinner, J.S.; von Zitzewitz, J.; Szucs, P.; Marquez-Cedillo, L.; Filichkin, T.; Amundsen, K.; Stockinger, E.J.; Thomashow, M.F.; Chen, T.H.; Hayes, P.M. Structural, functional, and phylogenetic characterization of a large CBF gene family in barley. Plant Mol. Biol 2005, 59, 533–551. [Google Scholar]
- Xiong, Y.; Fei, S.-Z. Functional and phylogenetic analysis of a DREB/CBF-like gene in perennial ryegrass (Lolium perenne L.). Planta 2006, 224, 878–888. [Google Scholar]
- Zhao, H.; Bughrara, S.S. Isolation and characterization of cold-regulated transcriptional activator LpCBF3 gene from perennial ryegrass (Lolium perenne L.). Mol. Genet. Genomics 2008, 279, 585–594. [Google Scholar]
- Hsieh, T.H.; Lee, J.T.; Yang, P.T.; Chiu, L.H.; Charng, Y.Y.; Wang, Y.C.; Chan, M.T. Heterology expression of the Arabidopsis C-repeat/dehydration response element binding factor 1 gene confers elevated tolerance to chilling and oxidative stresses in transgenic tomato. Plant Physiol 2002, 129, 1086–1094. [Google Scholar]
- Yang, W.; Liu, X.D.; Chi, X.J.; Wu, C.A.; Li, Y.Z.; Song, L.L.; Liu, X.M.; Wang, Y.F.; Wang, F.W.; Zhang, C.; Liu, Y.; et al. Dwarf apple MbDREB1 enhances plant tolerance to low temperature, drought, and salt stress via both ABA-dependent and ABA-independent pathways. Planta 2011, 233, 219–229. [Google Scholar]
- Chen, M.; Xu, Z.; Xia, L.; Li, L.; Cheng, X.; Dong, J.; Wang, Q.; Ma, Y. Cold-Induced modulation and functional analyses of the DRE-binding transcription factor gene, GmDREB3, in soybean (Glycine max L.). J. Exp. Bot 2009, 60, 121–135. [Google Scholar]
- Welling, A.; Palva, E.T. Involvement of CBF transcription factors in winter hardiness in birch. Plant Physiol 2008, 147, 1199–1211. [Google Scholar]
- Gutha, L.; Reddy, A. Rice DREB1B promoter shows distinct stress-specific responses, and the overexpression of cDNA in tobacco confers improved abiotic and biotic stress tolerance. Plant Mol. Biol 2008, 68, 533–555. [Google Scholar]
- Maruyama, K.; Sakuma, Y.; Kasuga, M.; Ito, Y.; Seki, M.; Goda, H.; Shimada, Y.; Yoshida, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Identification of cold-inducible downstream genes of the Arabidopsis DREB1A/CBF3 transcriptional factor using two microarray systems. Plant J 2004, 38, 982–993. [Google Scholar]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 86–96. [Google Scholar]
- Medina, J.; Bargues, M.; Terol, J.; Perez-Alonso, M.; Salinas, J. The Arabidopsis CBF gene family is composed of three genes encoding AP2 domain-containing proteins whose expression Is regulated by low temperature but not by abscisic acid or dehydration. Plant Physiol 1999, 119, 463–470. [Google Scholar]
- Medina, J.; Catalá, R.; Salinas, J. The CBFs: Three arabidopsis transcription factors to cold acclimate. Plant Sci 2011, 180, 3–11. [Google Scholar]
- Matsui, A.; Ishida, J.; Morosawa, T.; Okamoto, M.; Kim, J.M.; Kurihara, Y.; Kawashima, M.; Tanaka, M.; To, T.K.; Nakaminami, K.; et al. Arabidopsis tiling array analysis to identify the stress-responsive genes. Methods Mol. Biol 2010, 639, 141–155. [Google Scholar]
- Zeller, G.; Henz, S.R.; Widmer, C.K.; Sachsenberg, T.; Ratsch, G.; Weigel, D.; Laubinger, S. Stress-induced changes in the Arabidopsis thaliana transcriptome analyzed using whole-genome tiling arrays. Plant J 2009, 58, 1068–1082. [Google Scholar]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar]
- Vogel, J.T.; Zarka, D.G.; van Buskirk, H.A.; Fowler, S.G.; Thomashow, M.F. Roles of the CBF2 and ZAT12 transcription factors in configuring the low temperature transcriptome of Arabidopsis. Plant J 2005, 41, 195–211. [Google Scholar]
- Novillo, F.; Alonso, J.M.; Ecker, J.R.; Salinas, J. CBF2/DREB1C is a negative regulator of CBF1/DREB1B and CBF3/DREB1A expression and plays a central role in stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 3985–3990. [Google Scholar]
- Novillo, F.; Medina, J.; Salinas, J. Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon. Proc. Natl. Acad. Sci. USA 2007, 104, 21002–21007. [Google Scholar]
- Lee, S.C.; Huh, K.W.; An, K.; An, G.; Kim, S.R. Ectopic expression of a cold-inducible transcription factor, CBF1/DREB1b, in transgenic rice (Oryza sativa L.). Mol. Cells 2004, 18, 107–114. [Google Scholar]
- Francia, E.; Barabaschi, D.; Tondelli, A.; Laido, G.; Rizza, F.; Stanca, A.M.; Busconi, M.; Fogher, C.; Stockinger, E.J.; Pecchioni, N. Fine mapping of a HvCBF gene cluster at the frost resistance locus Fr-H2 in barley. Theor. Appl. Genet 2007, 115, 1083–1091. [Google Scholar]
- Stockinger, E.J.; Skinner, J.S.; Gardner, K.G.; Francia, E.; Pecchioni, N. Expression levels of barley Cbf genes at the Frost resistance-H2 locus are dependent upon alleles at Fr-H1 and Fr-H2. Plant J 2007, 51, 308–321. [Google Scholar]
- Knox, A.K.; Li, C.; Vagujfalvi, A.; Galiba, G.; Stockinger, E.J.; Dubcovsky, J. Identification of candidate CBF genes for the frost tolerance locus Fr-Am2 in Triticum monococcum. Plant Mol. Biol 2008, 67, 257–270. [Google Scholar]
- Miller, A.K.; Galiba, G.; Dubcovsky, J. A cluster of 11 CBF transcription factors is located at the frost tolerance locus Fr-Am2 in Triticum monococcum. Mol. Genet. Genomics 2006, 275, 193–203. [Google Scholar]
- Alm, V.; Busso, C.S.; Ergon, A.; Rudi, H.; Larsen, A.; Humphreys, M.W.; Rognli, O.A. QTL analyses and comparative genetic mapping of frost tolerance, winter survival and drought tolerance in meadow fescue (Festuca pratensis Huds.). Theor. Appl. Genet 2011, 123, 369–382. [Google Scholar]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant. Physiol. Plant Mol. Biol 1996, 47, 377–403. [Google Scholar]
- Thomashow, M.F. Role of cold-responsive genes in plant freezing tolerance. Plant Physiol 1998, 118, 1–8. [Google Scholar]
- Hundertmark, M.; Hincha, D. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 2008, 9, 118. [Google Scholar]
- Kiyosue, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of two cDNAs (ERD10 and ERD14) corresponding to genes that respond rapidly to dehydration stress in Arabidopsis thaliana. Plant Cell Physiol 1994, 35, 225–231. [Google Scholar]
- Kovacs, D.; Kalmar, E.; Torok, Z.; Tompa, P. Chaperone activity of ERD10 and ERD14, two disordered stress-related plant proteins. Plant Physiol 2008, 147, 381–390. [Google Scholar]
- Danyluk, J.; Perron, A.; Houde, M.; Limin, A.; Fowler, B.; Benhamou, N.; Sarhan, F. Accumulation of an acidic dehydrin in the vicinity of the plasma membrane during cold acclimation of wheat. Plant Cell 1998, 10, 623–638. [Google Scholar]
- Houde, M.; Dallaire, S.; N’Dong, D.; Sarhan, F. Overexpression of the acidic dehydrin WCOR410 improves freezing tolerance in transgenic strawberry leaves. Plant Biotechnol. J 2004, 2, 381–387. [Google Scholar]
- Bravo, L.A.; Gallardo, J.; Navarrete, A.; Olave, N.; Martínez, J.; Alberdi, M.; Close, T.J.; Corcuera, L.J. Cryoprotective activity of a cold-induced dehydrin purified from barley. Physiol. Plant 2003, 118, 262–269. [Google Scholar]
- Wisniewski, M.; Webb, R.; Balsamo, R.; Close, T.J.; Yu, X.-M.; Griffith, M. Purification, immunolocalization, cryoprotective, and antifreeze activity of PCA60: A dehydrin from peach (Prunus persica). Physiol. Plant 1999, 105, 600–608. [Google Scholar]
- Hara, M.; Terashima, S.; Kuboi, T. Characterization and cryoprotective activity of cold-responsive dehydrin from Citrus unshiu. J. Plant Physiol 2001, 158, 1333–1339. [Google Scholar]
- Timperio, A.M.; Egidi, M.G.; Zolla, L. Proteomics applied on plant abiotic stresses: Role of heat shock proteins (HSP). J. Proteomics 2008, 71, 391–411. [Google Scholar]
- Renaut, J.; Hausman, J.-F.; Wisniewski, M.E. Proteomics and low-temperature studies: Bridging the gap between gene expression and metabolism. Physiol. Plant 2006, 126, 97–109. [Google Scholar]
- Seo, P.J.; Lee, A.K.; Xiang, F.; Park, C.M. Molecular and functional profiling of Arabidopsis pathogenesis-related genes: Insights into their roles in salt response of seed germination. Plant Cell Physiol 2008, 49, 334–344. [Google Scholar]
- Zhang, R.; Wang, Y.; Liu, G.; Li, H. Cloning and characterization of a pathogenesis-related gene (ThPR10) from Tamarix hispida. Acta Biol. Crac. Ser. Bot. 2010, 52, 17–25. [Google Scholar]
- Liu, J.-J.; Ekramoddoullah, A.K.M.; Yu, X. Differential expression of multiple PR10 proteins in western white pine following wounding, fungal infection and cold-hardening. Physiol. Plant 2003, 119, 544–553. [Google Scholar]
- Lee, O.R.; Pulla, R.K.; Kim, Y.J.; Balusamy, S.R.; Yang, D.C. Expression and stress tolerance of PR10 genes from Panax ginseng C. A. Meyer. Mol. Biol. Rep 2012, 39, 2365–2374. [Google Scholar]
- Pak, J.-H.; Chung, E.-S.; Shin, S.-H.; Jeon, E.-H.; Kim, M.-J.; Lee, H.-Y.; Jeung, J.-U.; Hyung, N.-I.; Lee, J.-H.; Chung, Y.-S. Enhanced fungal resistance in Arabidopsis expressing wild rice PR-3 (OgChitIVa) encoding chitinase class IV. Plant Biotechnol. Rep 2009, 3, 147–155. [Google Scholar]
- Yeh, S.; Moffatt, B.A.; Griffith, M.; Xiong, F.; Yang, D.S.C.; Wiseman, S.B.; Sarhan, F.; Danyluk, J.; Xue, Y.Q.; Hew, C.L.; et al. Chitinase genes responsive to cold encode antifreeze proteins in winter cereals. Plant Physiol 2000, 124, 1251–1264. [Google Scholar]
- Pihakaski-Maunsbach, K.; Moffatt, B.; Testillano, P.; Risueño, M.; Yeh, S.; Griffith, M.; Maunsbach, A.B. Genes encoding chitinase-antifreeze proteins are regulated by cold and expressed by all cell types in winter rye shoots. Physiol. Plant 2001, 112, 359–371. [Google Scholar]
- Takenaka, Y.; Nakano, S.; Tamoi, M.; Sakuda, S.; Fukamizo, T. Chitinase gene expression in response to environmental stresses in Arabidopsis thaliana: Chitinase inhibitor allosamidin enhances stress tolerance. Biosci. Biotechnol. Biochem 2009, 73, 1066–1071. [Google Scholar]
- Gaudet, D.A.; Laroche, A.; Frick, M.; Huel, R.; Puchalski, B. Cold induced expression of plant defensin and lipid transfer protein transcripts in winter wheat. Physiol. Plant 2003, 117, 195–205. [Google Scholar]
- Janska, A.; Marsik, P.; Zelenkova, S.; Ovesna, J. Cold stress and acclimation—What is important for metabolic adjustment? Plant Biol 2010, 12, 395–405. [Google Scholar]
- Murata, N.; Ishizaki-Nishizawa, O.; Higashi, S.; Hayashi, H.; Tasaka, Y.; Nishida, I. Genetically engineered alteration in the chilling sensitivity of plants. Nature 1992, 356, 710–713. [Google Scholar]
- Hara, M.; Terashima, S.; Fukaya, T.; Kuboi, T. Enhancement of cold tolerance and inhibition of lipid peroxidation by citrus dehydrin in transgenic tobacco. Planta 2003, 217, 290–298. [Google Scholar]
- C, N.D.; Danyluk, J.; Wilson, K.E.; Pocock, T.; Huner, N.P.; Sarhan, F. Cold-regulated cereal chloroplast late embryogenesis abundant-like proteins. Molecular characterization and functional analyses. Plant Physiol 2002, 129, 1368–1381. [Google Scholar]
- Artus, N.N.; Uemura, M.; Steponkus, P.L.; Gilmour, S.J.; Lin, C.; Thomashow, M.F. Constitutive expression of the cold-regulated Arabidopsis thaliana COR15a gene affects both chloroplast and protoplast freezing tolerance. Proc. Natl. Acad. Sci. USA 1996, 93, 13404–13409. [Google Scholar]
- Puhakainen, T.; Hess, M.W.; Makela, P.; Svensson, J.; Heino, P.; Palva, E.T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Mol. Biol 2004, 54, 743–753. [Google Scholar]
- Ge, L.F.; Chao, D.Y.; Shi, M.; Zhu, M.Z.; Gao, J.P.; Lin, H.X. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta 2008, 228, 191–201. [Google Scholar]
- Jang, I.C.; Oh, S.J.; Seo, J.S.; Choi, W.B.; Song, S.I.; Kim, C.H.; Kim, Y.S.; Seo, H.S.; Choi, Y.D.; Nahm, B.H.; et al. Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. Plant Physiol 2003, 131, 516–524. [Google Scholar]
- Han, J.; Xiong, J.; Wang, D.; Fu, X.D. Pre-mRNA splicing: Where and when in the nucleus. Trends Cell Biol 2011, 21, 336–343. [Google Scholar]
- Ambrosone, A.; Costa, A.; Leone, A.; Grillo, S. Beyond transcription: RNA-binding proteins as emerging regulators of plant response to environmental constraints. Plant Sci 2012, 182, 12–18. [Google Scholar]
- Kim, J.S.; Jung, H.J.; Lee, H.J.; Kim, K.A.; Goh, C.H.; Woo, Y.; Oh, S.H.; Han, Y.S.; Kang, H. Glycine-rich RNA-binding protein 7 affects abiotic stress responses by regulating stomata opening and closing in Arabidopsis thaliana. Plant J 2008, 55, 455–466. [Google Scholar]
- Gong, Z.; Dong, C.H.; Lee, H.; Zhu, J.; Xiong, L.; Gong, D.; Stevenson, B.; Zhu, J.K. A DEAD box RNA helicase is essential for mRNA export and important for development and stress responses in Arabidopsis. Plant Cell 2005, 17, 256–267. [Google Scholar]
- Dong, C.H.; Hu, X.; Tang, W.; Zheng, X.; Kim, Y.S.; Lee, B.H.; Zhu, J.K. A putative Arabidopsis nucleoporin, AtNUP160, is critical for RNA export and required for plant tolerance to cold stress. Mol. Cell. Biol 2006, 26, 9533–9543. [Google Scholar]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.K.; Mockler, T.C. Genome-wide mapping of alternative splicing in Arabidopsis thaliana. Genome Res 2010, 20, 45–58. [Google Scholar]
- Wang, B.B.; Brendel, V. Genomewide comparative analysis of alternative splicing in plants. Proc. Natl. Acad. Sci. USA 2006, 103, 7175–7180. [Google Scholar]
- Mastrangelo, A.M.; Marone, D.; Laido, G.; de Leonardis, A.M.; de Vita, P. Alternative splicing: Enhancing ability to cope with stress via transcriptome plasticity. Plant Sci. 2012, 185–186, 40–49. [Google Scholar]
- Palusa, S.G.; Ali, G.S.; Reddy, A.S. Alternative splicing of pre-mRNAs of Arabidopsis serine/arginine-rich proteins: Regulation by hormones and stresses. Plant J 2007, 49, 1091–1107. [Google Scholar]
- Dong, M.A.; Farre, E.M.; Thomashow, M.F. Circadian clock-associated 1 and late elongated hypocotyl regulate expression of the C-repeat binding factor (CBF) pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 7241–7246. [Google Scholar]
- Seo, P.J.; Park, M.J.; Lim, M.H.; Kim, S.G.; Lee, M.; Baldwin, I.T.; Park, C.M. A self-regulatory circuit of circadian clock-associated1 underlies the circadian clock regulation of temperature responses in Arabidopsis. Plant Cell 2012, 24, 2427–2442. [Google Scholar]
- James, A.B.; Syed, N.H.; Bordage, S.; Marshall, J.; Nimmo, G.A.; Jenkins, G.I.; Herzyk, P.; Brown, J.W.; Nimmo, H.G. Alternative splicing mediates responses of the Arabidopsis circadian clock to temperature changes. Plant Cell 2012, 24, 961–981. [Google Scholar]
- Seo, P.J.; Kim, M.J.; Ryu, J.Y.; Jeong, E.Y.; Park, C.M. Two splice variants of the IDD14 transcription factor competitively form nonfunctional heterodimers which may regulate starch metabolism. Nat. Commun 2011, 2, 303. [Google Scholar]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet 2009, 10, 94–108. [Google Scholar]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar]
- Ding, J.; Zhou, S.; Guan, J. Finding microRNA targets in plants: Current status and perspectives. Genomics Proteomics Bioinforma 2012, 10, 264–275. [Google Scholar]
- Sunkar, R.; Li, Y.F.; Jagadeeswaran, G. Functions of microRNAs in plant stress responses. Trends Plant Sci 2012, 17, 196–203. [Google Scholar]
- Zhou, X.; Wang, G.; Sutoh, K.; Zhu, J.K.; Zhang, W. Identification of cold-inducible microRNAs in plants by transcriptome analysis. Biochim. Biophys. Acta 2008, 1779, 780–788. [Google Scholar]
- Chen, L.; Zhang, Y.; Ren, Y.; Xu, J.; Zhang, Z.; Wang, Y. Genome-wide identification of cold-responsive and new microRNAs in Populus tomentosa by high-throughput sequencing. Biochem. Biophys. Res. Commun 2012, 417, 892–896. [Google Scholar]
- Zhang, J.; Xu, Y.; Huan, Q.; Chong, K. Deep sequencing of Brachypodium small RNAs at the global genome level identifies microRNAs involved in cold stress response. BMC Genomics 2009, 10, 449. [Google Scholar]
- Lv, D.K.; Bai, X.; Li, Y.; Ding, X.D.; Ge, Y.; Cai, H.; Ji, W.; Wu, N.; Zhu, Y.M. Profiling of cold-stress-responsive miRNAs in rice by microarrays. Gene 2010, 459, 39–47. [Google Scholar]
- Zhao, M.; Ding, H.; Zhu, J.K.; Zhang, F.; Li, W.X. Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol 2011, 190, 906–915. [Google Scholar]
- Sunkar, R.; Kapoor, A.; Zhu, J.K. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell 2006, 18, 2051–2065. [Google Scholar]
- Sadanandom, A.; Bailey, M.; Ewan, R.; Lee, J.; Nelis, S. The ubiquitin-proteasome system: Central modifier of plant signalling. New Phytol 2012, 196, 13–28. [Google Scholar]
- Ishitani, M.; Xiong, L.; Lee, H.; Stevenson, B.; Zhu, J.K. HOS1, a genetic locus involved in cold-responsive gene expression in Arabidopsis. Plant Cell 1998, 10, 1151–1161. [Google Scholar]
- Lee, H.; Xiong, L.; Gong, Z.; Ishitani, M.; Stevenson, B.; Zhu, J.K. The Arabidopsis HOS1 gene negatively regulates cold signal transduction and encodes a RING finger protein that displays cold-regulated nucleo—Cytoplasmic partitioning. Genes Dev 2001, 15, 912–924. [Google Scholar]
- Miura, K.; Ohta, M.; Nakazawa, M.; Ono, M.; Hasegawa, P.M. ICE1 Ser403 is necessary for protein stabilization and regulation of cold signaling and tolerance. Plant J 2011, 67, 269–279. [Google Scholar] [Green Version]
- Lazaro, A.; Valverde, F.; Pineiro, M.; Jarillo, J.A. The Arabidopsis E3 ubiquitin ligase HOS1 negatively regulates CONSTANS abundance in the photoperiodic control of flowering. Plant Cell 2012, 24, 982–999. [Google Scholar]
- Lee, J.H.; Kim, S.H.; Kim, J.J.; Ahn, J.H. Alternative splicing and expression analysis of high expression of osmotically responsive genes1 (HOS1) in Arabidopsis. BMB Rep 2012, 45, 515–520. [Google Scholar]
- Lee, J.H.; Kim, J.J.; Kim, S.H.; Cho, H.J.; Kim, J.; Ahn, J.H. The E3 ubiquitin ligase HOS1 regulates low ambient temperature-responsive flowering in Arabidopsis thaliana. Plant Cell Physiol 2012, 53, 1802–1814. [Google Scholar]
- Jung, J.H.; Seo, P.J.; Park, C.M. The E3 ubiquitin ligase HOS1 regulates Arabidopsis flowering by mediating CONSTANS degradation under cold stress. J. Biol. Chem 2012, 287, 43277–43287. [Google Scholar]
- Yan, J.; Wang, J.; Li, Q.; Hwang, J.R.; Patterson, C.; Zhang, H. AtCHIP, a U-box-containing E3 ubiquitin ligase, plays a critical role in temperature stress tolerance in Arabidopsis. Plant Physiol 2003, 132, 861–869. [Google Scholar]
- Guerra, D.; Mastrangelo, A.M.; Lopez-Torrejon, G.; Marzin, S.; Schweizer, P.; Stanca, A.M.; del Pozo, J.C.; Cattivelli, L.; Mazzucotelli, E. Identification of a protein network interacting with TdRF1, a wheat RING ubiquitin ligase with a protective role against cellular dehydration. Plant Physiol 2012, 158, 777–789. [Google Scholar]
- Miura, K.; Hasegawa, P.M. Sumoylation and other ubiquitin-like post-translational modifications in plants. Trends Cell Biol 2010, 20, 223–232. [Google Scholar]
- Miura, K.; Jin, J.B.; Hasegawa, P.M. Sumoylation, a post-translational regulatory process in plants. Curr. Opin. Plant Biol 2007, 10, 495–502. [Google Scholar]
- Gareau, J.R.; Lima, C.D. The SUMO pathway: Emerging mechanisms that shape specificity, conjugation and recognition. Nat. Rev. Mol. Cell Biol 2010, 11, 861–871. [Google Scholar]
- Castro, P.H.; Tavares, R.M.; Bejarano, E.R.; Azevedo, H. SUMO, a heavyweight player in plant abiotic stress responses. Cell. Mol. Life Sci 2012, 69, 3269–3283. [Google Scholar]
- Miura, K.; Rus, A.; Sharkhuu, A.; Yokoi, S.; Karthikeyan, A.S.; Raghothama, K.G.; Baek, D.; Koo, Y.D.; Jin, J.B.; Bressan, R.A.; et al. The Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency responses. Proc. Natl. Acad. Sci. USA 2005, 102, 7760–7765. [Google Scholar]
- Miura, K.; Lee, J.; Gong, Q.; Ma, S.; Jin, J.B.; Yoo, C.Y.; Miura, T.; Sato, A.; Bohnert, H.J.; Hasegawa, P.M. SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation. Plant Physiol 2011, 155, 1000–1012. [Google Scholar]
- Park, B.S.; Song, J.T.; Seo, H.S. Arabidopsis nitrate reductase activity is stimulated by the E3 SUMO ligase AtSIZ1. Nat. Commun 2011, 2, 400. [Google Scholar]
- Catala, R.; Ouyang, J.; Abreu, I.A.; Hu, Y.; Seo, H.; Zhang, X.; Chua, N.H. The Arabidopsis E3 SUMO ligase SIZ1 regulates plant growth and drought responses. Plant Cell 2007, 19, 2952–2966. [Google Scholar]
- Miura, K.; Okamoto, H.; Okuma, E.; Shiba, H.; Kamada, H.; Hasegawa, P.M.; Murata, Y. SIZ1 deficiency causes reduced stomatal aperture and enhanced drought tolerance via controlling salicylic acid-induced accumulation of reactive oxygen species in Arabidopsis. Plant J 2012, 73, 91–104. [Google Scholar]
- Yoo, C.Y.; Miura, K.; Jin, J.B.; Lee, J.; Park, H.C.; Salt, D.E.; Yun, D.J.; Bressan, R.A.; Hasegawa, P.M. SIZ1 small ubiquitin-like modifier E3 ligase facilitates basal thermotolerance in Arabidopsis independent of salicylic acid. Plant Physiol 2006, 142, 1548–1558. [Google Scholar]
- Conti, L.; Kioumourtzoglou, D.; O’Donnell, E.; Dominy, P.; Sadanandom, A. OTS1 and OTS2 SUMO proteases link plant development and survival under salt stress. Plant Signal. Behav 2009, 4, 225–227. [Google Scholar]
- Miura, K.; Sato, A.; Ohta, M.; Furukawa, J. Increased tolerance to salt stress in the phosphate-accumulating Arabidopsis mutants siz1 and pho2. Planta 2011, 234, 1191–1199. [Google Scholar]
- Chen, C.C.; Chen, Y.Y.; Tang, I.C.; Liang, H.M.; Lai, C.C.; Chiou, J.M.; Yeh, K.C. Arabidopsis SUMO E3 ligase SIZ1 is involved in excess copper tolerance. Plant Physiol 2011, 156, 2225–2234. [Google Scholar]
- Lee, J.; Nam, J.; Park, H.C.; Na, G.; Miura, K.; Jin, J.B.; Yoo, C.Y.; Baek, D.; Kim, D.H.; Jeong, J.C.; et al. Salicylic acid-mediated innate immunity in Arabidopsis is regulated by SIZ1 SUMO E3 ligase. Plant J 2007, 49, 79–90. [Google Scholar]
- Van den Burg, H.A.; Kini, R.K.; Schuurink, R.C.; Takken, F.L. Arabidopsis small ubiquitin-like modifier paralogs have distinct functions in development and defense. Plant Cell 2010, 22, 1998–2016. [Google Scholar]
- Ishida, T.; Yoshimura, M.; Miura, K.; Sugimoto, K. MMS21/HPY2 and SIZ1, Two Arabidopsis SUMO E3 Ligases, Have Distinct Functions in Development. PLoS One 2012, 7, e46897. [Google Scholar]
- Miura, K.; Lee, J.; Miura, T.; Hasegawa, P.M. SIZ1 controls cell growth and plant development in Arabidopsis through salicylic acid. Plant Cell Phys 2010, 51, 103–113. [Google Scholar]
- Jin, J.B.; Jin, Y.H.; Lee, J.; Miura, K.; Yoo, C.Y.; Kim, W.Y.; van Oosten, M.; Hyun, Y.; Somers, D.E.; Lee, I.; et al. The SUMO E3 ligase, AtSIZ1, regulates flowering by controlling a salicylic acid-mediated floral promotion pathway and through affects on FLC chromatin structure. Plant J 2008, 53, 530–540. [Google Scholar]
- Ling, Y.; Zhang, C.; Chen, T.; Hao, H.; Liu, P.; Bressan, R.A.; Hasegawa, P.M.; Jin, J.B.; Lin, J. Mutation in SUMO E3 ligase, SIZ1, disrupts the mature female gametophyte in Arabidopsis. PLoS One 2012, 7, e29470. [Google Scholar]
- Ishida, T.; Fujiwara, S.; Miura, K.; Stacey, N.; Yoshimura, M.; Schneider, K.; Adachi, S.; Minamisawa, K.; Umeda, M.; Sugimoto, K. SUMO E3 ligase HIGH PLOIDY2 regulates endocycle onset and meristem maintenance in Arabidopsis. Plant Cell 2009, 21, 2284–2297. [Google Scholar]
- Miura, K.; Lee, J.; Jin, J.B.; Yoo, C.Y.; Miura, T.; Hasegawa, P.M. Sumoylation of ABI5 by the Arabidopsis SUMO E3 ligase SIZ1 negatively regulates abscisic acid signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 5418–5423. [Google Scholar]
- Miura, K.; Hasegawa, P.M. Sumoylation and abscisic acid signaling. Plant Signal. Behav 2009, 4, 1176–1178. [Google Scholar]
- Park, H.C.; Choi, W.; Park, H.J.; Cheong, M.S.; Koo, Y.D.; Shin, G.; Chung, W.S.; Kim, W.Y.; Kim, M.G.; Bressan, R.A.; et al. Identification and molecular properties of SUMO-binding proteins in Arabidopsis. Mol. Cells 2011, 32, 143–151. [Google Scholar]
- Miller, M.J.; Barrett-Wilt, G.A.; Hua, Z.; Vierstra, R.D. Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 16512–16517. [Google Scholar]
- Elrouby, N.; Coupland, G. Proteome-Wide screens for small ubiquitin-like modifier (SUMO) substrates identify Arabidopsis proteins implicated in diverse biological processes. Proc. Natl. Acad. Sci. USA 2010, 107, 17415–17420. [Google Scholar]
- Budhiraja, R.; Hermkes, R.; Muller, S.; Schmidt, J.; Colby, T.; Panigrahi, K.; Coupland, G.; Bachmair, A. Substrates related to chromatin and to RNA-dependent processes are modified by Arabidopsis SUMO isoforms that differ in a conserved residue with influence on desumoylation. Plant Physiol 2009, 149, 1529–1540. [Google Scholar]
- Stockinger, E.J.; Mao, Y.; Regier, M.K.; Triezenberg, S.J.; Thomashow, M.F. Transcriptional adaptor and histone acetyltransferase proteins in Arabidopsis and their interactions with CBF1, a transcriptional activator involved in cold-regulated gene expression. Nucleic Acids Res 2001, 29, 1524–1533. [Google Scholar]
- Vlachonasios, K.E.; Thomashow, M.F.; Triezenberg, S.J. Disruption mutations of ADA2b and GCN5 transcriptional adaptor genes dramatically affect Arabidopsis growth, development, and gene expression. Plant Cell 2003, 15, 626–638. [Google Scholar]
- Lee, B.H.; Kapoor, A.; Zhu, J.; Zhu, J.K. STABILIZED1, a stress-upregulated nuclear protein, is required for pre-mRNA splicing, mRNA turnover, and stress tolerance in Arabidopsis. Plant Cell 2006, 18, 1736–1749. [Google Scholar]
- Lissarre, M.; Ohta, M.; Sato, A.; Miura, K. Cold-Responsive gene regulation during cold acclimation in plants. Plant Signal. Behav 2010, 5, 948–952. [Google Scholar]
- Chen, T.H.; Gusta, L.V. Abscisic acid-induced freezing resistance in cultured plant cells. Plant Physiol 1983, 73, 71–75. [Google Scholar]
- Lang, V.; Palva, E.T. The expression of a rab-related gene, rab18, is induced by abscisic acid during the cold acclimation process of Arabidopsis thaliana (L.) Heynh. Plant Mol. Biol 1992, 20, 951–962. [Google Scholar]
- Knight, H.; Zarka, D.G.; Okamoto, H.; Thomashow, M.F.; Knight, M.R. Abscisic acid induces CBF gene transcription and subsequent induction of cold-regulated genes via the CRT promoter element. Plant Physiol 2004, 135, 1710–1717. [Google Scholar]
- Dolferus, R.; Jacobs, M.; Peacock, W.J.; Dennis, E.S. Differential interactions of promoter elements in stress responses of the Arabidopsis Adh gene. Plant Physiol 1994, 105, 1075–1087. [Google Scholar]
- Chandler, J.; Martinez-Zapater, J.M.; Dean, C. Mutations causing defects in the biosynthesis and response to gibberellins, abscisic acid and phytochrome B do not inhibit vernalization in Arabidopsis fca-1. Planta 2000, 210, 677–682. [Google Scholar]
- Liu, J.; Gilmour, S.J.; Thomashow, M.F.; Van Nocker, S. Cold signalling associated with vernalization in Arabidopsis thaliana does not involve CBF1 or abscisic acid. Physiol. Plant 2002, 114, 125–134. [Google Scholar]
- Penfield, S. Temperature perception and signal transduction in plants. New Phytol 2008, 179, 615–628. [Google Scholar]
- Tonkinson, C.L.; Lyndon, R.F.; Arnold, G.M.; Lenton, J.R. The effects of temperature and the Rht3 dwarfing gene on growth, cell extension, and gibberellin content and responsiveness in the wheat leaf. J. Exp. Bot 1997, 48, 963–970. [Google Scholar]
- Achard, P.; Gong, F.; Cheminant, S.; Alioua, M.; Hedden, P.; Genschik, P. The cold-inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. Plant Cell 2008, 20, 2117–2129. [Google Scholar]
- Dill, A.; Thomas, S.G.; Hu, J.; Steber, C.M.; Sun, T.P. The Arabidopsis F-box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation. Plant Cell 2004, 16, 1392–1405. [Google Scholar]
- Kosova, K.; Prasil, I.T.; Vitamvas, P.; Dobrev, P.; Motyka, V.; Flokova, K.; Novak, O.; Tureckova, V.; Rolcik, J.; Pesek, B.; et al. Complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold tolerance, winter Samanta and spring Sandra. J. Plant Physiol 2012, 169, 567–576. [Google Scholar]
- Scott, I.M.; Clarke, S.M.; Wood, J.E.; Mur, L.A. Salicylate accumulation inhibits growth at chilling temperature in Arabidopsis. Plant Physiol 2004, 135, 1040–1049. [Google Scholar]
- Wan, S.-B.; Tian, L.; Tian, R.-R.; Pan, Q.-H.; Zhan, J.-C.; Wen, P.-F.; Chen, J.-Y.; Zhang, P.; Wang, W.; Huang, W.-D. Involvement of phospholipase D in the low temperature acclimation-induced thermotolerance in grape berry. Plant Physiol. Biochem 2009, 47, 504–510. [Google Scholar]
- Hara, M.; Furukawa, J.; Sato, A.; Mizoguchi, T.; Miura, K. Abiotic Stress Responses in Plant; Parvaiz, A., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; Volume Chapter 13, pp. 235–251. [Google Scholar]
- Kang, H.M.; Saltveit, M.E. Chilling tolerance of maize, cucumber and rice seedling leaves and roots are differentially affected by salicylic acid. Physiol. Plant 2002, 115, 571–576. [Google Scholar]
- Tasgin, E.; Atici, O.; Nalbantoglu, B.; Popova, L.P. Effects of salicylic acid and cold treatments on protein levels and on the activities of antioxidant enzymes in the apoplast of winter wheat leaves. Phytochemistry 2006, 67, 710–715. [Google Scholar]
- Mora-Herrera, M.E.; López-Delgado, H.; Castillo-Morales, A.; Foyer, C.H. Salicylic acid and H2O2 function by independent pathways in the induction of freezing tolerance in potato. Physiol. Plant 2005, 125, 430–440. [Google Scholar]
- Miura, K.; Ohta, M. SIZ1, a small ubiquitin-related modifier ligase, controls cold signaling through regulation of salicylic acid accumulation. J. Plant Physiol 2010, 167, 555–560. [Google Scholar]
- Du, L.; Ali, G.S.; Simons, K.A.; Hou, J.; Yang, T.; Reddy, A.S.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar]
- Masclaux-Daubresse, C.; Purdy, S.; Lemaitre, T.; Pourtau, N.; Taconnat, L.; Renou, J.-P.; Wingler, A. Genetic Variation suggests interaction between cold acclimation and metabolic regulation of leaf senescence. Plant Physiol 2007, 143, 434–446. [Google Scholar]
- Sharabi-Schwager, M.; Lers, A.; Samach, A.; Guy, C.L.; Porat, R. Overexpression of the CBF2 transcriptional activator in Arabidopsis delays leaf senescence and extends plant longevity. J. Exp. Bot 2010, 61, 261–273. [Google Scholar]
- Huner, N.P.A.; Öquist, G.; Sarhan, F. Energy balance and acclimation to light and cold. Trends Plant Sci 1998, 3, 224–230. [Google Scholar]
- Ivanov, A.G.; Rosso, D.; Savitch, L.V.; Stachula, P.; Rosembert, M.; Oquist, G.; Hurry, V.; Huner, N.P. Implications of alternative electron sinks in increased resistance of PSII and PSI photochemistry to high light stress in cold-acclimated Arabidopsis thaliana. Photosynth. Res 2012, 113, 191–206. [Google Scholar]
- Oquist, G.; Huner, N.P. Photosynthesis of overwintering evergreen plants. Annu. Rev. Plant Biol 2003, 54, 329–355. [Google Scholar]
- Scheller, H.V.; Haldrup, A. Photoinhibition of photosystem I. Planta 2005, 221, 5–8. [Google Scholar]
- Tyystjarvi, E. Photoinhibition of photosystem II. Int. Rev. Cell Mol. Biol 2013, 300, 243–303. [Google Scholar]
- Takahashi, S.; Murata, N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci 2008, 13, 178–182. [Google Scholar]
- Ndong, C.; Danyluk, J.; Huner, N.P.; Sarhan, F. Survey of gene expression in winter rye during changes in growth temperature, irradiance or excitation pressure. Plant Mol. Biol 2001, 45, 691–703. [Google Scholar]
- Yamori, W.; Noguchi, K.; Hikosaka, K.; Terashima, I. Phenotypic plasticity in photosynthetic temperature acclimation among crop species with different cold tolerances. Plant Physiol 2010, 152, 388–399. [Google Scholar]
- Hurry, V.M.; Malmberg, G.; Gardestrom, P.; Oquist, G. Effects of a short-term shift to low temperature and of long-term cold hardening on photosynthesis and Ribulose-1,5-Bisphosphate carboxylase/oxygenase and sucrose phosphate synthase activity in leaves of winter rye (Secale cereale L.). Plant Physiol 1994, 106, 983–990. [Google Scholar]
- Strand, A.; Hurry, V.; Gustafsson, P.; Gardestrom, P. Development of Arabidopsis thaliana leaves at low temperatures releases the suppression of photosynthesis and photosynthetic gene expression despite the accumulation of soluble carbohydrates. Plant J 1997, 12, 605–614. [Google Scholar]
- Dahal, K.; Kane, K.; Gadapati, W.; Webb, E.; Savitch, L.V.; Singh, J.; Sharma, P.; Sarhan, F.; Longstaffe, F.J.; Grodzinski, B.; et al. The effects of phenotypic plasticity on photosynthetic performance in winter rye, winter wheat and Brassica napus. Physiol. Plant 2012, 144, 169–188. [Google Scholar]
- Ivanov, A.G.; Sane, P.; Hurry, V.; Król, M.; Sveshnikov, D.; Huner, N.P.A.; Öquist, G. Low-temperature modulation of the redox properties of the acceptor side of photosystem II: photoprotection through reaction centre quenching of excess energy. Physiol. Plant 2003, 119, 376–383. [Google Scholar]
- Ivanov, A.G.; Sane, P.V.; Hurry, V.; Oquist, G.; Huner, N.P. Photosystem II reaction centre quenching: Mechanisms and physiological role. Photosynth. Res 2008, 98, 565–574. [Google Scholar]
- Nellaepalli, S.; Kodru, S.; Subramanyam, R. Effect of cold temperature on regulation of state transitions in Arabidopsis thaliana. J. Photochem. Photobiol. B 2012, 112, 23–30. [Google Scholar]
Figure 1.
The cold signaling pathway, which involves ICE1 and CBF3/DREB1A. ICE1 is a MYC-type transcription factor and binds to cis-elements in the promoter of CBF3/DREB1A to induce its expression. CBF3/DREB1A is an AP2-type transcription factor to regulate expression of COR (cold-regulated genes) and cold tolerance. Ubiquitylation of ICE1 is mediated by HOS1, an ubiquitin E3 ligase for proteasome-dependent degradation. SIZ1, a SUMO E3 ligase, mediates sumoylation (SUMO conjugation) of ICE1, probably leading to blocking of ubiquitylation and stabilization of ICE1.
Figure 1.
The cold signaling pathway, which involves ICE1 and CBF3/DREB1A. ICE1 is a MYC-type transcription factor and binds to cis-elements in the promoter of CBF3/DREB1A to induce its expression. CBF3/DREB1A is an AP2-type transcription factor to regulate expression of COR (cold-regulated genes) and cold tolerance. Ubiquitylation of ICE1 is mediated by HOS1, an ubiquitin E3 ligase for proteasome-dependent degradation. SIZ1, a SUMO E3 ligase, mediates sumoylation (SUMO conjugation) of ICE1, probably leading to blocking of ubiquitylation and stabilization of ICE1.
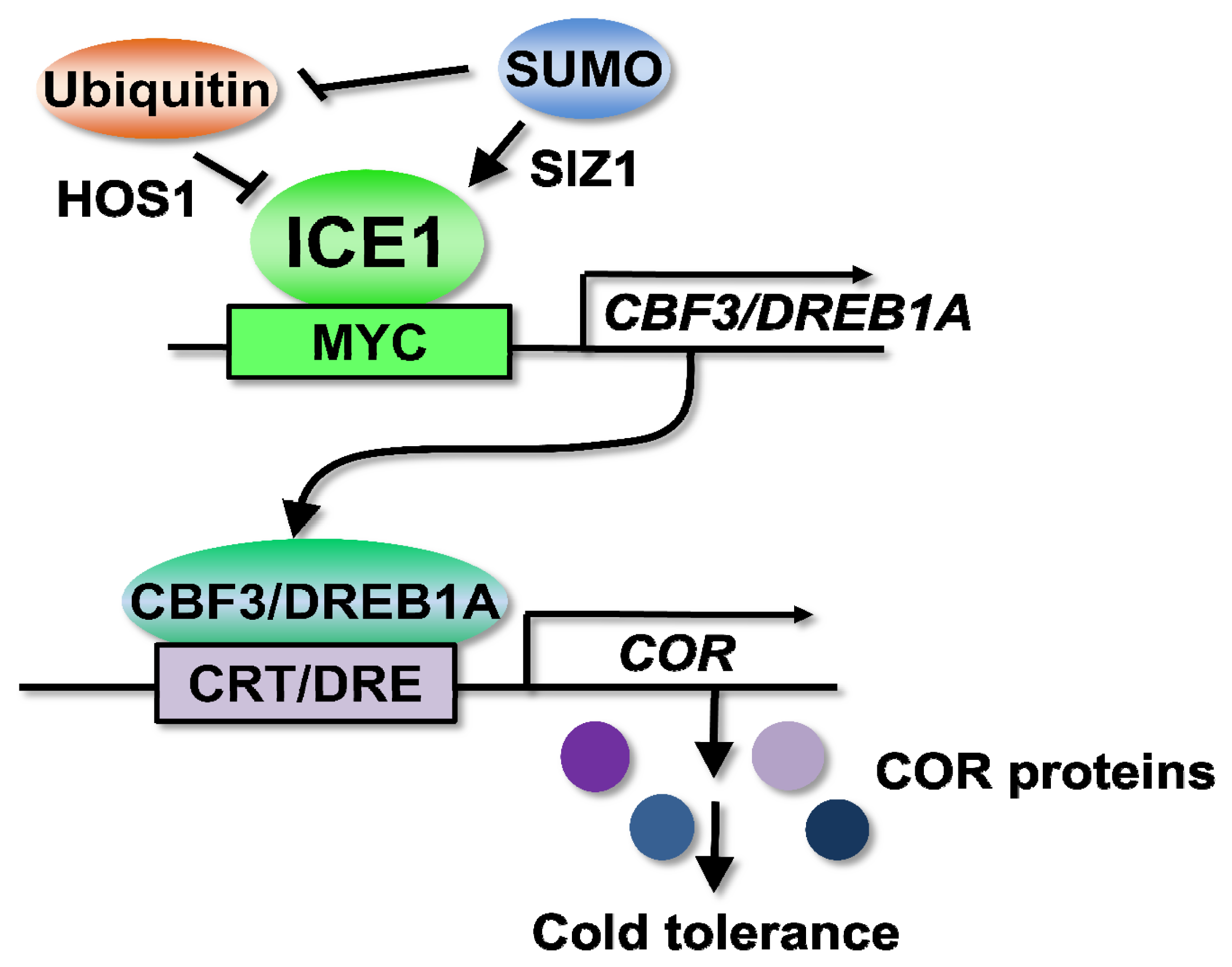

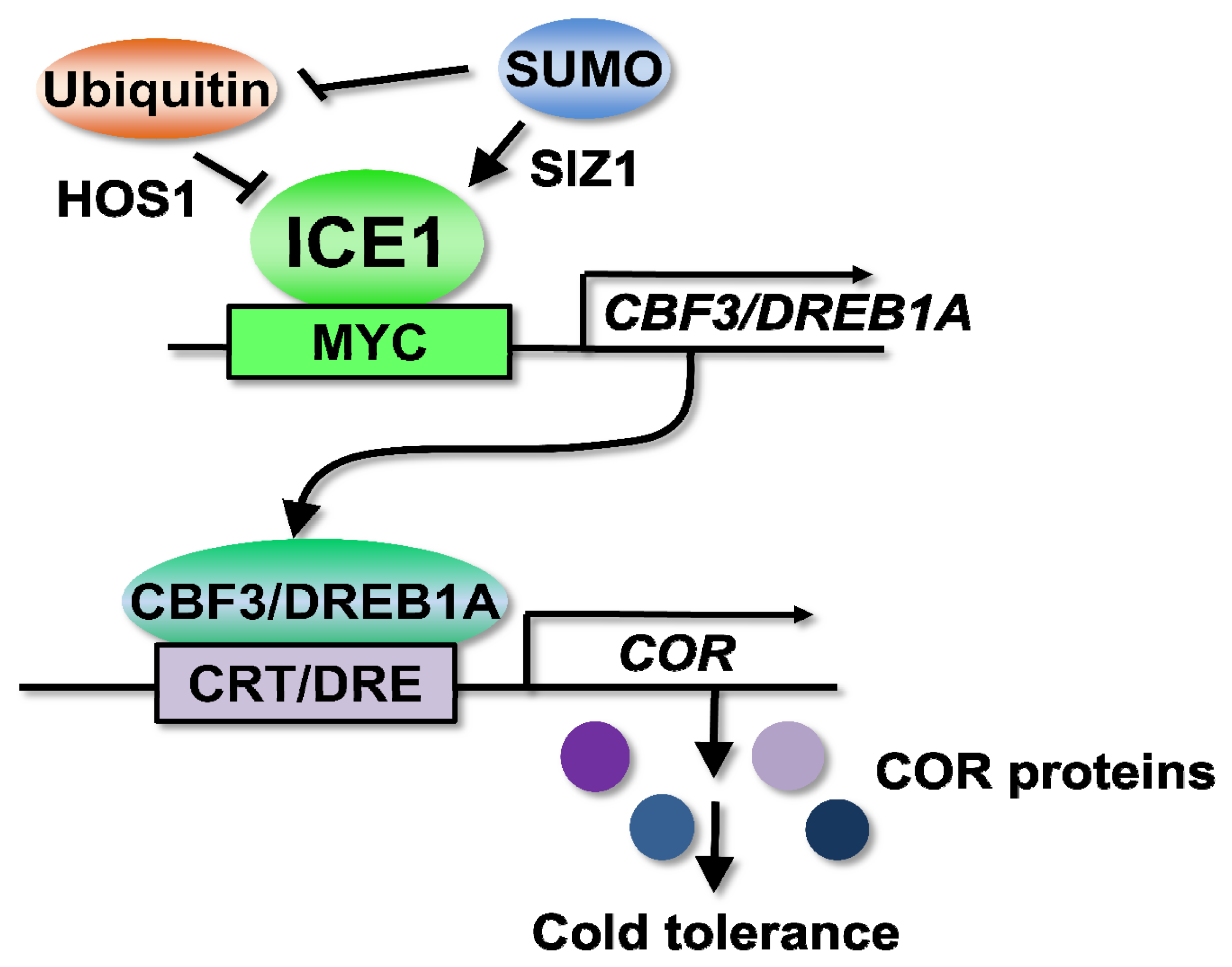{kind=link}
| Gene | Transgenic host | Source plant | Phenotype and effects | References |
|---|---|---|---|---|
| AtICE1 | Arabidopsis thaliana | Arabidopsis thaliana | Freezing tolerance; activation of CBF3/DREB1A | [21] |
| AtICE2 | Arabidopsis thaliana | Arabidopsis thaliana | Freezing tolerance; activation of CBF1/DREB1B | [23] |
| AtICE1 | Cucumis sativus | Arabidopsis thaliana | Chilling tolerance; dwarf | [32] |
| SlICE1 | Solanum lycopersicum | Solanum lycopersicum | Chilling tolerance; accumulation of antioxidants | [27,28] |
| TaICE141, TaICE187 | Arabidopsis thaliana | Triticum aestivum | Freezing tolerance | [29] |
| AtCBF1, AtCBF2, AtCBF3 | Arabidopsis thaliana | Arabidopsis thaliana | Freezing, salt and drought tolerance; constitutive expression of COR | [20,33,34] |
| OsDREB1A, OsDREB1B, OsDREB1C | Oryza sativa | Oryza sativa | Chilling, salt and drought tolerance; dwarf | [35] |
| HvCBF4 | Oryza sativa | Hordeum vulgare | Chilling, drought and salt tolerance | [36] |
| TaDREB2, TaDREB3 | Triticum aestivum | Triticum aestivum | Freezing and drought tolerance; dwarf | [37] |
| AtCBF1, AtCBF2, AtCBF3 | Brassica napus | Arabidopsis thaliana | Freezing tolerance; constitutive expression of COR | [38] |
| AtCBF1 | Fragaria ananassa | Arabidopsis thaliana | Freezing tolerance | [39] |
| AtCBF3 | Solanum tuberosum | Arabidopsis thaliana | Freezing tolerance | [40] |
| AtCBF1 | Populus tremula x alba | Arabidopsis thaliana | Freezing tolerance | [41] |
| AtCBF3 | Triticum aestivum | Arabidopsis thaliana | Freezing tolerance | [42] |
| AtCBF3 | Nicotiana tabacum | Arabidopsis thaliana | Freezing tolerance | [43] |
| SlCBF1 | Arabidopsis thaliana | Solanum lycopersicum | Freezing tolerance | [44] |
| OsDREB1A | Arabidopsis thaliana | Oryza sativa | Freezing, drought and salt tolerance | [45] |
| ZmDREB1A | Arabidopsis thaliana | Zea mays | Freezing and drought tolerance; dwarf | [46] |
| VrCBF1, VrCBF4 | Arabidopsis thaliana | Vitis riparia | Freezing and drought tolerance; dwarf | [47] |
| HvCBF3 | Arabidopsis thaliana | Hordeum vulgare | Freezing tolerance | [48] |
| LpCBF3 | Arabidopsis thaliana | Lolium perenne | Freezing tolerance; dwarf | [49,50] |
| SlCBF1 | Arabidopsis thaliana | Solanum lycopersicum | Chilling and oxidative tolerance | [51] |
| MbDREB1 | Arabidopsis thaliana | Malus baccata | Chilling, drought and salt tolerance | [52] |
| GmDREB3 | Arabidopsis thaliana | Glycine max | Freezing, drought and salt tolerance | [53] |
| BpCBF1 | Arabidopsis thaliana | Betula pendula | Freezing tolerance; dwarf | [54] |
| OsDREB1B | Nicotiana plumbaginifolia | Oryza sativa | Freezing, oxidative and drought tolerance; disease resistance | [55] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Miura, K.; Furumoto, T. Cold Signaling and Cold Response in Plants. Int. J. Mol. Sci. 2013, 14, 5312-5337. https://doi.org/10.3390/ijms14035312
AMA Style
Miura K, Furumoto T. Cold Signaling and Cold Response in Plants. International Journal of Molecular Sciences. 2013; 14(3):5312-5337. https://doi.org/10.3390/ijms14035312
Chicago/Turabian StyleMiura, Kenji, and Tsuyoshi Furumoto. 2013. "Cold Signaling and Cold Response in Plants" International Journal of Molecular Sciences 14, no. 3: 5312-5337. https://doi.org/10.3390/ijms14035312